


Abstract
The induction and inhibition of human cytochrome P450 (P450) enzymes are clinically responsible for drug interactions. Although the induction of P450s is investigated using human hepatocytes in the drug development process, there are some disadvantages, such as the decline of the enzyme activity during culture. In the present study, we examined the in vivo induction potency in chimeric mice with humanized liver, which was recently established in Japan to clarify whether this chimeric mouse model would be more suitable for human induction studies. Rifampicin and 3-methylcholanthrene (3-MC) were used in vivo as typical P450 inducers in the chimeric mice. The expression levels of human CYP3A4 mRNA and CYP3A4 protein and dexamethasone 6-hydroxylase activity, specific for human CYP3A4, were increased 8- to 22-, 3- to 10-, and 5- to 12-fold, respectively, by treatment with rifampicin. In addition, the expression levels of human CYP1A2 mRNA and CYP1A2 protein were also increased 2- to 9- and 5-fold, respectively, by treatment with 3-MC. Although other human P450s are expressed in the chimeric mice, there were few effects by the treatment of rifampicin and 3-MC on the mRNA, protein, and enzyme activity of those P450s. It was demonstrated that human P450s expressed in the chimeric mice with humanized liver were induced by rifampicin and 3-MC. This chimeric mouse model may be a useful animal model to estimate and predict the in vivo induction of P450s in humans.
Studies of drug metabolism are important for the determination of pharmacokinetic behavior and interindividual variability. Cytochrome P450 (P450) enzymes play a central role in the oxidative, peroxidative, and reductive metabolism of numerous endogenous compounds as well as drugs, environmental chemicals, and pollutants (Li et al., 1995). CYP3A4 is the predominant isoform in human liver and small intestine (Shimada et al., 1994; Ding and Kaminsky, 2003) and is responsible for the metabolism of many clinical drugs (Li et al., 1995). In clinical practice, serious drug interactions are frequently caused by the induction and inhibition of P450s (Dresser et al., 2000; Niemi et al., 2003). Induction is a long-term consequence of chemical exposure, whereas inhibition is an acute decrease of metabolism by another drug or a time-dependent decrease in the amount of an enzyme by several factors (Pelkonen et al., 1998). Therefore, the prediction of drug interactions involving P450s is essential during drug development.
Recently, human hepatocytes, human liver microsomes, and recombinant human P450 microsomes have become available as enzyme sources for in vitro experimental systems. The major limitation of microsomes is that they cannot be used for induction studies, whereas the limitation of in vivo studies using experimental animals is the existence of species differences. Human hepatocytes in primary culture are considered to be the most suitable model for induction studies but also have some problems, such as the inability to proliferate, the quick degradation of P450 activities during culture, and the requirement for specific culturing or technical conditions (Li et al., 1997). On the other hand, a transgenic mouse model containing human P450 was generated (Corchero et al., 2001; Robertson et al., 2003; Zhang et al., 2003). These transgenic mice were mainly used for studying transcriptional regulation. Thus, the development of a better model of the human liver is needed.
Recently, the generation of chimeric mice with humanized liver by the transplantation of human hepatocytes has been attempted (Dandri et al., 2001; Mercer et al., 2001). Using the urokinase-type plasminogen activator (uPA)/SCID mice, Tateno et al. (2004) succeeded in establishing chimeric mice whose livers could be replaced by more than 80% with human hepatocytes. At present, there are no reports of chimeric mice with as high a percentage of human hepatocytes as that reported by Tateno et al. (2004).
In the livers of these chimeric mice, we investigated the expression of human P450s (Katoh et al., 2004). In the present study, we investigated the in vivo induction of human P450s by the treatment of these chimeric mice with some model P450 inducers [rifampicin and 3-methylcholanthrene (3-MC)]. When we discuss the induction of P450, it is controversial whether the mRNA, protein, or enzyme activity is optimal to evaluate the induction. Roymans et al. (2004) reported that the induction ratios of CYP3A4 mRNA and testosterone 6β-hydroxylase activity (TESOH) by rifampicin in human hepatocytes QKR were greater than those in human hepatocytes 130, but the induction ratios of CYP3A4 protein were almost the same. On the other hand, Nallani et al. (2004) reported that there was a significant correlation between CYP3A4 protein and TESOH activity as well as between CYP3A4 protein and its mRNA after paclitaxel treatment using human hepatocytes. In induction studies using human hepatocytes or animals, some reports showed only the expression levels of mRNA, but others showed the protein or the enzyme activity. In the present study, we measured the changes of mRNA, protein, and enzyme activity.
Primarily, the changes of hepatic P450s needed to be evaluated because the induction of human P450s in the livers of the chimeric mice led to changes in the pharmacokinetics of the drugs. Therefore, we measured the mRNA, protein, and enzyme activity in the liver of the chimeric mice after rifampicin and 3-MC treatment.
Materials and Methods
Materials. All primers shown in Table 1 were commercially synthesized at Hokkaido System Sciences (Sapporo, Japan). Polyclonal rabbit anti-human CYP1A2 antibody, polyclonal rabbit anti-human CYP2A6 antibody, and polyclonal rabbit anti-human CYP2C8 antibody were purchased from Nosan (Yokohama, Japan). Polyclonal rabbit anti-human CYP1A1 antibody was purchased from Chemicon International (Temecula, CA). Polyclonal rabbit anti-human CYP2C9 antibody, monoclonal anti-human CYP2D6 antibody, and polyclonal rabbit anti-human CYP3A4 antibody were from BD Gentest (Worburn, MA). Pooled human liver microsomes and recombinant human CYP1A1, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2D6, and CYP3A4 expressed in baculovirus-infected insect cells were also from BD Gentest. Diclofenac and 3-MC were purchased from Sigma-Aldrich (St. Louis, MO). Coumarin, 7-hydroxycoumarin, dexamethasone, and rifampicin were purchased from Wako Pure Chemical Industries (Osaka, Japan). S-Mephenytoin and (±)-4′-hydroxymephenytoin were obtained from Toronto Research Chemicals (Toronto, Canada) and Sigma-Aldrich, respectively. 4′-Hydroxydiclofenac was purchased from BD Gentest. Nicotinamide adenine dinucleotide phosphate (oxidized form, NADP+) and glucose-6-phosphate dehydrogenase were purchased from Oriental Yeast (Tokyo, Japan). Nafamostat mesilate was kindly provided by Torii Pharmaceutical (Tokyo, Japan). All other chemicals and solvents were of the highest grade commercially available.
Sequence of the primers used in the present study
Generation of the Chimeric Mice with Humanized Liver. The present study was approved by the Ethics Committees of Kanazawa University and the Hiroshima Prefectural Institute of Industrial Science and Technology Ethics Board. The cryopreserved human hepatocytes from donor A (9-month-old white male) were purchased from In Vitro Technologies (Catonsville, MD). The human liver sample from donor B (12-year-old Japanese male) was obtained at autopsy after receiving written informed assent. The chimeric mice with humanized liver were generated by the method described previously (Tateno et al., 2004). Briefly, uPA+/+/SCID mice at 20 to 30 days after birth were injected with human hepatocytes through a small left-flank incision into the inferior splenic pole. When necessary, the chimeric mice were treated intraperitoneally with nafamostat mesilate. The role of nafamostat mesilate was to prolong survival and promote steady gains in body weight within the 2 months after transplantation, because the chimeric mice die due to complement-induced disorders in organs other than the liver (Tateno et al., 2004). The concentration of human albumin (hAlb) in the blood of the chimeric mice and the replacement index (RI; the rate of the replacement from mice to humans) were measured using enzyme-linked immunosorbent assay and anti-human specific cytokeratin 8 and 18 antibody, respectively. There was a good correlation between the hAlb concentration and RI (Tateno et al., 2004). The male chimeric mice used in this study were 11 to 14 weeks old (Table 2). The uPA+/+/SCID mice, uPA+/-/SCID mice, and uPA-/-/SCID mice were obtained as previously reported (Tateno et al., 2004).
Chimeric mice used in the present study
Animal Treatments. The chimeric mice, uPA+/-/SCID mice, and uPA-/-/SCID mice were intraperitoneally treated daily for 4 days with rifampicin (50 mg/kg/day) or 3-MC (20 mg/kg/day). The mice used in the present study are listed in Tables 2 and 3.
Control mice used in the present study
Hepatic RNA Extraction and Real-Time Reverse Transcription-PCR. Human P450 mRNA was quantified by real-time reverse transcription-PCR. Total hepatic RNA was extracted using ISOGEN (Nippon Gene, Tokyo, Japan), and cDNAs were synthesized as described previously (Iwanari et al., 2002). The sequences of primers that were specific to each human P450 are shown in Table 1. PCR was performed using the Smart Cycler (Cepheid, Sunnyvale, CA) with Smart Cycler software (version 1.2b). The PCR conditions were as follows. After an initial denaturation at 95°C for 30 s, amplification was performed by denaturation at 94°C for 4 s, and annealing and extension were performed at 64°C for 20 s for 45 cycles. Amplified products were monitored directly by measuring the increase of the dye intensity of the SYBR Green I (Molecular Probes, Eugene, OR) that binds to double-strand DNA amplified by PCR. The copy number of mRNA in the cDNA samples was calculated using standard amplification curves. It was confirmed that the primer for human P450s used in this study did not cross-react with murine P450 mRNA.
Liver Microsomes. Liver microsomes from the chimeric or control mice were prepared as described previously (Yamazaki et al., 1999) and stored at -80°C until analysis. The protein concentration was determined using Bradford protein assay reagent (Bio-Rad, Hercules, CA) with bovine γ globulin as the standard.
Immunoblot Analysis of Human P450 Isoforms. SDS-polyacrylamide gel electrophoresis and immunoblot analysis of human CYP1A1, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2D6, and CYP3A4 were performed according to Laemmli (1970) with slight modifications. The liver microsomes were separated on 7.5% polyacrylamide gel and transferred electrophoretically to a polyvinylidene difluoride membrane. Recombinant human P450s were also applied as the standards. Biotinylated anti-rabbit or mouse IgG and a VECTASTAIN ABC kit (Vector Laboratories, Burlingame, CA) were used for diaminobenzidine staining.
Enzyme Assays. The typical incubation mixtures (total volume, 0.20 ml) consisted of microsomes in 100 mM potassium phosphate buffer (pH 7.4) containing an NADPH-generating system (0.5 mM NADP+, 5 mM glucose 6-phosphate, 5 mM MgCl2, and 1 U/ml glucose-6-phosphate dehydrogenase) and a substrate. Coumarin 7-hydroxylase activity (COH) catalyzed by CYP2A6 was measured as described previously (Katoh et al., 2004). In brief, the concentrations of microsomes and coumarin were 0.1 mg/ml and 1 μM, respectively. The reaction mixture was incubated for 3 min at 37°C. The product formation was determined using high-performance liquid chromatography (HPLC) with a C18 5-μm analytical column (4.6 × 150 mm). Diclofenac 4′-hydroxylase activity (DICOH) catalyzed by CYP2C9 was determined as described previously (Katoh et al., 2004). The concentrations of microsomes and diclofenac were 0.2 mg/ml and 30 μM, respectively. The reaction mixture was incubated for 30 min at 37°C. The mobile phase was 22% acetonitrile in 50 mM phosphate buffer (pH 7.4). The product formation was determined using HPLC with a C18 5-μm analytical column (4.6 × 150 mm). S-Mephenytoin 4′-hydroxylase activity (MPOH) catalyzed by CYP2C19 was measured as described previously (Katoh et al., 2004). The concentrations of microsomes and S-mephenytoin were 0.2 mg/ml and 200 μM, respectively. The reaction mixture was incubated for 30 min at 37°C. The product formation was determined using HPLC with a C18 5-μm analytical column (4.6 × 150 mm). The eluent was monitored at 204 nm with a noise-base clean Uni-3 (Union, Gunma, Japan). The mobile phase was 18% acetonitrile in 50 mM potassium dihydrogen phosphate. Dexamethasone 6-hydroxylase activity (DEXOH) catalyzed by CYP3A4 was examined according to the method described previously (Katoh et al., 2004). The concentrations of microsomes and dexamethasone were 0.2 mg/ml and 100 μM, respectively. The reaction mixture was incubated for 30 min at 37°C. The product formation was determined using HPLC with a C8 5-μm analytical column (4.6 × 150 mm). The mobile phase was acetonitrile/0.018% formic acid = 18:82 (v/v). The eluent was monitored at 243 nm with a Uni-3. DEXOH was quantified using a standard curve of dexamethasone because we could not obtain authentic 6-hydroxydexamethasone. The retention time of 6-hydroxydexamethasone was confirmed using the incubation product of recombinant CYP3A4 and dexamethasone. The final concentration of the solvent in the incubation mixture was <1%. Data were analyzed using the mean of duplicate determinations.
Effect of Nafamostat Mesilate of Human P450 on Induction by Rifampicin or 3-MC in Chimeric Mice. To investigate the effect of nafamostat mesilate of human P450 on the induction by rifampicin or 3-MC, the induction of human P450 was compared between chimeric mice treated with and without nafamostat mesilate. The chimeric mice without nafamostat mesilate were not treated from 3 days before the start of the inducer treatment to the last day of this study.

Selectivity of human P450 antibodies to human P450 on immunoblot analysis. Immunoblot analyses of microsomes from pooled human liver, an induced chimeric mouse, and an induced uPA+/-/SCID mouse were performed using human CYP3A4 antibodies (A) and human CYP1A2 antibodies (B). A, lanes of human liver microsomes (H) and chimeric mouse 3 were 10 μg; lanes of chimeric mouse 13, M2, and M5 were 2 μg. The recombinant human CYP3A4 from BD Gentest (0.2, 0.7, and 0.9 pmol) was used as a standard. B, lanes of H and chimeric mouse 3 were 10 μg; lanes of chimeric mouse 24, M3, and M6 were 1 μg. The recombinant human CYP1A2 from BD Gentest (25, 50, 100, and 200 fmol) was used as a standard. M2, rifampicin-treated uPA+/-/SCID mouse; M3, 3-MC-treated uPA+/-/SCID mouse; M4, rifampicin-treated uPA-/-/SCID mouse; M5, R3-MC-treated uPA -/-/SCID mouse.
Results
Chimeric Mice Used in the Present Study. Ten and four chimeric mice generated using hepatocytes from donor A and B were used in the present study (Table 2). The hAlb concentration, approximate RI, and inducer are shown in Table 2. The albumin concentration in the chimeric mice was measured 1 day before starting the inducer treatment. We confirmed that the expression of all hepatic mRNA and enzyme activities in uPA+/-/SCID mice and uPA-/-/SCID mice were not affected by the administration of nafamostat mesilate (data not shown). To investigate the induction of P450s, the expression levels of P450 mRNA and the protein content and enzyme activity in a chimeric mouse were compared to those in a chimeric mouse with a similar hAlb concentration or RI. The expression levels of human glyceraldehyde-3-phosphate dehydrogenase (hGAPDH) mRNA were increased in a hAlb concentration-dependent manner without the effect of inducers (data not shown).
Selectivity of Human P450 Antibodies to Human Microsomes in Immunoblot Analysis. The selectivities of human P450 antibodies, especially human CYP3A4 (Fig. 1A) and human CYP1A2 (Fig. 1B), were investigated. The amounts of microsomal protein used in those immunoblot analyses, which were determined not to cross-react with the autologous murine P450 proteins using induced murine liver microsomes, were as follows: CYP1A2, 10 μg (no treatment) and 1 μg (3-MC treatment); CYP2A6, 20 μg; CYP2C9, 5 μg; and CYP3A4, 10 μg (no treatment) and 2 μg (rifampicin treatment). Human CYP1A1 antibodies were raised against a C-terminal peptide of human CYP1A1 and did not cross-react with induced murine liver microsomes (data not shown).
Induction of Human CYP3A4 in Rifampicin-Treated Chimeric Mice. The effects of rifampicin treatment on CYP3A4 expression are shown in Fig. 2. The CYP3A4/hGAPDH mRNA ratios in rifampicin-treated chimeric mice with hepatocytes from donor A and B were 8.2- and 22.1-fold higher, respectively, than those in nontreated chimeric mice with similar concentrations of hAlb (Fig. 2A). The copy number of CYP3A4 mRNA was correlated with the hAlb concentration (Fig. 2B; none, r = 0.77; rifampicin, r = 0.97), and the slope of the fitted curve was increased in rifampicin-treated chimeric mice compared with nontreated chimeric mice. Treatment with rifampicin increased the levels of CYP3A4 protein by 9.8-fold in donor A chimeric mice and 3.0-fold in donor B chimeric mice (Fig. 2C). Rifampicin caused a significant enhancement of DEXOH in both donor A chimeric mice (5.1- to 5.3-fold; mean, 5.2) and donor B chimeric mice (10.0- to 14.6-fold; mean, 12.0) but no enhancement in uPA+/-/SCID and uPA-/-/SCID mice.

Induction of human CYP3A4 expression in rifampicin-treated chimeric mice. Hepatic CYP3A4 mRNAs in the chimeric mice were measured by real-time PCR (A and B). The CYP3A4 mRNA was expressed as the relative expression to hGAPDH (A, CYP3A4/hGAPDH) and copy number (B, arbitrary unit). The CYP3A4 protein content (C) and DEXOH catalyzed by CYP3A4 (D) were determined by Western blot analysis and HPLC, respectively. A, C, and D, open and closed columns are expressed values of the non- and rifampicin-treated chimeric mice, respectively. B, circles and triangles represent the chimeric mice with hepatocytes from donor A and B, respectively. The open and closed symbols represent non- and rifampicin-treated chimeric mice, respectively. D, columns of M1, M2, M4, and M5 represent the mean ± S.D. (n = 3). None, nontreated; Rif, rifampicin-treated; M1, nontreated uPA+/-/SCID mouse; M2, rifampicin-treated uPA+/-/SCID mouse; M4, nontreated uPA-/-/SCID mouse; M5, rifampicin-treated uPA-/-/SCID mouse.
Changes of Human CYP2A6 Expression in Rifampicin-Treated Chimeric Mice. The effects of rifampicin treatment on the CYP2A6 expression are shown in Fig. 3. The CYP2A6 mRNA, protein, and COH in the chimeric mice from donor B hepatocytes were not detected since these chimeric mice were genotyped as homozygous for the human CYP2A6*4 allele (Katoh et al., 2004), which deletes the whole human CYP2A6 gene. The CYP2A6/hGAPDH mRNA ratios in rifampicin-treated chimeric mice with hepatocytes from donor A were 1.7-fold higher than those in nontreated chimeric mice with similar concentrations of hAlb (Fig. 3A). Rifampicin treatment increased the CYP2A6 protein and COH by 3.5- and 2.4-fold, respectively (Fig. 2, B and C). There were no changes of COH caused by rifampicin treatment in either uPA+/-/SCID or uPA-/-/SCID mice.
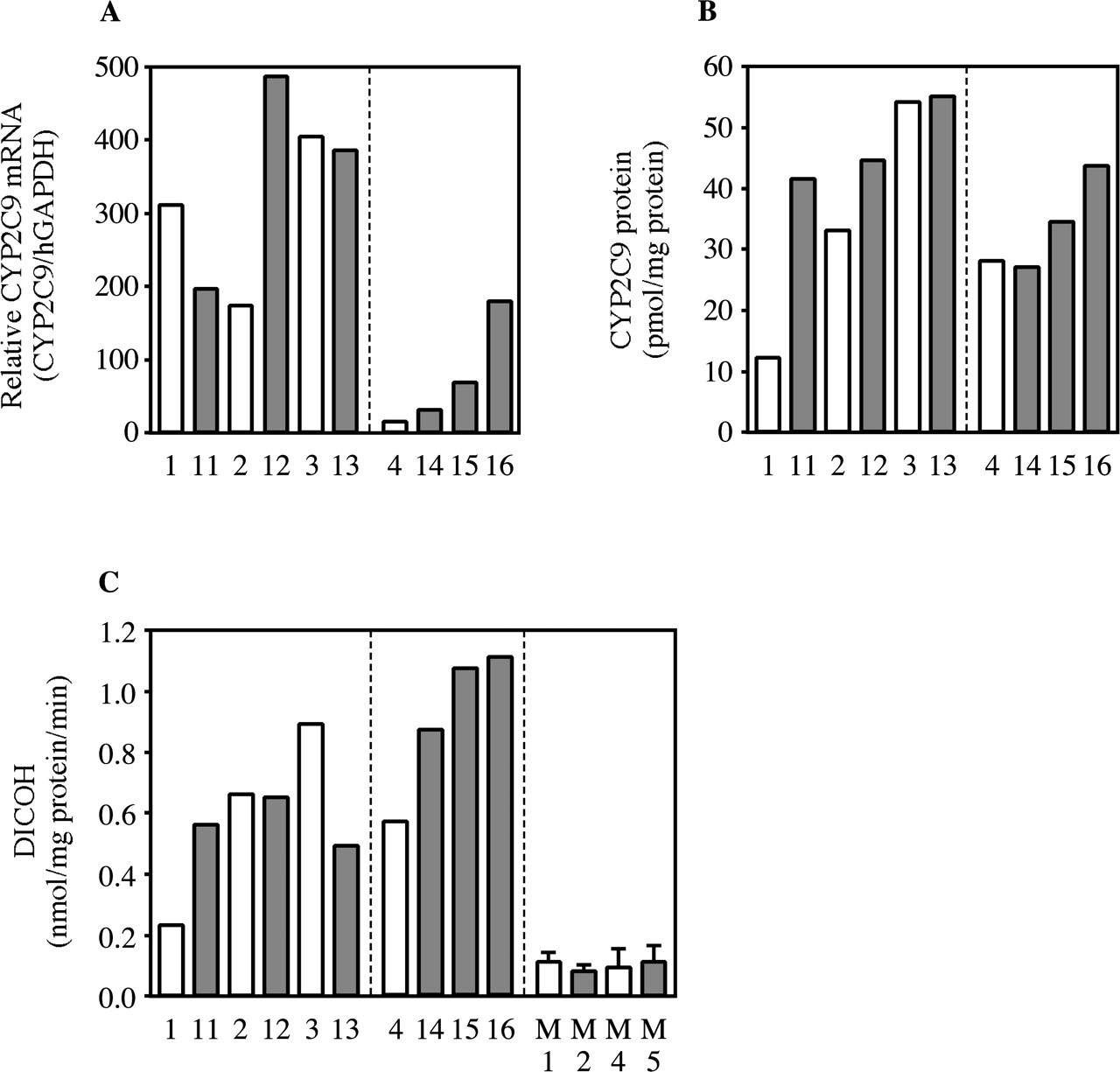
Changes of Human CYP2C9 Expression in Rifampicin-Treated Chimeric Mice. The effects of rifampicin treatment on CYP2C9 expression are shown in Fig. 4. The CYP2C9/hGAPDH mRNA ratio, protein content, and DICOH in rifampicin-treated chimeric mice with hepatocytes from donor B were 22.1-, 1.3-, and 1.8-fold higher, respectively, than those in nontreated chimeric mice with similar concentrations of hAlb (Fig. 4). On the other hand, the chimeric mice with hepatocytes from donor A did not exhibit evidence of the induction. No changes of DICOH were caused by rifampicin treatment in either uPA+/-/SCID or uPA-/-/SCID mice.
Changes of Human CYP2C19 Expression in Rifampicin-Treated Chimeric Mice. The effects of rifampicin treatment on CYP2C19 expression are shown in Fig. 5. The CYP2C19/hGAPDH mRNA ratios in rifampicin-treated chimeric mice with hepatocytes from donor A and B were higher than those in nontreated chimeric mice (Fig. 5A). The content of human CYP2C19 protein could not be quantified because no applicable primary antibodies are commercially available. There was not enough MPOH to calculate the induction ratio in the chimeric mice with donor B hepatocytes, but there was a 2.4-fold increase by rifampicin treatment in the chimeric mice with donor A hepatocytes (Fig. 5B). No changes in MPOH were caused by rifampicin treatment in either uPA+/-/SCID mice or uPA-/-/SCID mice.
Changes of Other Human P450s in Rifampicin-Treated Chimeric Mice. The mRNA and protein expressions of human CYP1A2 and CYP2D6 were not affected by treatment with rifampicin. The expression levels of human CYP2C8 mRNA in rifampicin-treated chimeric mice were 1.4- to 3.7-fold higher than those in nontreated chimeric mice. Similarly, the expression of human CYP2C8 protein was increased 1.6- to 2.5-fold by rifampicin (data not shown).
Induction of Human CYP1A2 Expression in 3-MC-Treated Chimeric Mice. The effects of 3-MC treatment on human CYP1A2 (hCYP1A2) expression are shown in Fig. 6. The hCYP1A2/hGAPDH mRNA ratios in 3-MC-treated chimeric mice were 2.1- to 8.6-fold higher than those in nontreated chimeric mice (Fig. 6A). The slope of the fitted curve of human CYP1A2 mRNA was increased in 3-MC-treated chimeric mice compared with nontreated chimeric mice (Fig. 6B; none, r = 0.86; 3-MC, r = 0.96). The 3-MC treatment increased the content of human CYP1A2 protein by 4.9-fold (3.6–6.4) in donor A chimeric mice (Fig. 6C). Because of the lack of a substrate with specificity to human CYP1A2 that does not cross-react with murine Cyp1a, the induction of human CYP1A2 activity in the chimeric mice could not be evaluated.
Changes of Human CYP1A1 Expression in 3-MC-Treated Chimeric Mice. The effects of 3-MC on human CYP1A1 (hCYP1A1) expression are shown in Fig. 7. The hCYP1A1/hGAPDH mRNA ratios were increased 18.7-fold (10.0–37.7) by 3-MC treatment (Fig. 7A). The slope of the fitted curve of human CYP1A1 mRNA was increased in 3-MC-treated chimeric mice compared with nontreated chimeric mice (Fig. 7B; none, r = 0.83; 3-MC, r = 0.82). Human CYP1A1 proteins in both non- and 3-MC-treated chimeric mice were below 0.6 fmol/mg protein.

Changes of human CYP2A6 expression in rifampicin-treated chimeric mice. Hepatic CYP2A6 mRNAs in the chimeric mice measured by real-time PCR were expressed as the relative expression to hGAPDH (A, CYP2A6/hGAPDH). The CYP2A6 protein content (B) and COH catalyzed by CYP2A6 (C) were determined by Western blot analysis and HPLC, respectively. Open and closed columns are the values of the non- and rifampicin-treated chimeric mice, respectively. C, columns of M1, M2, M4, and M5 represent the mean ± S.D. (n = 3). ND, not detected; M1, nontreated uPA+/-/SCID mouse; M2, rifampicin-treated uPA+/-/SCID mouse; M4, nontreated uPA-/-/SCID mouse; M5, rifampicin-treated uPA-/-/SCID mouse.
Changes of Other Human P450s in Chimeric Mice. The changes of CYP3A4 protein, CYP2A6 protein, CYP2C9 protein, DEXOH, COH, DICOH, and MPOH by 3-MC treatment are shown in Fig. 8. The treatment with 3-MC did not affect those enzyme activities. Similarly, in both uPA+/-/SCID and uPA-/-/SCID mice, there were no changes in DEXOH, COH, DICOH, and MPOH by 3-MC treatment. The expression levels of mRNA and protein in CYP2C8 and CYP2D6 exhibited no changes by 3-MC treatment (data not shown).
Effect of Nafamostat Mesilate of Human P450 on Induction by Rifampicin or 3-MC in Chimeric Mice. In the induction study of rifampicin, DEXOH was 38.7, 43.7, and 59.2 pmol/mg protein/min in chimeric mice 11, 12, and 13, respectively, which were treated with both rifampicin and nafamostat mesilate (Fig. 2D). In the chimeric mice treated with rifampicin but not with nafamostat mesilate, DEXOH was 34.1 (hAlb concentration = 5.0 mg/ml), 43.3 (hAlb concentration = 7.6 pmol/mg protein/min), and 45.8 (hAlb concentration = 9.6 mg/ml) pmol/mg protein/min. On the other hand, in the induction study of 3-MC, the expression levels of human CYP1A2 protein were 13.7, 17.9, and 17.8 pmol/mg protein in chimeric mice 22, 23, and 24, respectively (Fig. 6C). In the chimeric mice treated with 3-MC but not with nafamostat mesilate, the expression levels of human CYP1A2 protein were 10.6 (hAlb concentration = 5.1 mg/ml), 17.0 (hAlb concentration = 7.6 mg/ml), and 20.0 pmol/mg protein (hAlb concentration = 9.9 mg/ml). Nafamostat mesilate treatment caused no significant differences in the induction of DEXOH by rifampicin and CYP1A2 protein by 3-MC.
Discussion
The induction and inhibition of P450 enzymes are considered to cause many drug interactions. In the drug development process, it is extremely important to investigate whether a drug candidate will be an inducer or an inhibitor of P450s to predict potential drug interactions in humans (Lin and Lu, 2001). The induction of P450 is defined as an increase in P450 activity associated with an increase in the intracellular P450 concentration (Ronis and Ingelman-Sundberg, 1999). At present, the inhibition of P450s in humans can be predicted relatively easily from in vitro approaches using recombinant human P450s and/or human liver microsomes; however, the results of P450 induction studies obtained from experimental animals are difficult to extrapolate to humans due to species differences. Primary-cultured human hepatocytes are considered to be a more appropriate tool for the evaluation of the induction in humans; however, it has been reported that P450 mRNA in human hepatocytes declined rapidly after the isolation from liver (Gómez-Lechón et al., 2003). Moreover, the supply of human hepatocytes is sometimes limited.
Recently, chimeric mice with humanized liver were established by Tateno et al. (2004). The livers of the mice could be replaced by more than 80% with human hepatocytes. We clarified that major human P450s were expressed in such livers and that the enzyme activities were almost the same as those of the donor (Katoh et al., 2004). In this study, we investigated the induction potency of human P450s in chimeric mice with humanized liver to determine whether the chimeric mice can be a useful tool in studies of human P450 induction.

Changes of human CYP2C9 expression in rifampicin-treated chimeric mice. Hepatic CYP2C9 mRNAs in the chimeric mice measured by real-time PCR were expressed as the relative expression to hGAPDH (A, CYP2C9/hGAPDH). The CYP2C9 protein content (B) and DICOH catalyzed by CYP2C9 (C) were determined by Western blot analysis and HPLC, respectively. Open and closed columns are the values of the non- and rifampicin-treated chimeric mice, respectively. C, columns of M1, M2, M4, and M5 represent the mean ± S.D. (n = 3). M1, nontreated uPA+/-/SCID mouse; M2, rifampicin-treated uPA+/-/SCID mouse; M4, nontreated uPA-/-/SCID mouse; M5, rifampicin-treated uPA-/-/SCID mouse.

Changes of human CYP2C19 expression in rifampicin-treated chimeric mice. Hepatic CYP2C19 mRNAs in the chimeric mice measured by real-time PCR were expressed as the relative expression to hGAPDH (A, CYP2C9/hGAPDH). The MPOH catalyzed by CYP2C19 (B) was determined by HPLC. Open and closed columns are the values of the non- and rifampicin-treated chimeric mice, respectively. B, columns of M1, M2, M4, and M5 represent the mean ± S.D. (n = 3). ND, not detected; M1, nontreated uPA+/-/SCID mouse; M2, rifampicin-treated uPA+/-/SCID mouse; M4, nontreated uPA-/-/SCID mouse; M5, rifampicin-treated uPA-/-/SCID mouse.
There are many drug interactions with rifampicin caused by the induction of drug-metabolizing enzymes such as CYP3A4 (Niemi et al., 2003). It has been reported that the exposure of human hepatocytes to rifampicin caused an approximately 10- and 5-fold increase in the expression levels of CYP3A4 mRNA and the protein content, respectively (Drocourt et al., 2001; Desai et al., 2002; Raucy et al., 2002). The induction effects of rifampicin on TESOH catalyzed by CYP3A4 in human hepatocytes averaged 10-fold compared with the control, but large interindividual differences were reported (Desai et al., 2002; Madan et al., 2003). In the present study, rifampicin significantly induced CYP3A4 mRNA (8.2-fold in donor A chimeric mice and 22.1-fold in donor B chimeric mice) and increased the protein content (9.8- and 3.0-fold) and DEXOH (5.2- and 12.0-fold) in a manner consistent with previous reports. As we described, it is not always the case that the induction between the mRNA, protein, and enzyme activity can be correlated. This point concerning both constitutive and inducible expression is still controversial. This phenomenon is observed not only in chimeric mice, but also in human hepatocytes.

Changes of human CYP1A2 expression in 3-MC-treated chimeric mice. Hepatic human CYP1A2 mRNAs in the chimeric mice were measured by real-time PCR (A and B). The human CYP1A2 mRNA was expressed as the relative expression to human GAPDH (A, hCYP1A2/hGAPDH) and the copy number (B, arbitrary unit). The human CYP1A2 protein content (C) was determined by Western blot analysis. A and C, open and closed columns are values of the non- and 3-MC-treated chimeric mice, respectively. None, nontreated; Rif, rifampicin-treated; ND, not detected.
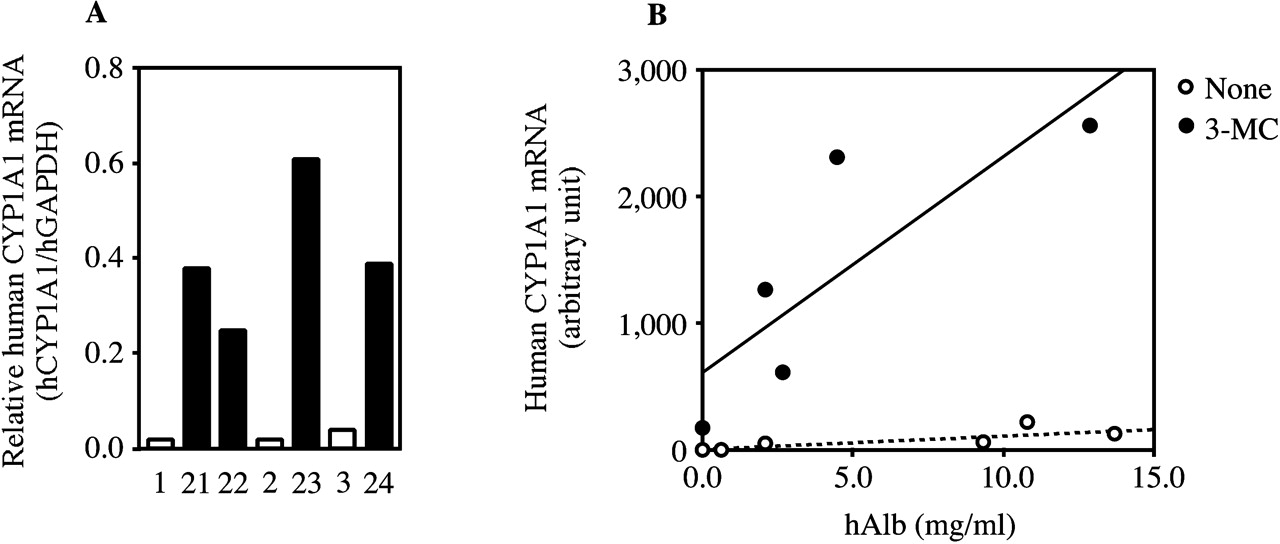
Changes of human CYP1A1 expression in 3-MC-treated chimeric mice. Hepatic human CYP1A1 mRNAs in the chimeric mice were measured by real-time PCR (A and B). The human CYP1A1 mRNA was expressed as the relative expression to human GAPDH (A, hCYP1A1/hGAPDH) and the copy number (B, arbitrary unit). A, open and closed columns are the expressed values of the non- and 3-MC-treated chimeric mice, respectively. None, nontreated; Rif, rifampicin-treated.
Rifampicin induced CYP3A4 protein but not CYP3A5 protein in human hepatocytes (Schuetz et al., 1993). Since both donors in this study were genotyped as homozygous for the CYP3A5*3 allele (Katoh et al., 2004), further studies using chimeric mice generated from various donors are needed to clarify the induction of CYP3A5.
Rifampicin moderately induced murine Cyp3a11 mRNA and Cyp3a protein at a dose of 50 to 100 mg/kg/day for 2 to 4 days (Yanagimoto et al., 1997; Schuetz et al., 2000). TESOH is catalyzed by Cyp3a in mice. TESOH in uPA+/-/SCID mice was not changed by rifampicin treatment, whereas it was increased 2.4-fold in uPA-/-/SCID compared with the control (P < 0.01; data not shown), suggesting that rifampicin weakly induced Cyp3a. In both uPA+/-/SCID and uPA-/-/SCID mice, rifampicin had no effect on DEXOH, indicating that DEXOH was specific to human CYP3A but not to murine Cyp3a (Tomlinson et al., 1997).
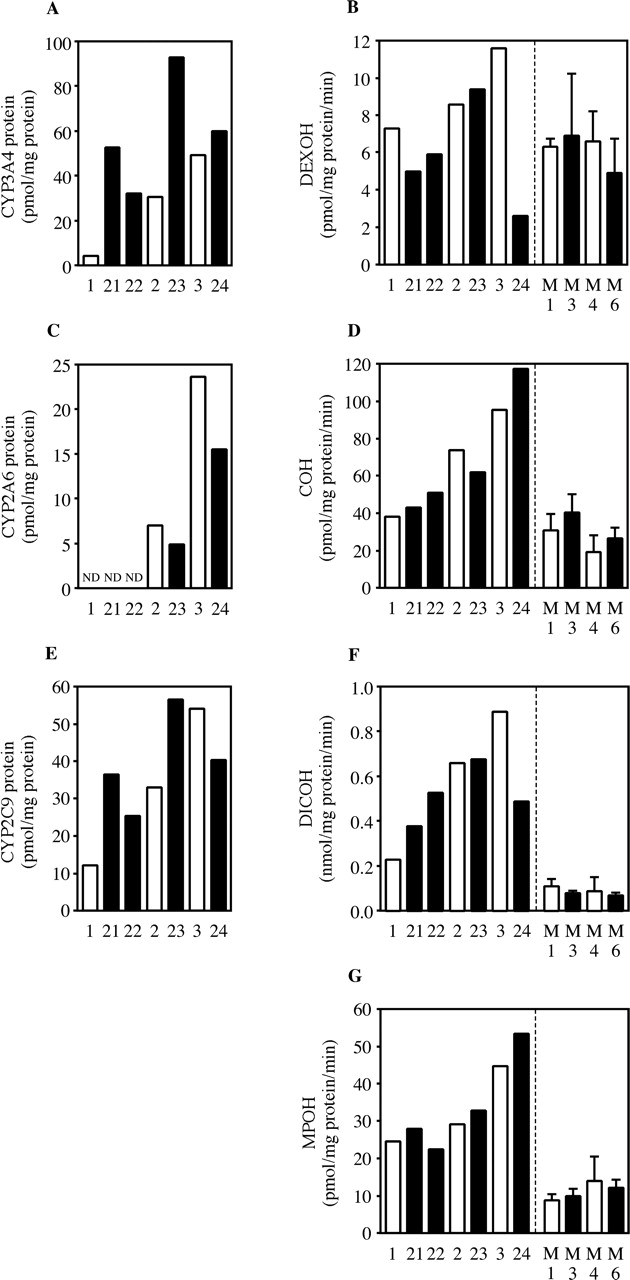
Changes of human CYP3A4, CYP2A6, CYP2C9, and CYP2C19 expressions in 3-MC-treated chimeric mice. Protein contents of human CYP3A4 (A), CYP2A6 (C), and CYP2C9 (E) in the chimeric mice were measured by Western blot analysis. DEXOH catalyzed by CYP3A4 (B), COH catalyzed by CYP2A6 (D), DICOH catalyzed by CYP2C9 (F), and MPOH catalyzed by CYP2C19 (G) were determined by HPLC. Open and closed columns are the values of the non- and 3-MC-treated chimeric mice, respectively. B, D, F, and G, columns of M1, M3, M4, and M6 represent the mean ± S.D. (n = 3). ND, not detected; M1, nontreated uPA+/-/SCID mouse; M3, 3-MC-treated uPA+/-/SCID mouse; M4, nontreated uPA-/-/SCID mouse; M6, 3-MC-treated uPA-/-/SCID mouse.
Rifampicin also induces the mRNAs and proteins of CYP2C8, CYP2C9, and CYP2C19 in human hepatocytes (Gerbal-Chaloin et al., 2001; Raucy et al., 2002). On the other hand, it has been reported that there were no changes in CYP2C8 protein (Edwards et al., 2003), CYP2C9 protein (Runge et al., 2000; Edwards et al., 2003), and CYP2C19 protein (Runge et al., 2000) following treatment with rifampicin in human hepatocytes or human liver slices. In addition, Raucy et al. (2002) demonstrated that the induction of CYP2C8 protein exhibited large interindividual variability, and HH954 hepatocytes in their report failed to respond to rifampicin treatment. They also reported that the induction of CYP2C19 protein exhibited large interindividual differences (5.7 ± 5.3-fold) (Raucy et al., 2002). In clinical practice, rifampicin may induce CYP2C-mediated metabolism and thus reduce the plasma concentration of CYP2C9 substrates such as warfarin and sulfonylurea antidiabetic drugs (Niemi et al., 2003). In the present study, rifampicin tended to cause a slight increase in CYP2C8 protein, CYP2C9 protein, DICOH catalyzed by CYP2C9, and MPOH catalyzed by CYP2C19 and showed interindividual differences. In the case of CYP2C19, it was difficult to estimate the induction, because the MPOH in donor B chimeric mice was lower. Donor B chimeric mice were genotyped as CYP2C19*1/CYP2C19*2 (Katoh et al., 2004), which would lead to a reduction of the enzyme activity (Bramness et al., 2003). In this study, the calculated induction ratio is an apparent value, since the hAlb concentrations are similar but not the same between non- and inducer-treated chimeric mice. Further investigations will be needed to clarify the induction potency by rifampicin of CYP2Cs in chimeric mice.
In relation to human CYP2A6, some in vitro reports using human liver slices and human hepatocytes exhibited the induction of CYP2A6 protein and COH by rifampicin treatment, respectively (Edwards et al., 2003; Madan et al., 2003); however, another study that used human hepatocytes showed no change in CYP2A6 mRNA and COH (Donato et al., 2000). Ethoxyresorufin O-dealkylase activity catalyzed by CYP1A2 was increased by treatment with rifampicin in human hepatocytes (Madan et al., 2003). In CYP1A2 and CYP2A6, rifampicin increased the mRNA, protein content, and enzyme activity 3-fold at most compared with the control (Edwards et al., 2003; Madan et al., 2003). Therefore, these results did not contradict those of the present study.
Following the exposure of 3-MC, human CYP1A2 protein and ethoxyresorufin O-dealkylase activities were increased in human hepatocytes, leading to the induction of CYP1A2 in humans (Donato et al., 1995; Runge et al., 2000). In the present study, human CYP1A2 mRNA and protein were induced by treatment with 3-MC in the chimeric mice, which was consistent with previous reports (Donato et al., 1995; Runge et al., 2000). 3-MC significantly increased the expression levels of human CYP1A1 mRNA in the present study. The CYP1A1 antibodies used in this study were very sensitive and could detect 25 fmol of the recombinant human CYP1A1, but the pooled human liver microsomes from BD Gentest did not show the band (data not shown). CYP1A1 is known as an isoform with low expression in normal human liver (Turesky et al., 1998). CYP1A1 protein in both non- and 3-MC-treated chimeric mice could not be detected. CYP1A1 protein in the liver of donor A may have been very low; therefore, human CYP1A1 proteins in donor A chimeric mice could not be detected, although human CYP1A1 mRNA could be detected.
There seemed to be some variability in the responses to these inducers in terms of the mRNA, protein, and enzyme activity. The difference is thought to be due to the interindividual variability of the chimeric mice, but the reasons are still unclear. We think that such variability could be overcome by increasing the number of chimeric mice, because it was also observed in studies using human hepatocytes.
As described above, it was demonstrated that P450 enzymes were induced in the chimeric mice with humanized liver. The expression of each murine P450 mRNA in chimeric mouse 3, which exhibited the highest hAlb concentration in this study, was no more than 5% compared with that in uPA+/-/SCID mice (data not shown). It was surmised that human P450s in the chimeric mice were induced by rifampicin or 3-MC treatment, but further study is needed to clarify the expression of human nuclear receptors and the transcriptional regulation mechanism in the chimeric mice.
In conclusion, human P450s expressed in chimeric mice with humanized liver respond to induction via treatment with rifampicin and 3-MC. At present, human hepatocytes are still a better model for investigating the induction of P450s, but the number of human hepatocytes that can be obtained from one donor may not be sufficient for the experimental purposes, and frequently none can be obtained at all. Using these chimeric mice, human hepatocytes could be made to proliferate easily at low cost. In some countries, including Japan, large amounts of human organ materials such as hepatocytes and microsomes are very difficult to obtain. In such cases, the lack of a stable supply of human liver is a serious problem. One of the advantages of the chimeric mice is that they could be used to proliferate human hepatocytes. In addition, this chimeric mouse line would be a better tool than any other experimental animal for estimating the in vivo induction potency in humans. It would be of interest to measure the pharmacokinetics of drugs in chimeric mice treated with a typical P450 inducer. This chimeric mouse line could be more useful than human hepatocytes for estimating the pharmacokinetics and drug metabolism in humans. We hope that this study will greatly contribute to future advances in studies of drug metabolism as well as drug development.
Acknowledgments
We acknowledge Brent Bell for reviewing the manuscript.
Footnotes
-
This work was supported by a Research on Advanced Medical Technology, Health, and Labor Sciences Research grant from the Ministry of Health, Labor, and Welfare of Japan.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.002600.
-
ABBREVIATIONS: P450, cytochrome P450; uPA, urokinase-type plasminogen activator; SCID, severe combined immunodeficient; 3-MC, 3-methylcholanthrene; TESOH, testosterone 6β-hydroxylase activity; hAlb, human albumin; RI, replacement index; PCR, polymerase chain reaction; HPLC, high-performance liquid chromatography; COH, coumarin 7-hydroxylase activity; DICOH, diclofenac 4′-hydroxylase activity; MPOH, S-mephenytoin 4′-hydroxylase activity; DEXOH, dexamethasone 6-hydroxylase activity; hGAPDH, human glyceraldehyde-3-phosphate dehydrogenase.
- Received October 6, 2004.
- Accepted March 10, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}