


Abstract
Posaconazole (Noxafil, SCH 56592), an orally available broad-spectrum triazole antifungal, is currently in phase III clinical studies for treating serious opportunistic fungal infections. The major in vitro metabolite of posaconazole formed by human liver microsomes supplemented with uridine 5′-diphosphate-glucuronic acid was a glucuronide of posaconazole (m/z877). Screening of 10 cDNA-expressed recombinant human UDP-glucuronosyltransferase (UGT) enzymes showed that only UGT1A4 exhibited catalytic activity with respect to the formation of the glucuronide of posaconazole. The formation of glucuronide by human liver microsomes and UGT1A4 was inhibited by bilirubin, a known inhibitor of UGT1A4. There was a high correlation (r =0.90) between the rate of formation of glucuronide, determined in 10 human liver microsomal samples, and trifluoperazine glucuronidation catalyzed by UGT1A4. These results confirmed that the formation of major posaconazole-glucuronide produced from human liver microsomes was mediated via UGT1A4.
UGTs1 are a superfamily of membrane-bound enzymes located in the endoplasmic reticulum and catalyze the conjugation of the sugar acid moiety of UDP glucuronic acid with many endogenous substrates such as steroid hormones, bilirubin, fat-soluble vitamins, and biogenic amines. UGTs convert lipophilic molecules into more polar and hydrophilic glucuronides facilitating their elimination via bile, feces, and urine. Although liver appears to be the major organ involved in glucuronidation, some UGT isoforms exist at high levels in the kidney and intestine, suggesting that extrahepatic glucuronidation can be significant (Tukey and Strassburg, 2000).
Posaconazole is a triazole derivative that is being developed as an antifungal agent (Barchiesi et al., 2001; Pfaller et al., 2001; Gonzalez et al., 2002). As with all azole antifungal agents, posaconazole works principally by inhibition of cytochrome P450 14α-demethylase (P45014DM). This enzyme is in the sterol biosynthesis pathway that leads from lanosterol to ergosterol. Compared with itraconazole, posaconazole is a significantly more potent inhibitor of sterol C14 demethylation, particularly in Aspergillus. Preliminary data showed that posaconazole and several glucuronide conjugates of posaconazole were present in human plasma and urine (P. Krieter, B. Flannery, T. Musick, M. Gohdes, M. Martinho, and R. Courtney, manuscript submitted for publication). The objective of this study was to identify the predominant in vitro UGT-conjugation pathway(s) for posaconazole.
Materials and Methods
Chemicals. UDP-glucuronic acid, Glucurase, magnesium chloride, Trizma base, saccharolactone (d-saccharic acid 1,4-lactone), alamethicin, trifluoperazine, bilirubin, and 4-trifluoromethyl-7-hydroxycoumarin glucuronide (4-trifluoromethylumbelliferyl glucuronide) were purchased from Sigma-Aldrich (St. Louis, MO). 7-Hydroxy-4-trifluoromethylcoumarin (7-HFC) was obtained from BD Gentest (Woburn, MA). Acetonitrile (HPLC grade) was obtained from Burdick & Jackson (Muskegon, MI). Dimethyl sulfoxide and methanol were obtained from Fisher Scientific (Pittsburgh, PA). Unlabeled and radiolabeled posaconazole (14C-SCH 56592, specific activity 80.3 μCi/mg, radiochemical purity 98.6%; Fig. 1) were prepared at Schering-Plough Research Institute (Kenilworth, NJ). Pooled human liver microsomes (n = 10) were purchased from XenoTech, LLC (Kansas City, KS). UGT Supersomes (human UGT enzymes developed using baculovirus-insect cell-expression system) were purchased from BD Gentest and Baculosomes (UGT1A7 and UGT1A10) were obtained from PanVera (Madison, WI).
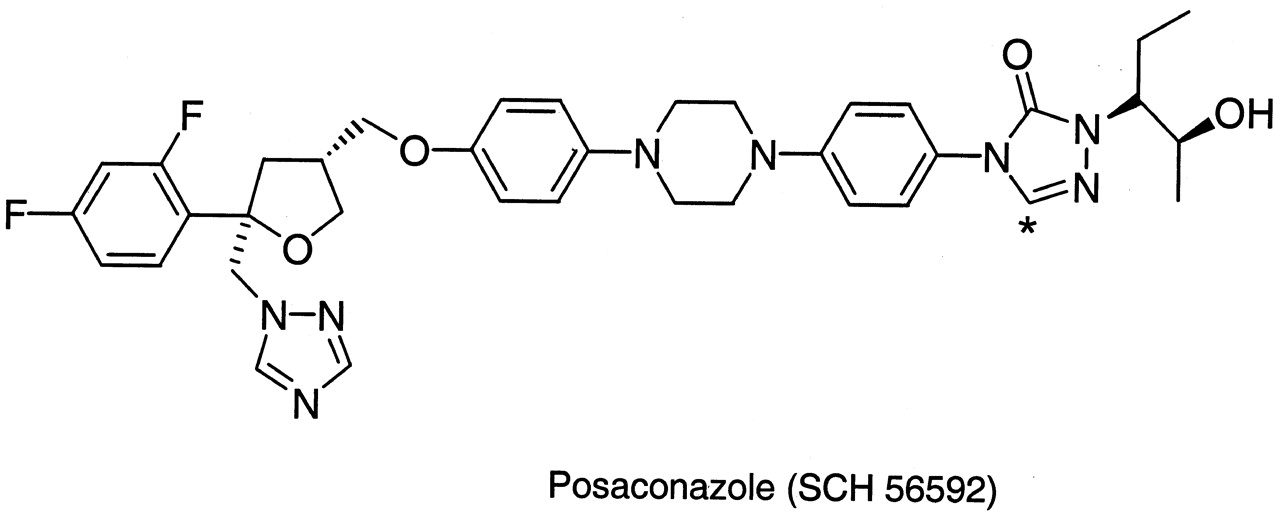
Chemical structure of posaconazole (SCH 56592).
* denotes site of 14C label
Analysis of Posaconazole and Glucuronide(s). The analysis was performed on a Waters Alliance HPLC system (Alliance model 2690; Waters Corporation, Milford, MA), equipped with a model 996 Photodiode Array Detector (Waters Corporation), a model 500TR Radioactivity Detector (PerkinElmer Life and Analytical Sciences, Boston, MA), and a 5-μm Zorbax RX-C8 250 × 4.6 mm column (Mac-Mod Analytical Inc., Chadds Ford, PA). The analytical column was maintained at room temperature for all HPLC experiments. The mobile phase, consisting of water (A) and methanol (B), was operated at a constant flow rate (1 ml/min), and the metabolite was detected at 260 nm. The gradient was 10% B for 0.1 min, followed by a rapid increase to 100% B in 46 min, 5 min at 100%, and a re-equilibrium at 10% B. After LC analysis, the concentrations of glucuronides were calculated based on radiometric detection and a standard curve (linear) of five points. All LC/MS and LC/tandem MS experiments were performed by using a TSQ 7000 mass spectrometer (Thermo Finnigan, San Jose, CA).
Enzyme Assays.Incubation of posaconazole with pooled human livermicrosomes. Substrate concentrations of 0.5 to 50 μM, microsomal protein concentrations of 0.05 to 2 mg/ml, and incubation times of 15 to 240 min were used to optimize the condition of the assay. Instead of treating with detergent, the microsomes were treated with alamethicin (a 20-amino acid α-helical pore-forming peptide) at 50 μg of alamethicin/mg of microsomal protein on ice for 15 min to diminish the latency of UGT activity (Fisher et al., 2000). All microsomal incubations contained microsomes treated with alamethicin, 10 mM magnesium chloride, 5 mM saccharolactone (an inhibitor of β-glucuronidase), and 14C-posaconazole in 0.5 ml of 0.1 M Tris-HCl buffer, pH 7.4 (Pless et al., 1999). Before the addition of UDPGA, the incubation mixtures were prewarmed for 3 min at 37°C. The reactions were initiated by the addition of 2 mM UDPGA, allowed to proceed for 120 min at 37°C, and terminated with ice-cold methanol. The incubation mixtures were vortexed and centrifuged (10,010g) at 4°C for 10 min, and the supernatants were analyzed by HPLC. Incubations without UDPGA and boiled human liver microsomes served as a negative control. For LC/MS analyses, the reactions were either terminated as described above or terminated by cooling in ice-water followed by solid phase extraction.
Determination of enzyme activities of UGT Supersomes and microsomes. The activities of UGT Supersomes (UGT1A1, UGT1A3, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B7, and UGT2B15) and human liver microsomes were determined in assays using HPLC and 7-HFC as substrate according to the UGT Batch Data Sheet provided by BD Gentest. The assay was performed with 1 mg/ml UGT Supersomes or microsomes treated with alamethicin (50 μg/ml), 10 mM magnesium chloride, 5 mM saccharolactone, and 50 μM 7-HFC in 0.1 M Tris-HCl buffer, pH 7.4. Before the addition of UDPGA, the incubation mixture was prewarmed for 3 min at 37°C. The reactions were initiated by the addition of 2 mM UDPGA and incubated for 20 min at 37°C; the reactions were terminated with 250 μl of ice-cold methanol. The incubation mixture was vortexed and centrifuged (10,010g) at 4°C for 10 min, and the supernatants were analyzed by HPLC. The formation of glucuronide was quantitated by comparing to the absorbance of a standard curve for 4-trifluoromethyl-7-hydroxycoumarin glucuronide.
Determination of enzyme activity of UGT1A4 Supersomes using trifluoperazine. UGT1A4 Supersomes do not form glucuronides with 7-HFC as a substrate. Therefore, the activity of UGT1A4 Supersomes was determined in assays using HPLC and trifluoperazine substrate as described by Dehal et al. (2001) with modifications. The assay was performed with 0.5 mg/ml UGT1A4 and 200 μM trifluoperazine substrate in 0.1 M Tris-HCl buffer, pH 7.4. The reactions were initiated by the addition of 2 mM UDPGA and incubated for 20 min at 37°C; the reactions were terminated with 250 μl of ice-cold methanol. The incubation mixture was vortexed and centrifuged (10,010g) at 4°C for 10 min, and the supernatants were analyzed by HPLC (Dehal et al., 2001).
Screening of 10 human UGT Supersomes. In vitro screening of 10 human UGT Supersomes (UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B7, and UGT2B15) with posaconazole were performed using a constant amount of microsomal protein (1 mg/ml) and 5 and 30 μM SCH 56592. All incubations with Supersomes were carried out as described earlier. Insect cell microsomes without UGT cDNA served as the negative control. These samples were also analyzed by LC/MS.
Kinetic parameters (Km and Vmax) of posaconazole glucuronidation by UGT1A4 Supersomes was determined as described below. Substrate concentrations of 0.5 to 50 μM, UGT protein concentrations of 1 mg/ml, and incubation time of 60 min were used to incubate UGT1A4 to determine Km and Vmax. The incubation was performed as described above, and insect microsomes without UGT cDNA served as negative control.
Inhibition with chemical inhibitors of UGTs. Inhibition of posaconazole metabolism was evaluated using bilirubin, a known chemical inhibitor of UGT1A4. Human liver microsomes (1 mg/ml) or human UGT Supersomes (1 mg/ml) were preincubated with various concentrations of bilirubin in dimethyl sulfoxide for 15 min at room temperature followed by the addition of buffer, alamethicin, saccharolactone, cofactor, and substrate. The final concentration of the organic solvent in the incubation system was 1 to 3%, and all control incubations contained the same volume of appropriate vehicle. All organic solvents tested up to 3% (v/v) had no effect on the formation of glucuronides. All incubations were carried out as described before except that the concentration of 14C-posaconazole was 30 μM in the case of human liver microsomes and 20 μM in the case of UGT1A4. The reactions were initiated by the addition of 2 mM UDPGA, incubated for 120 min (human liver microsomes) or 60 min (UGT1A4) at 37°C, and terminated with cold methanol. The incubation mixture was vortexed and centrifuged (10,010g) at 4°C for 10 min; the supernatants were saved for further analyses by HPLC.
Correlation Study. Ten individual human liver microsomal preparations from 10 individual donors (includes data for UGT-specific enzyme activities) were obtained from BD Gentest. The ability of human liver microsomes from each donor to glucuronidate posaconazole was correlated with the UGT-specific enzyme activities for each sample. The assays were performed as described previously with 30 μM substrate and incubated for 120 min.
Analysis of Kinetic Data. Untransformed enzyme kinetic data were analyzed by a nonlinear regression data analysis program (GraFit 4.0; Erithacus Software Ltd., Horley, Surrey, UK), assuming Michaelis-Menten kinetics over the substrate range studied.
Results
Optimization and Incubation with Pooled Human Liver Microsomes. After incubation of 30 μM 14C-posaconazole (Fig. 1) with pooled human liver microsomes and UDPGA, one major glucuronide was observed by radiometric detection (Fig. 2). As expected, incubation without UDPGA and with boiled microsomes did not yield any glucuronide (results not shown). LC/MS analysis of the extract following incubation of 14C-posaconazole with pooled human liver microsomes and UDPGA confirmed the presence of glucuronide (m/z 877). A posaconazole-glucuronide at this retention time was observed also in an in vivo study in healthy male subjects following oral administration of posaconazole (P. Krieter, B. Flannery, T. Musick, M. Gohdes, M. Martinho, and R. Courtney, manuscript submitted for publication). Four separate posaconazole-glucuronides were detected in plasma and urine (P. Krieter, B. Flannery, T. Musick, M. Gohdes, M. Martinho, and R. Courtney, manuscript submitted for publication). Based on NMR spectra and electronic calculations, this glucuronide was identified as N-glucuronide (P. Krieter, B. Flannery, T. Musick, M. Gohdes, M. Martinho, and R. Courtney, manuscript submitted for publication). However, the chemical structure of the other three glucuronides has not been identified.

Radiometric profile of metabolites following 120-min incubation of 30 μM 14C-posaconazole (14C-SCH 56592) with active human liver microsomes (top panel) or recombinant UGT1A4 Supersomes (bottom panel) supplemented with UDPGA.
When pooled human liver microsomes were incubated using various protein concentrations (0.05-2 mg/ml) and for various time periods (15-240 min), 1 mg/ml protein and 120 min were found to be optimal based on the linearity and percentage of conversion to its glucuronide. The effect of various substrate concentrations (0.5-50 μM) on posaconazole-glucuronides demonstrated that the apparent Km and Vmax values for the glucuronide formation were 27.9 μM and 15.3 pmol/mg protein/min, respectively (Table 1; Fig. 3). The intrinsic clearance (Vmax/Km) value for the glucuronide was estimated to be 0.55. A substrate concentration of 30 μM was chosen for further experiments with human liver microsomes considering the linearity and sensitivity of detection. In the absence of the UDPGA or with boiled microsomes, no metabolite formation was observed.
Kinetic parameters for the formation of glucuronide from 14C-posaconazole with pooled human liver microsomes and human UGT1A4 Supersomes

Enzyme kinetics for the formation of glucuronide conjugate of posaconazole (SCH 56592) incubated with pooled human liver microsomes (top panel) or UGT1A4 Supersomes (bottom panel).
Activities of Human UGT Supersomes and Human Liver Microsomes(Positive Control). The results of the activity determination of 10 human UGT Supersomes (UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B7, and UGT2B15) and human liver microsomes are presented in Table 2. These results served as positive control for the UGT activities and demonstrated that the Supersomes and microsomes were active. The data from these positive control experiments demonstrate that 7-HFC is a substrate for all UGTs studied except UGT1A4, whereas trifluoperazine is the substrate for only UGT1A4 (Table 2).
Determination of activities of cDNA-expressed human UGT Supersomes and human liver microsomes using 7-HFC and trifluoperazine (positive control) Values represent means of duplicate determinations.
Screening with Human UGT Supersomes. Incubation of posaconazole (5 and 30 μM) with 10 human UGT Supersomes (UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B7, and UGT2B15) revealed that only UGT1A4 metabolized posaconazole to the major in vitro glucuronide (Fig. 4). No metabolism was observed with other UGTs and control insect microsomes. The metabolite profiles suggested that only UGT1A4 is primarily responsible for the in vitro glucuronidation of posaconazole (Fig. 2). The recovery of metabolites from the HPLC column was on average >96%.
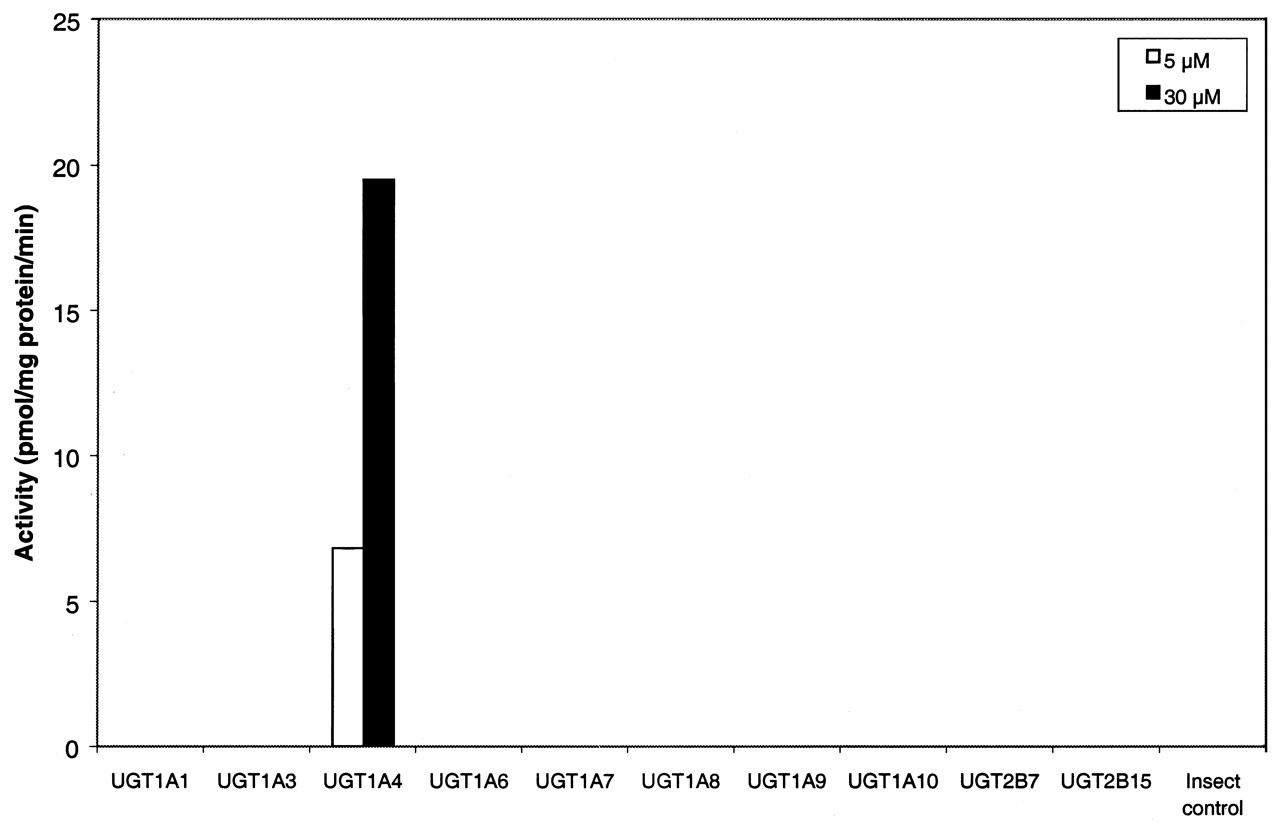
Screening of UGT Supersomes for the formation of glucuronide conjugate of posaconazole (SCH 56592) at 5 and 30 μM.
When UGT1A4 Supersomes were incubated using various protein concentrations (0.5-2 mg/ml) and for various time periods (15-120 min), 1 mg/ml protein and 60 min were found to be optimal based on the linearity and percentage of conversion to its glucuronide. Incubation of various concentrations of posaconazole (0.5-50 μM) with UGT1A4 Supersomes demonstrated that the apparent Km and Vmax values for posaconazole-glucuronide were 15.9 μM and 16.9 pmol/mg protein/min, respectively (Table 1; Fig. 3). Intrinsic clearance (Vmax/Km) for this glucuronide was estimated to be 1.06.
Inhibition Study. All inhibition studies were performed with pooled human liver microsomes at a drug concentration of 30 μM. UGT1A4 Supersomes were also incubated with inhibitors as positive control (20 μM posaconazole) to confirm the specificity of the inhibitors. The effect of incubations with UGT1A4-inhibitor and (bilirubin) is presented in Fig. 5 and Table 3. Bilirubin was shown to be a potent inhibitor of glucuronide formation by human liver microsomes (>79% inhibition). The IC50 value was 11.1 μM (Fig. 5). Bilirubin was also an inhibitor of UGT1A4 Supersomes (Table 3). These results suggested that UGT1A4 might be involved in the formation of glucuronide in human liver microsomes.

Inhibition curve of bilirubin (IC50) for the formation of glucuronide from posaconazole incubated with pooled human liver microsomes.
Inhibition of glucuronide formation following incubation of posaconazole with human liver microsomes and UGT1A4 in the presence of UGT-inhibitor bilirubin
Correlation Studies. The rate of formation of 14C-posaconazole-glucuronide was measured in each of the 10 human liver microsomal samples obtained from BD Gentest and correlated with the biochemical activities data provided by the vendor. Since the biochemical activity data were mediated by specific UGT enzymes, high correlation would suggest that similar enzymes were involved in the formation of metabolites from posaconazole.
The highest correlation between the BD Gentest assay data (n = 10) and the formation of glucuronide was noted for trifluoperazine glucuronidation (r = 0.90) catalyzed by UGT1A4 at 30 μM posaconazole (Fig. 6). Trifluoperazine was reported as a substrate of UGT1A4 in the literature by Ghosheh and Hawes (2002) and Linnet (2002). There was no significant correlation between estradiol 3-glucuronidation (catalyzed by UGT1A1, r = 0.35) or propofol glucuronidation (catalyzed by UGT1A9, r = 0.38) and posaconazole-glucuronide formation (results not shown). The results of correlation analysis between the enzyme activities and metabolite formation suggested that posaconazole is glucuronidated exclusively by UGT1A4 in human liver microsomes.

Correlation between formation of glucuronide from 30 μM posaconazole incubated separately with 10 individual human liver microsomes and trifluoperazine glucuronidation (UGT1A4 activity).
Discussion
Incubation of posaconazole with human liver microsomes supplemented with UDPGA resulted in the formation of one major metabolite (Fig. 2). This metabolite (m/z 877) was identified by LC/MS as the glucuronide of posaconazole (SCH 56592). Formation of this posaconazole-glucuronide was mediated by UGT1A4, based on the metabolite profiles following incubation of posaconazole with cell microsomes overexpressing UGT1A4, high correlation with trifluoperazine glucuronidation, and inhibition by bilirubin (Table 3; Figs. 4, 5, 6).
In vitro incubation with 10 different recombinant human UGT Supersomes showed that only UGT1A4 exhibited activity for the formation of posaconazole-glucuronide (Fig. 4). Bilirubin is a known specific inhibitor of UGT1A1 in the literature. However, bilirubin was also reported as a substrate and inhibitor of UGT1A4 by Hiller et al., 1999. Since the results of inhibition studies with recombinant UGT1A4 Supersomes showed that bilirubin was an inhibitor of posaconazole-glucuronide formation from UGT1A4 (Table 3), it was used as a UGT1A4-inhibitor in this study. At 100 μM concentration, bilirubin inhibited formation of the glucuronide from human liver microsomes and UGT1A4 Supersomes by 79.6% and 63.5%, respectively. In addition, there was highly significant correlation between the formation of posaconazole-glucuronide and trifluoperazine glucuronidation known to be mediated by UGT1A4 (Fig. 6). These results confirmed the involvement of UGT1A4 in the formation of posaconazole-glucuronide.
Footnotes
-
↵1 Abbreviations used are: UGT, UDP-glucuronosyltransferase; UDPGA, uridine 5′-diphosphate-glucuronic acid; 7-HFC, 7-hydroxy-4-trifluoromethylcoumarin; HPLC, high-performance liquid chromatography; SCH 56592, posaconazole; LC/MS, liquid chromatography/mass spectrometry; saccharolactone,d-saccharic acid 1,4-lactone.
- Received August 20, 2003.
- Accepted October 10, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}