


Abstract
The purpose of the present study was to evaluate the effect of 1,7-phenanthroline (PH), which has been proposed to be a selective phase II enzyme inducer, on the gene expression of xenobiotic transporters, as well as hepatic and renal drug-metabolizing enzymes. After oral administration of PH for 3 days to male Sprague-Dawley rats, mRNA levels in liver (75 and 150 mg/kg doses) and kidney (75 mg/kg dose only) were determined using real-time quantitative polymerase chain reaction. At 150 mg/kg/day, PH treatment resulted in significant increases in hepatic mRNA levels of Mrp3 (36-fold), UGT1A6 (20-fold), UGT2B1 (4-fold), and quinone reductase (QR, 5-fold), compared with the vehicle-treated group. Similar increases in Mrp3 (99-fold), UGT1A6 (17-fold), UGT2B1 (3-fold), and QR (11-fold) mRNA levels were observed in the liver after PH treatment of rats at 75 mg/kg/day. In contrast, the expression levels of CYP2C11 and Oatp2 were decreased by ∼80 and 50%, respectively. In addition, PH (75 mg/kg/day) elicited statistically significant changes in renal gene expression of CYP3A1, UGT1A6, QR, and Mrp3, but the magnitude of renal Mrp3 induction was less than 2-fold over control. Although PH is known to modulate hepatic glucuronidation in vivo, these data indicated that PH induced mRNA levels of the efflux transporter, Mrp3, which may also affect the disposition of xenobiotics.
Multidrug resistance proteins 1, 2, and 3 (Mrp1,1 2, and 3) have gained significance during the last few years because of their function as transporters of organic anions and conjugates, and their involvement in hepatic detoxification and tissue-specific distribution of drugs (Konig et al., 1999; Hirohashi et al., 1999; Borst et al., 2000). Notably, induction of hepatic rat Mrp3 and another transporter, Oatp2, by drugs like phenobarbital was implicated in the altered disposition of acetaminophen and the enhanced uptake of digoxin in rats (Rausch-Derra et al., 2001; Xiong et al., 2002a). Mrp3 induction has been shown also with oltipraz (Cherrington et al., 2002), a dithiolthione phase II enzyme inducer being investigated as a chemoprotectant against aflatoxin carcinogenicity and hepatotoxicity (Kwak et al., 2001a). Oltipraz and other selective phase II inducers, such as 1,7-phenanthroline (PH), offer a new mechanism of protection against carcinogenic and hepatotoxic compounds by inducing the enzymes involved in their metabolism [glutathione S-transferase (GST) and UDP-glucuronosyltransferase (UGT)], without increasing cytochrome P450-mediated bioactivation (Franklin and Moody, 1992; Franklin et al., 1993; Buetler et al., 1995; Vargas et al., 1998; Dong et al., 1999; Lamb and Franklin, 2000). Additionally, selective phase II inducers can be used to modulate in vivo exposure to acyl glucuronides. In this regard, a significant increase in the glucuronidation of benoxaprofen, a nonsteroidal anti-inflammatory drug, was observed when rats were treated with PH at 75 mg/kg/day for 3 days, resulting in a 2-fold increase in the biliary excretion of the acyl glucuronide, and an 8-fold increase in the peak plasma concentration (Cmax) and area under the curve values of the acyl glucuronide (Dong et al., 1999).
In the present study, we report that PH significantly induced not only mRNA levels of hepatic phase II enzymes, but also mRNA levels of Mrp3, which is likely to contribute to the altered metabolism and disposition of xenobiotics and their metabolites in PH-treated rats.
Materials and Methods
Materials. PH was purchased from Sigma-Aldrich (St. Louis, MO). The purity, verified by high-pressure liquid chromatography, was 99.5%. Solvents used for analysis were of analytical or high-pressure liquid chromatography grade (Fisher Scientific, Pittsburgh, PA).
In Vivo Animal Studies. All studies were reviewed and approved by the Merck Research Laboratories Institutional Animal Care and Use Committee. Male Sprague-Dawley rats were obtained from Charles River Laboratories Inc. (Wilmington, MA). After an overnight fast, five male Sprague-Dawley rats were dosed orally with PH at 150 mg/kg once daily for 3 days, and four male Sprague-Dawley rats were dosed with vehicle (0.1 M citric acid in 0.5% methylcellulose) in the same manner. Livers were removed from treated and vehicle control rats at 24 h after the last dose and stored at -70°C for quantitation of mRNA. In a separate experiment, three rats were dosed with PH at 75 mg/kg/day for 3 days, and an additional three rats were dosed with vehicle in the same manner. At 24 h after the last dose, the livers and kidneys were removed and stored at -70°C for mRNA quantification.
Development of Specific Primers and Probes for Quantitative Real-Time Polymerase Chain Reaction. Coding sequences for the genes listed in Table 1 were accessed from GenBank. Specific target regions within the coding sequences were determined through nucleotide sequence alignment comparisons of targets within multiple member gene families (e.g., Oatp2 with Oatps, Mrp2 with Mrps, CYP2C11 with CYP2C12, etc.). Primers and probes were designed to the selected target using Applied Biosystems Inc. (Foster City, CA) Primer Express software (v.2.0). All primers and probes were submitted to the National Center for Biotechnological Information for nucleotide comparison using the basic logarithmic alignment search tool (BLASTn) search for short, nearly exact sequences to ensure specificity. Primers and probes were synthesized by QIAGEN Operon (Alameda, CA), where primers were 5′- and 3′-labeled with the 6-carboxyfluorescein and 6-carboxytetram-ethylrhodamine reporter dyes, respectively. The rodent glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primer/probe set was purchased from Applied Biosystems Inc. and used per manufacturer's instructions. Each RNA sample was reverse-transcribed before analysis of different gene expression by PCR. Real-time quantitative PCR was performed using an ABI PRISM 7700 Sequence Detector instrument and Sequence Detector v.1.7 software (PerkinElmer Instruments, Skelton, CT).
Rat primer-probe sets and gene abbreviations
mRNA Isolation and Quantitative Real-Time Polymerase Chain Reaction. Total RNA from rat tissues was isolated using the SV Total RNA Isolation System (Promega, Madison, WI) according to the manufacturer's instructions. Samples were quantitated by spectrophotometry and diluted to a concentration of 15 ng/μl. Aliquots (500 ng) of RNA were analyzed by agarose/formaldehyde gel electrophoresis to check RNA integrity. Samples were then assayed in triplicate 25-μl reactions using 25 ng of RNA per reaction. Gene-specific primers were used at 7.5 pmol per reaction, and the gene-specific probes were used at 5 pmol per reaction. GAPDH was used to normalize gene expression in all samples since it is a highly expressed gene in rat liver and did not change in response to PH treatment [cycle threshold (Ct), control, 21.8 ± 0.2; PH, 22 ± 0.3]. Fold induction values were calculated by subtracting the mean difference of gene and GAPDH Ct number for each treatment group from the mean difference of gene and GAPDH Ct number for the vehicle group and raising this difference to the power of 2.
Statistical analyses were performed using a two-tailed Student's t test at α = 0.01 level of significance.
Results and Discussion
As expected from the literature (Vargas et al., 1998; Lamb and Franklin, 2000), significant increases in the amount of mRNA for UGT1A6 (20-fold) and, to a lesser extent, UGT2B1 (4-fold) and QR (5-fold) were detected in rat liver in response to PH treatment of 150 mg/kg/day (Fig. 1). The mRNA levels of UGT1A1 were not affected substantially (<2-fold) by this treatment. Similar increases in UGT1A6 (17-fold), UGT2B1 (3-fold), GST (6-fold), and QR (11-fold) mRNA were observed in liver after PH treatment of rats at 75 mg/kg/day for 3 days (Table 2). These results were similar to those reported by Vargas et al. (1998). The observed lack of dose proportionality in the increase of the mRNA of these enzymes between 75 and 150 mg/kg of PH needs further investigation to establish the time-dependence of the induction.
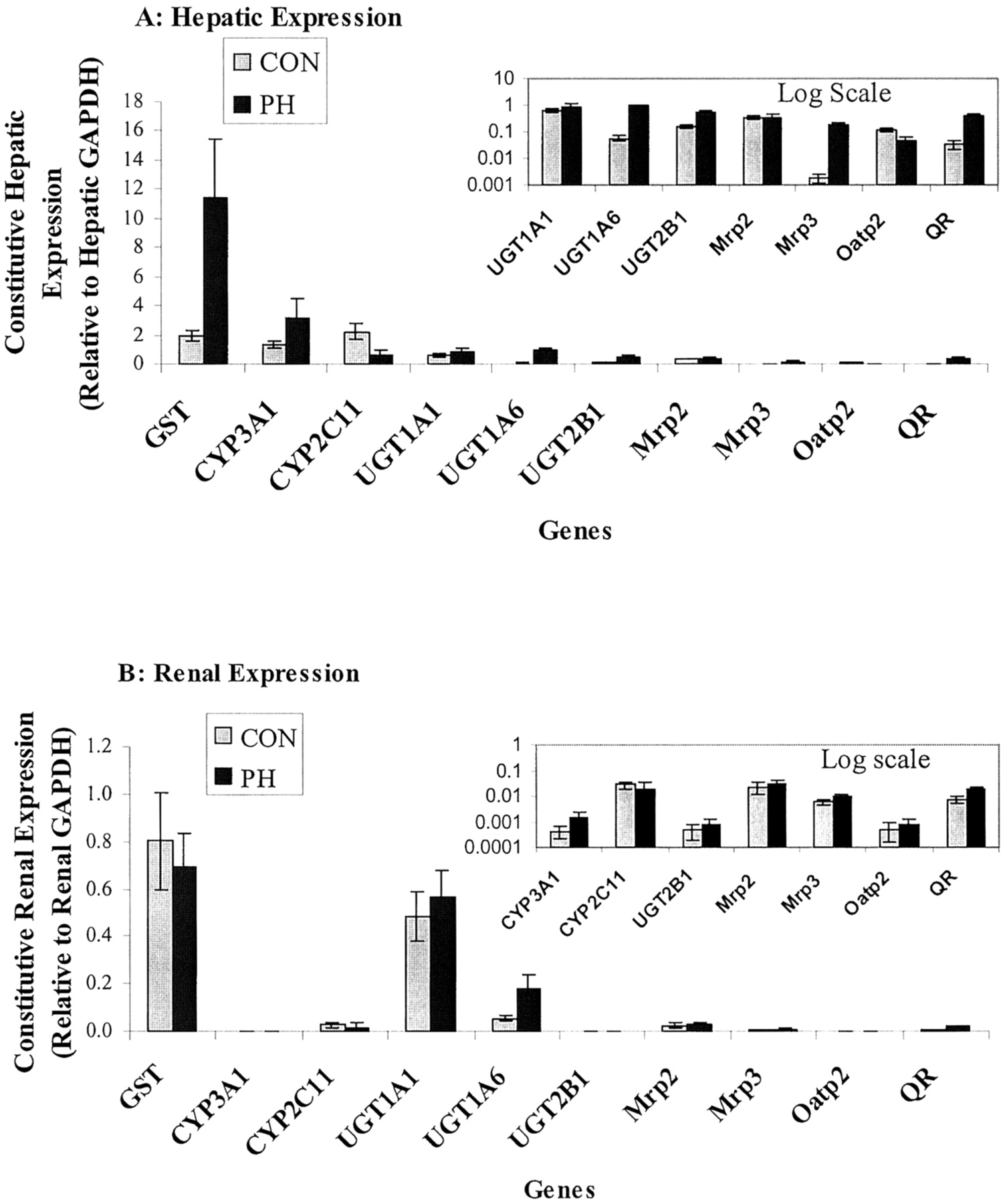
Comparison of constitutive (gray) and 1,7-phenanthroline-inducible (75 mg/kg/day for 3 days; black) expression of hepatic (A) and renal (B) drug-metabolizing enzymes and xenobiotic transporters.
Three animals were in each group; three determinations were performed for each animal. Some of the gene expression levels in log scale are shown in the box on the right in the figure.
Effects of 1,7-phenanthroline (75 mg/kg/day and 150 mg/kg/day, 3 days) on hepatic and kidney gene expression in male Sprague-Dawley rats
Data represent mean ± S.D.; n = 5 for the 150 mg/kg treatment group and n = 3 for the 75 mg/kg group. Three determinations were performed for each animal. Statistically significant differences between treated and control groups were evaluated by the Student's t test.
The transcription factor Nrf2 has been shown to be important for the induction of phase II enzymes. Induction of QR and UGT1A6 by oltipraz was observed in the wild-type, but not nrf2-deficient mice, which suggested that Nrf2 plays a major role in the regulation of these genes (Kwak et al., 2001b). Given that PH causes induction of a battery of genes similar to that of oltipraz, further studies are needed to elucidate the role of Nrf2, if any, in the inductive effects of PH.
PH caused a 3- to 4-fold induction of CYP3A1 gene expression in both liver and kidney; however, this is considered minor in comparison to the induction of CYP3A1 gene expression by pregnane X receptor agonists (∼30-fold) (Hartley and Klaassen, 2000) and may not be associated with increased protein levels (Dong et al., 1999). Significant decreases in the mRNA levels of CYP2C11 (17% of control) and Oatp2 (50% of control) were observed by PH treatment at 150 mg/kg/day. Down-regulation of CYP2C11 and Oatp2 by some aryl hydrocarbon receptor agonists, such as 3-methylcholanthrene, has been reported previously (Lee and Riddick, 2000; Rausch-Derra et al., 2001; Guo et al., 2002). However, PH does not induce CYP1A2 activity (Dong et al., 1999), suggesting that aryl hydrocarbon receptor may not be involved in the PH down-regulation of CYP2C11 and Oatp2.
In addition to the increases in QR and UGT mRNA levels, which are consistent with the previous report (Vargas et al., 1998), we also demonstrated that PH markedly increased hepatic Mrp3 mRNA levels (35-fold at 150 mg/kg and 99-fold at 75 mg/kg), but Mrp2 levels remained unaffected. Mrp3 is located on the basolateral membrane of polarized cells, and it plays a role in the hepatic elimination by transporting organic anions from liver to blood, which could lead to increased levels of xenobiotics and their metabolites in plasma and possibly an increase in urinary excretion (Konig et al., 1999; Kool et al., 1999). In normal rat liver, constitutive expression of Mrp3 mRNA is very low; however, the inducible nature of Mrp3 is well documented (Ogawa et al., 2000; Cherrington et al., 2002). Mrp3 has been shown to be induced by activators of the constitutive androstane receptor and an antioxidant/electrophile responsive element (Cherrington et al., 2002). However, a recent report by Xiong et al. (2002b) suggests that Mrp3 regulation occurs independent of constitutive androstane receptor.
In contrast to the robust increase in gene expression levels for UGT1A6 and Mrp3 in liver of PH-treated rats, there were much smaller changes in these genes in kidney by PH treatment at 75 mg/kg for 3 days (Table 2). The lack of evidence for a robust renal induction of xenobiotic transporters in rats treated with known hepatic enzyme inducers has been reported previously (Brady et al., 2002; Cherrington et al., 2002). In addition, previous studies have shown minimal inductive effects on drug-metabolizing genes by PH in the small intestine (Vargas et al., 1998).
Thus, PH is a pleiotropic inducer of genes responsible for drug metabolism and transport. These results indicate that in vivo data from PH-induced rats should be interpreted with caution, since up-regulation of Mrp3 gene expression may result in increased efflux of glucuronides and other xenobiotics from liver into the plasma compartment (Gregus et al., 1990; Xiong et al., 2002a). Further studies are needed to investigate the mechanism by which PH induces Mrp3 and phase II enzymes, and to determine whether these changes in mRNA levels will correlate with protein levels and/or activity.
Footnotes
-
↵ 1 Abbreviations used are: Mrp, multidrug resistance protein; Oatp, organic anion transporting polypeptide; PH, 1,7-phenanthroline; GST, glutathione S-transferase; UGT, UDP-glucuronosyltransferase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Ct, cycle threshold; QR, quinone reductase; Nrf2, nuclear factor-erythroid 2-related factor.
- Received December 10, 2002.
- Accepted March 12, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}