


Abstract
Although cytochrome P450 (CYP) 2D6 has been widely considered to be noninducible on the basis of human hepatocyte studies, in vivo data suggests that it is inducible by endo- and xenobiotics. Therefore, we investigated if the experimental conditions routinely used in human hepatocyte studies may be a confounding factor in the lack of in vitro induction of CYP2D6. Sandwich cultured human hepatocytes (SCHH) were preincubated with or without dexamethasone (100 nM) for 72 hours before incubation with 1 μM endogenous (cortisol or corticosterone) or exogenous (dexamethasone or prednisolone) corticosteroids. At 72 hours, CYP2D6 mRNA, protein, and activity were quantified by real-time quantitative polymerase chain reaction, quantitative proteomics, and formation of dextrorphan from dextromethorphan, respectively. In the absence of supplemental dexamethasone, CYP2D6 activity, mRNA, and protein were significantly and robustly (>10-fold) induced by all four corticosteroids. However, this CYP2D6 induction was abolished in cells preincubated with supplemental dexamethasone. These data show, for the first time, that CYP2D6 is inducible in vitro but the routine presence of 100 nM dexamethasone in the culture medium masks this induction. Our cortisol data are in agreement with the clinical observation that CYP2D6 is inducible during the third trimester of pregnancy when the plasma concentrations of cortisol increase to ∼1 μM. These findings, if confirmed in vivo, have implications for predicting CYP2D6-mediated drug-drug interactions and call for re-evaluation of regulatory guidelines on screening for CYP2D6 induction by xenobiotics. Our findings also suggest that cortisol may be a causative factor in the in vivo induction of CYP2D6 during pregnancy.
Introduction
Cytochrome P450 (CYP) 2D6 is a major drug-metabolizing enzyme that biotransforms ∼20% of all marketed drugs (Yu et al., 2004; Zanger et al., 2004). Although there is a significant interindividual variation attributed to genetic polymorphism, it is widely accepted that CYP2D6 protein is noninducible by xenobiotics (Benedetti, 2000). For this reason, during drug development a candidate drug is not screened for its CYP2D6 induction profile nor is such a profile required by regulatory agencies (FDA Website, 2015). However, there is considerable evidence that CYP2D6 is modestly inducible in vivo. For example, the oral clearance of the CYP2D6 substrates sparteine, dextromethorphan, and debrisoquine is increased (∼2-fold) in response to rifampin pretreatment (Eichelbaum et al., 1986; Leclercq et al., 1989; Kirby et al., 2011), that of desipramine is increased (∼2-fold) in response to phenobarbital pretreatment (Spina et al., 1996), and oral clearance of risperidone is increased (∼2-fold) in response to carbamazepine pretreatment (Ono et al., 2002). Moreover, the oral clearance of the prototypical CYP2D6 substrates metoprolol and dextromethorphan is significantly increased, by 3-fold and 2-fold, respectively, during pregnancy (Tracy et al., 2005; Ryu et al., 2015).
Despite the above in vivo evidence, CYP2D6 is either not inducible or only modestly inducible in the most commonly used in vitro system, the human hepatocytes (Benedetti, 2000; Dixit et al., 2007). Here we present data that challenge this dogma and demonstrate that CYP2D6 is indeed significantly and highly inducible by endo- and xenobiotics. We also show that the in vitro inducibility of CYP2D6 by xenobiotics in human hepatocytes is prevented by the culture conditions commonly used in these studies.
Materials and Methods
Chemicals and Reagents.
Cortisol, dexamethasone, prednisolone, corticosterone, dextromethorphan, and diazepam were purchased from Sigma-Aldrich (St. Louis, MO). Dextrorphan-d3 was purchased from Toronto Research Chemicals (Toronto, Canada). Midazolam, 1-OH-midazolam, and 1-OH-midazolam-d4 were purchased from Cerilliant Corporation (Round Rock, TX). Acetic acid (American Chemistry Society grade) and acetonitrile (mass spectrometry grade) were obtained from Fisher Scientific (Pittsburgh, PA). William’s medium E, Dulbecco’s phosphate-buffered saline, GlutaMAX-I, penicillin-streptomycin (10,000 IU/ml), insulin-transferrin-selenium, Trizol reagent, TaqMan reverse transcription reagents, universal Polymerase Chain Reaction Master Mix, CYP2D6, glyceraldehyde 3-phosphate dehydrogenase, and human β-glucuronidase Taqman primer-probe sets, and collagen-coated cell culture plates were purchased from Life Technologies/Thermo Fisher Scientific (Carlsbad, CA). Matrigel (growth-factor reduced, phenol red free) was obtained from BD Biosciences (San Jose, CA). In-solution trypsin digestion kit, BCA protein assay kit, iodoacetamide, and dithiothreitol were obtained from Pierce Biotechnology (Rockford, IL). The ProteoExtract Native Membrane Protein Extraction Kit was obtained from Calbiochem/MilliporeSigma (Temecula, CA). Synthetic signature peptides for CYP2D6 protein was obtained from New England Peptides (Boston, MA). The corresponding stable isotope-labeled internal standards were purchased from ThermoFisher Scientific (Rockford, IL). Deionized water was generated from a Q-Gard 2 Purification Pack water purifying system (EMDMillipore, Bedford, MA). Cryopreserved human hepatocytes from three premenopausal donors (Table 1) were provided by Corning (Tewksbury, MA). Collagen-coated cell culture plates were purchased from Life Technologies (Carlsbad, CA).
Demographic information of the hepatocyte donors
Treatment of Sandwich Cultured Human Hepatocytes or HepaRG Cells.
Proliferative-state HepaRG cells were expanded and differentiated in 75-cm2 flasks according to the provider’s protocol (Biopredic International, Saint-Grégoire, France). Differentiated HepaRG cells were trypsinized and seeded in 24-well plates and initially maintained for 48 hours in differentiation medium before being switched to dexamethasone (100 nM) containing or dexamethasone-free maintenance medium (prepared using William’s medium E supplemented with GlutaMAX-I, insulin-transferrin-selenium, and penicillin-streptomycin) for 72 hours. At the end of the maintenance period, medium was removed and the cells were treated for 72 hours with 1 μM of cortisol, corticosterone, dexamethasone, prednisolone, rifampin, or vehicle (0.9% methanol). Stock solutions of rifampin and the corticosteroids were made in 100% methanol. To compensate for possible depletion, induction medium (containing the test compounds) was renewed every 12 hours (Zhang et al., 2015).
Cryopreserved human hepatocytes were thawed and plated in collagen-coated 24-well plates at a density of ∼0.35 million viable cells/well and were overlaid, after 24 hours of plating, with Matrigel per supplier’s instructions. After 24 hours of Matrigel overlay, commercially available hepatocyte maintenance medium was substituted with dexamethasone (100 nM) containing or dexamethasone-free maintenance medium (composition as described above). At the end of the 72-hour maintenance period, sandwich cultured human hepatocytes (SCHH) were treated as described above.
Quantification of CYP2D6 and CYP3A Activity.
To quantify CYP2D6 activity in SCHH or HepaRG cells, the medium was aspirated at the end of induction period and cells were rinsed twice with prewarmed Dulbecco’s phosphate-buffered saline (DPBS). Then, cells were incubated for 1 hour with 5 μM dextromethorphan dissolved in serum-free William’s medium E. For measurement of CYP3A activity, HepaRG cells or SCHH were incubated for 1 hour with 2 μM midazolam dissolved in serum-free William’s medium E. For both CYP2D6 and CYP3A activity quantification, at the end of incubation period, supernatant was collected, and an equal volume of ice-cold acetonitrile containing 100 nM dextrophan-d3 (internal standard) and/or 1′-OH-midazolam-d4 (internal standard) was immediately added to the supernatant and vials were immediately sealed with airtight covers. Dextrorphan formation was quantified using an Agilent 1290 Infinity ultra-performance liquid chromatography (UPLC) system coupled to an Agilent 6400Triple Quad mass spectrometer (Agilent Technologies, Santa Clara, CA). An Acquity UPLC BEH [ethylene bridged hybrid (BEH)] C18 Column (1.7 μm, 2.1 mm × 50 mm; Waters Corporation, Milford, MA) was used with the mobile phase consisting of 0.1% acetic acid (v/v) in water (phase A) or 0.1% acetic acid (v/v) in acetonitrile (phase B) (total flow rate 0.4 ml min−1). Phase B percentage was linearly increased from 10% at 0–0.5 minutes to 40% at 0.8 minutes to 100% at 2.5–3.5 minutes followed by 10% at 3.51–4.51 minutes. Dextrorphan daughter ions 157 and 133 were detected using the dynamic multiple reaction monitoring mode with 38 and 26 V collision energy respectively. 1′-OH-midazolam formation was quantified by liquid chromatography–tandem mass spectrometry method as described previously (Shirasaka et al., 2013).
CYP2D6 mRNA and Protein Quantification.
For CYP2D6 mRNA quantification, cells were washed and lysed using Trizol reagent and total RNA was extracted, reverse transcribed, quantified and analyzed as described previously (Dixit et al., 2007). For protein quantification, total plasma membranes were extracted, quantified and analyzed as described previously (Kumar et al., 2015; Wang et al., 2015). This membrane fraction contains all cell membranes including the microsomes (unpublished data). Briefly, CYP2D6 signature peptide analysis was performed using Waters Xevo TQS tandem mass spectrometer coupled to Waters Acquity UPLC system (Waters, Hertfordshire, UK) operated in electrospray positive ionization mode. An Acquity UPLC BEH C18 Column (1.7 μm, 2.1 mm × 50 mm; Waters Corporation, Milford, MA) wa18s used with the mobile phase consisting of 0.1% acetic acid (v/v) in water (phase A) or 0.1% acetic acid (v/v) in acetonitrile (phase B) (total flow rate 0.3 ml min−1). Phase B percentage was linearly increased from 3% at 0–1.5 minutes to 15% at 3 minutes to 40% at 6 minutes to 80% at 6.1 minutes followed by 3% at 7.1–9 minutes. Daughter ions (m/z 861.3 and 974.4) of the target CYP2D6 peptide (m/z 673.9) were detected using multiple reaction monitoring mode with 17-V collision energy. Also daughter ions (m/z 869.3 and 982.4) of the heavy-labeled version of the target CYP2D6 peptide (GTTLITNLSSVLK) (m/z 677.9), which served as an internal standard, were detected using the same parameters.
Data Analysis.
Unless otherwise stated, data were expressed relative to the control as mean ± S.D. Data were compared with the corresponding controls (e.g., absence or presence of dexamethasone supplement). Unless otherwise stated, all experiments in SCHH or HepaRG cells were conducted in triplicates (treatment) or six determinations (control). For comparisons of treatments, one-way analysis of variance followed by post-hoc Dunnett’s multiple comparison was performed. The maximal CYP2D6 activity (Emax)(relative to control = 1), half maximal effective concentration (EC50), and baseline CYP2D6 activity (E0)(baseline value) of induction of CYP2D6 expression and activity was estimated by fitting the simple Emax model to the concentration-response data using nonlinear regression (Graphpad Prism 5.0, La Jolla, CA).
Results
Induction of CYP2D6 Activity in SCHH by Endogenous or Exogenous Corticosteroids in the Presence or Absence of Dexamethasone as a Medium Supplement.
Basal CY2D6 activity as measured by dextrorphan formation in SCHH in the presence of dexamethasone as a medium supplement was >5-fold higher compared with cells maintained in the absence of dexamethasone (Fig. 1). SCHH maintained in the culture medium supplemented with dexamethasone did not significantly induce CYP2D6 activity in response to treatment with 1 μM endogenous (cortisol, corticosterone) or exogenous corticosteroids (dexamethasone, prednisolone) (Fig. 1). In contrast, cells maintained in the dexamethasone-free cell culture medium showed robust >10-fold CYP2D6 induction when treated with corticosteroids. Interestingly, the magnitude of CYP2D6 induction (∼2-fold) by rifampin was modest and independent of the presence or absence of dexamethasone as a medium supplement.
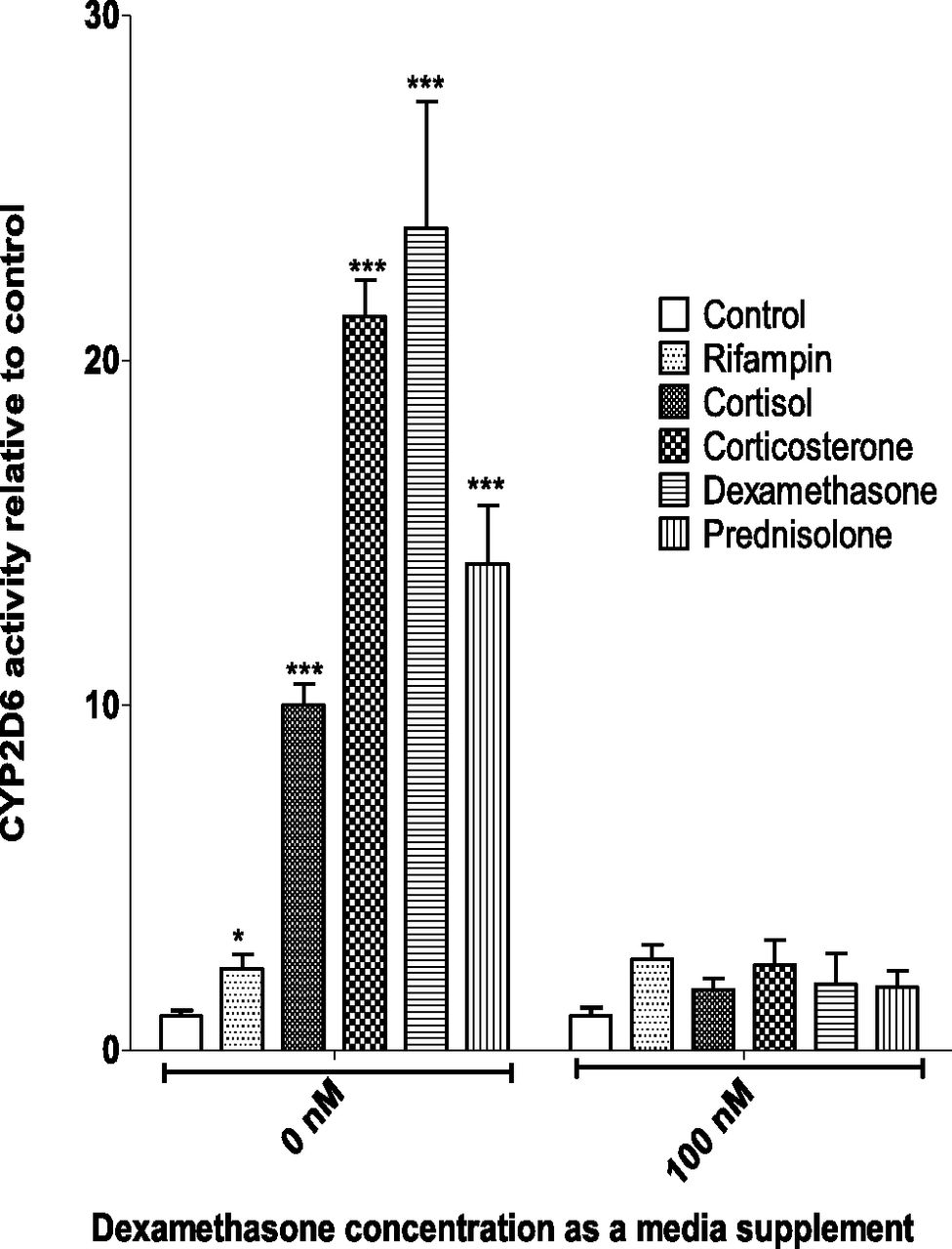
Induction of CYP2D6 activity in SCHH by endogenous or exogenous corticosteroids in the presence or absence of dexamethasone as a medium supplement. SCHH (Lot. 393) preincubated for 72 hours in the presence or absence of dexamethasone (100 nM) were treated with 1 μM corticosteroids or rifampin for 72 hours before quantifying CYP2D6 activity by incubating with dextromethorphan (5 μM for 1 hour) and measuring dextrorphan formation. Fold induction (mean ± S.D. of triplicates) was expressed relative to the respective control (mean ± S.D. of six determinations). CYP2D6 activity induced by 1 μM corticosteroid treatment of SCHH was abolished when dexamethasone was included as a medium supplement 72 hours prior to corticosteroid treatment. However, CYP2D6 activity in the control group of SCHH incubated with supplemental dexamethasone was 5.4-fold higher than the control group incubated without supplemental dexamethasone. Data were analyzed by one-way analysis of variance followed by Dunnett’s multiple comparison test. *P < 0.05; ***P < 0.001 compared with respective control.
CYP2D6 Induction in SCHH by Endogenous or Exogenous Corticosteroids as Measured by CYP2D6 mRNA, Protein, or Activity.
To determine the cause of the observed CYP2D6 induction, we measured CYP2D6 mRNA, protein, and activity in SCHH treated with 1 μM endogenous or exogenous corticosteroids. Interestingly, CYP2D6 mRNA, protein, and activity all showed robust induction in response to corticosteroid treatment (Fig. 2). Surprisingly, the magnitude of induction of CYP2D6 protein and activity was greater than that of mRNA. The reverse was true for 1 μM rifampin (Fig. 2). In all cases the magnitude of induction of CYP2D6 activity was similar to the induction of protein, indicating that the induction of CYP2D6 activity was not an artifact of the use of dextromethorphan as a CYP2D6 substrate (e.g., owing to inhibition of secondary metabolism of dextrorphan by the corticosteroids).

CYP2D6 induction in SCHH by endogenous or exogenous corticosteroids as measured by CYP2D6 mRNA, protein, or activity. SCHH (Lot. 393) maintained for 72 hours in dexamethasone-free incubation medium were treated with 1 μM corticosteroids or rifampin for 72 hours. At the end of treatment period, CYP2D6 mRNA, protein, and activity was quantified. Fold-induction (mean ± S.D. of triplicates) was expressed relative to control (mean ± S.D. of six determinations). CYP2D6 protein and activity were robustly induced in response to corticosteroid but not rifampin treatment (1 μM). However, the induction of CYP2D6 protein or activity by the corticosteroids was always greater than the induction of mRNA. The reverse was true with rifampin. Data were analyzed using one-way analysis of variance followed by Dunnett’s multiple comparison test. *P < 0.05; ***P < 0.001 compared with the control.
CYP2D6 Induction Potential in SCHH When Supplemental Dexamethasone Is Substituted with Physiologically Relevant Concentration of Cortisol.
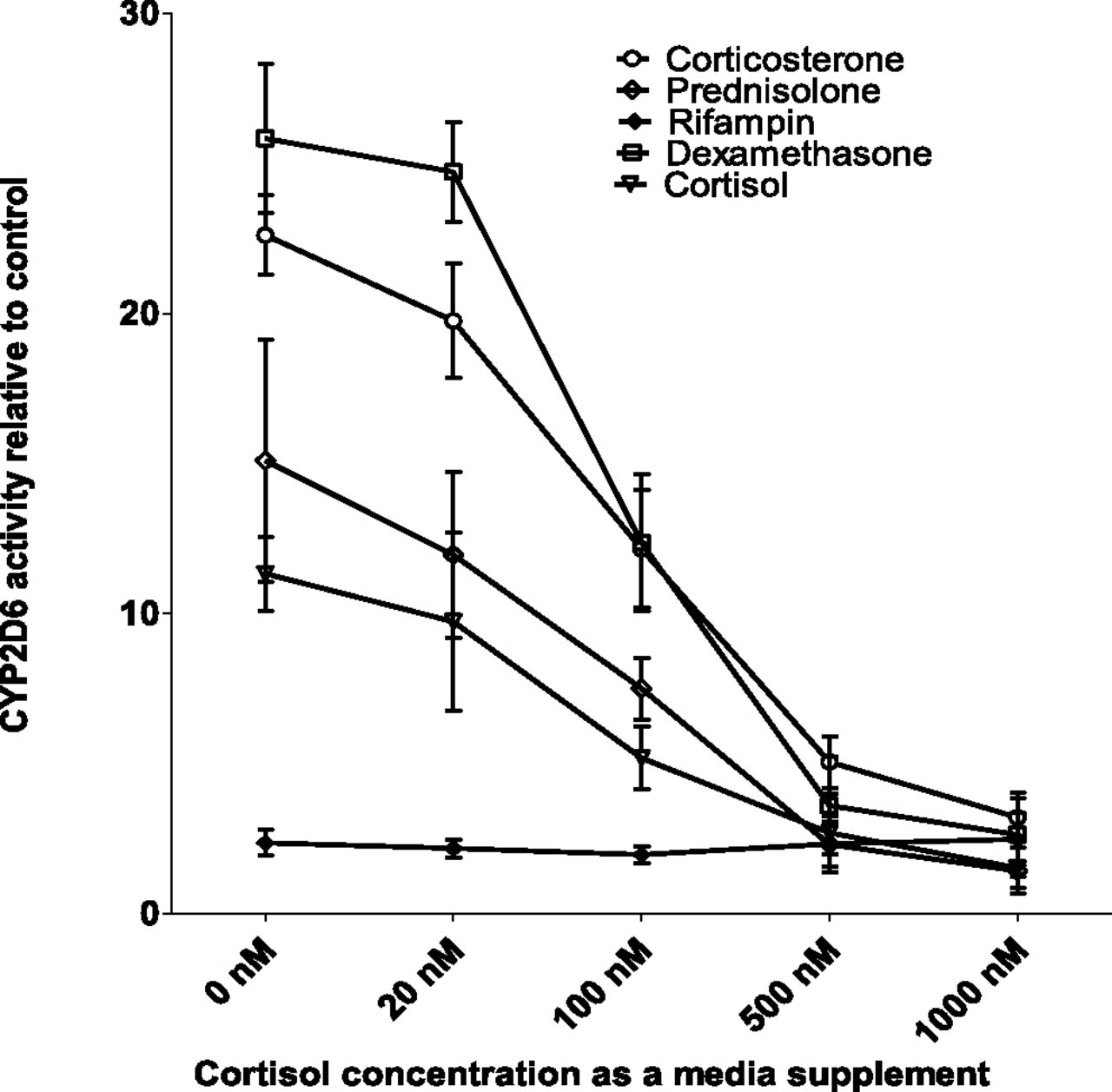
Since the presence of supplemental dexamethasone abolished CYP2D6 induction by corticosteroids, we tested if substitution of cortisol for dexamethasone had a similar effect. Physiologically relevant unbound plasma concentration of cortisol (20 nM) (Zhang et al., 2015), as a medium supplement, had a negligible effect on the magnitude of CYP2D6 induction by 1 μM corticosteroids (Fig. 3). However, increasing concentration of cortisol as a medium supplement reduced the ability of corticosteroids to induce CYP2D6 activity. At the highest concentration of cortisol (1 μM), the induction of CYP2D6 by the corticosteroids was abolished, mirroring the effect of 100 nM dexamethasone. In contrast, the induction by rifampin was unaffected by supplemental cortisol concentration (Fig. 3).

CYP2D6 induction in SCHH when supplemental dexamethasone is replaced with physiologically relevant concentration of cortisol. SCHH (Lot. 393) preincubated for 72 hours in the presence or absence of cortisol (20, 100, 500, 1000 nM) were treated with 1 μM corticosteroids or rifampin for 72 hours before quantification of CYP2D6 activity by incubation with dextromethorphan (5 μM for 1 hour) and measurement of dextrorphan formation. Fold induction (mean ± S.D. of triplicates) was expressed relative to the respective control (mean ± S.D. six determinations). Induction of CYP2D6 in SCHH by 1 μM corticosteroids (but not by rifampin) was reduced, in a concentration-dependent manner, by inclusion of cortisol as a medium supplement 72 hours prior to treatment with the corticosteroids. Notably, physiologically relevant unbound plasma concentration of cortisol (20 nM) slightly (but not significantly) decreased the magnitude of CYP2D6 induction by the corticosteroids. However, CYP2D6 activity in the control group of SCHH incubated in the presence of supplemental cortisol increased in a concentration-dependent manner (up to 5-fold at 1 μM) compared with the control group incubated in the absence of supplemental cortisol.
Hepatocyte Morphology in the Presence or Absence of Dexamethasone or Cortisol as a Culture Medium Supplement.
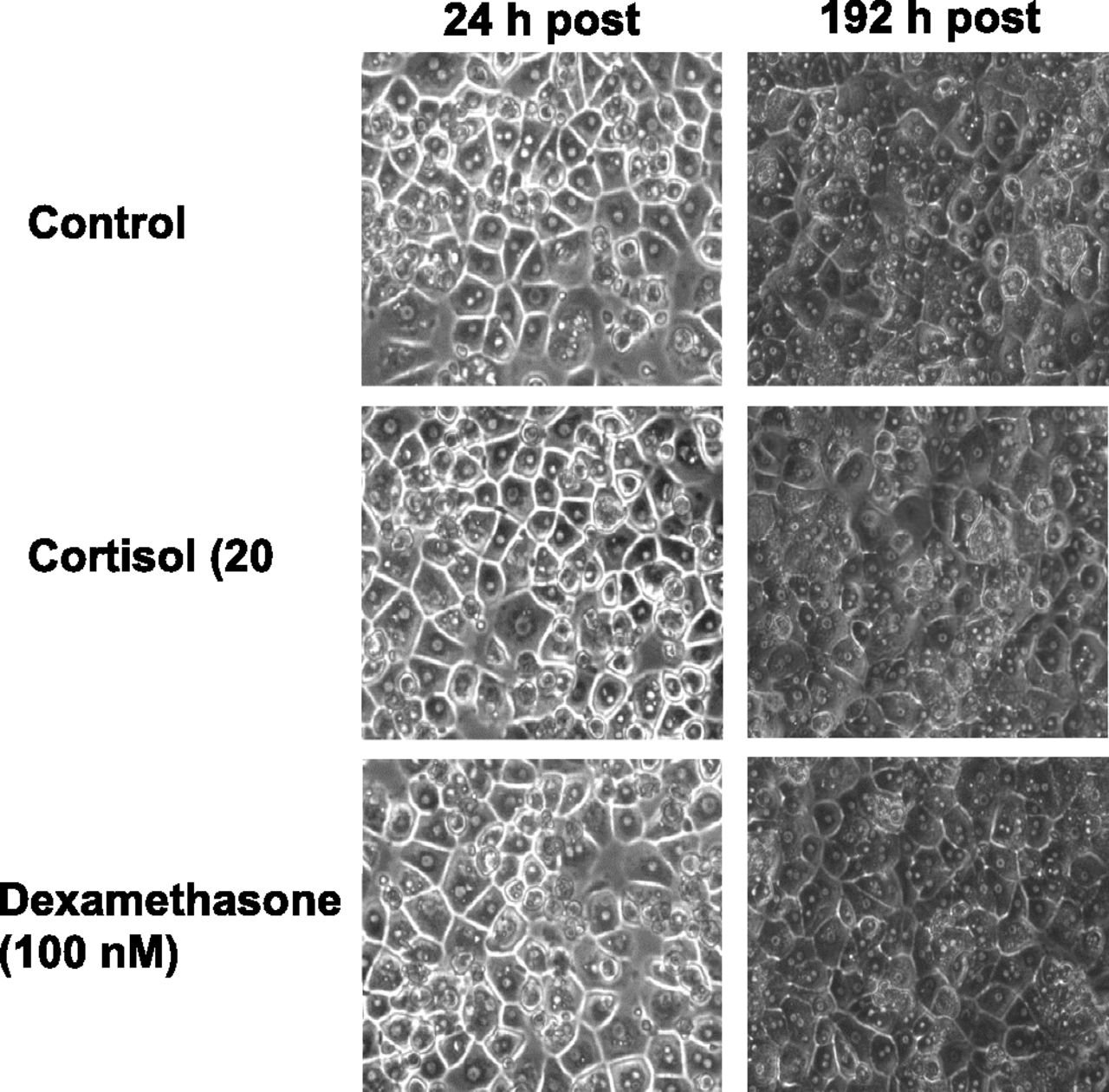
Since CYP2D6 was robustly induced in SCHH by corticosteroids only when SCHH were exposed to dexamethasone-free medium, we asked if the absence of dexamethasone (100 nM) had any deleterious effect on hepatocyte morphology. SCHH maintained in the presence or absence of dexamethasone as a medium supplement exhibited identical cuboidal cell morphologies (Fig. 4). Moreover, substitution of dexamethasone by a physiologically relevant concentration of cortisol (20 nM) as a medium supplement did not affect cell morphology (Fig. 4).

Hepatocyte morphology in the presence or absence of dexamethasone or cortisol as a culture medium supplement. SCHH (Lot. 393) were maintained for 192 hours (total culture time equivalent to treatment of SCHH with corticosteroids, i.e., 48 hours of acclimatization period plus 72 hours of preincubation period plus 72 hours of treatment period) in the presence or absence of cortisol (20 nM) or dexamethasone (100 nM) as a medium supplement and photographed at 24 hours and 192 hours at 20× magnification. Removal of dexamethasone (100 nM) or substitution with cortisol (20 nM) as cell culture medium supplement was not detrimental to hepatocyte morphology. Photographs taken at 192 hours are darker compared with 24 hours owing to the low background light intensity.
CYP3A Induction in the Presence or Absence of Dexamethasone as a Culture Medium Supplement.
Unlike induction of CYP2D6 (Fig. 1), the presence of dexamethasone as a medium supplement did not abolish CYP3A induction in response to treatment with 1-μM concentration of cortisol or dexamethasone. However, the presence of dexamethasone supplement showed significantly lower magnitude of CYP3A induction by the corticosteroids compared with control (Fig. 5). Furthermore, when compared with control, the presence or absence of cortisol (20 nM) as a medium supplement did not show any significant decrease in the magnitude of CYP3A induction in response to treatment with 1 μM cortisol or dexamethasone.

Effects of presence or absence of dexamethasone as a culture medium supplement on CYP3A induction. SCHH (Lot. 393) were maintained for 72 hours in dexamethasone (100 nM) or cortisol (20 nM) -containing or -free incubation medium. Cells were then treated with 1-μM concentrations of corticosteroids for 72 hours in the presence or absence of dexamethasone (100 nM) or cortisol (20 nM). At the end of treatment period, CYP3A activity (midazolam hydroxylation) was quantified. Fold induction (mean ± S.D. of triplicates) is expressed relative to respective control (mean ± S.D. six determinations). Unlike CYP2D6 situation, CYP3A induction in response to cortisol or dexamethasone (1 μM) treatment was significantly decreased but not abolished by the presence of supplemental dexamethasone. Data were analyzed by one-way analysis of variance followed by Dunnett’s multiple comparison test. *P < 0.05 compared with respective control.
Concentration-Dependent Induction of CYP2D6 mRNA and Activity in SCHH by Endogenous or Exogenous Corticosteroids.
In the absence of dexamethasone as a medium supplement, all the corticosteroids induced CYP2D6 mRNA expression and activity in SCHH in a concentration-dependent manner (Fig. 6, A and B). In contrast, although rifampin treatment induced CYP2D6 mRNA in a concentration-dependent manner and to an extent similar to the corticosteroids, it induced CYP2D6 activity to a much smaller extent (Fig. 6, A and B). Dexamethasone and corticosterone were the most potent inducers of CYP2D6 followed by prednisolone and cortisol.

Concentration-dependent induction of CYP2D6 mRNA and activity in SCHH by endogenous or exogenous corticosteroids. SCHH (Lot. 393) cells maintained for 72 hours in dexamethasone-free incubation medium were treated with varying concentrations of the steroids for 72 hours. At the end of treatment period, CYP2D6 mRNA (A) and activity (B) was quantified. Data are presented for a representative lot out of three lots. Data are expressed as fold-induction (mean ± S.D. of triplicates) relative to the respective control (mean ± S.D. six determinations). Emax (relative to control = 1) and EC50 (estimate and 95% confidence interval) were estimated by fitting the simple Emax model to the individual data (see Table 2).
CYP2D6 Induction in HepaRG Cells.
The pattern and concentration-dependent induction of CYP2D6 mRNA and activity by corticosteroids and rifampin in HepaRG cells in the absence and presence of supplemental dexamethasone were similar to those observed in SCHH. However, the magnitude of induction of both CYP2D6 mRNA expression and activity was smaller (Fig. 7, A and B). Not surprisingly, batch-to-batch variability in CYP2D6 induction in HepaRG cells was much smaller than that with SCHH.

CYP2D6 induction in HepaRG cells. HepaRG cells maintained for 72 hours in dexamethasone-free incubation medium were treated with varying concentrations of individual steroids or rifampin for 72 hours. At the end of treatment period, CYP2D6 mRNA (A) and activity (B) was quantified. Fold-induction is expressed relative to control and data are expressed as mean ± S.D. from a single representative lot of HepaRG cells. Emax (relative to control = 1) and EC50 (estimate and 95% confidence interval) were estimated by fitting the simple Emax model to the individual data (see Table 2).
Quantification of Emax and EC50 of Induction of CYP2D6 mRNA and Activity by Corticosteroid Treatments in SCHH and HepaRG cells.
Although HepaRG cells displayed an identical CYP2D6 induction pattern in response to corticosteroid treatments, the magnitude of CYP2D6 induction in HepaRG cells was smaller compared with SCHH. To analyze this difference in response, Emax (relative to control = 1) values generated using SCHH and HepaRG were compared. Emax values for both CYP2D6 activity and mRNA were consistently and significantly higher in SCHH (unpaired t test) for all the corticosteroid treatments compared with the respective HepaRG values (Table 2).
Potency (EC50) and efficacy (Emax) of corticosteroids and rifampin to induce CYP2D6 activity and mRNA in SCHH and HepaRG cells
SCHH (Lots 292, 299, and 393) or HepaRG cells maintained for 72 hours in dexamethasone-free incubation media (100 nM) were treated with varying concentrations of individual steroids for 72 hours. At the end of treatment period, CYP2D6 mRNA and activity were quantified. Emax values are fold-induction relative to control = 1. Data are expressed as population mean ± S.D. from three different lots of human hepatocytes or HepaRG cells.
Discussion
Human hepatocyte cultures are known to lose their basal metabolic activity in a time-dependent manner (Guzelian et al., 1977; Pitot and Sirica, 1980; Guguen-Guillouzo and Guillouzo, 1983). To maintain enzyme activity (Nawa et al., 1986), cells are routinely cultured in the presence of serum (usually bovine) that contains physiologically relevant concentrations of all the endogenous hormones on the assumption that presence of these hormones will enhance cell survival (Laishes and Williams, 1976; Engelmann et al., 1985) and maintain enzyme activity. To further enhance enzyme activity, supplementing the culture medium with the synthetic glucocorticoid dexamethasone (100 nM) has become a routine and established culturing method (Schaefer et al., 2012). Although it has long been recognized that addition of dexamethasone may induce certain metabolic enzymes, this complication has been deemed acceptable (Wrighton et al., 1985; Gonzalez et al., 1986; Arlotto et al., 1987). Here we show for the first time that addition of dexamethasone, as a supplement to the culture medium, abolishes the ability of endogenous and exogenous corticosteroids to induce CYP2D6 in SCHH.
Absence of dexamethasone as a medium supplement resulted in robust induction of CYP2D6 in SCHH by both endogenous and exogenous corticosteroids (Fig. 1). However, the presence of supplement dexamethasone abolished this induction, which is probably attributable to the fact that CYP2D6 was already significantly induced by the supplemental dexamethasone. This assertion is supported by two observations. First, that the basal CYP2D6 activity was higher in the presence versus absence of supplemental dexamethasone. Second, the concentration of supplemental dexamethasone is comparable to the dexamethasone EC50 for induction of CYP2D6 activity in the absence of supplemental dexamethasone (Table 2). In contrast to the corticosteroids, 1 μM rifampin induced CYP2D6 activity only modestly (∼2-fold) and this magnitude of induction was similar whether dexamethasone was included as a medium supplement. These findings suggest differences in mechanisms of induction of CYP2D6 activity between rifampin and the corticosteroids.
To ensure that the observed induction of CYP2D6 activity was not an artifact (inhibition of secondary metabolism of dextrorphan by the corticosteroids could be misinterpreted as induction of CYP2D6 activity), we measured CYP2D6 mRNA, protein, and activity in SCHH treated with 1 μM endogenous or exogenous corticosteroids in the absence of dexamethasone as a medium supplement. Surprisingly, whereas the magnitude of induction of CYP2D6 activity and protein by the corticosteroids was similar, this induction was much greater than that of CYP2D6 mRNA. This is in contrast to what is observed with other cytochrome P450 enzymes (Dixit et al., 2007; Papageorgiou et al., 2013). The reverse was true of CYP2D6 induction by rifampin. Rifampin induced CYP2D6 mRNA to a greater extent (∼7-fold) than either CYP2D6 protein or activity. Consistent with these in vitro data, rifampin also induces in vivo CYP2D6 activity by ∼2-fold (Kirby et al., 2011). Our in vitro data speak to differences in mechanisms of induction of CYP2D6 activity between rifampin and the corticosteroids. Therefore, our data suggest that although corticosteroids appear to induce CYP2D6 activity by increasing CYP2D6 transcripts, other mechanisms such as stabilization of CYP2D6 mRNA or protein cannot be discounted. It is also possible, though less probable, that by day 3 of corticosteroid treatment, CYP2D6 mRNA levels had already peaked and declined (owing to increased degradation rate of CYP2D6 mRNA by the corticosteroids), whereas CYP2D6 protein and activity levels peaked on day 3 (owing to the longer half-life of the protein versus mRNA). Further studies need to be conducted to elucidate the exact molecular mechanisms of CYP2D6 induction by corticosteroids.
Dexamethasone is a potent exogenous corticosteroid and therefore its inclusion in human hepatocyte cultures is not representative of physiology. In contrast, cortisol is an endogenous circulating corticosteroid. We found that supplemental dexamethasone could be substituted with cortisol, at its circulating unbound plasma concentrations (∼20 nM) found in men and nonpregnant women (Zhang et al., 2015), without affecting the morphology of the cells (Fig. 4) or the ability of endogenous and exogenous corticosteroids to induce CYP2D6. However, increasing the supplemental cortisol concentration reduced CYP2D6 induction by the endogenous and exogenous corticosteroids. As was the case with supplemental dexamethasone, supplemental 1 μM cortisol abolished CYP2D6 induction by the endogenous and exogenous corticosteroids. As observed before (Fig. 1), the magnitude of CYP2D6 induction by rifampin was not affected by the concentration of supplemental cortisol (Fig. 3).
To ensure that, in the absence of supplemental dexamethasone, cytochrome P450 enzyme functionality of the SCHH was not compromised, the activity of well-studied enzymes, CYP3A4/5, was used as a control. The absence of supplemental dexamethasone did not affect the robust induction of CYP3A activity by 1 μM cortisol or dexamethasone (Fig. 5). Unlike induction of CYP2D6 activity, the inclusion of supplemental dexamethasone significantly reduced, but did not abolish, induction of CYP3A activity by these corticosteroids. These data on CYP3A induction by 1 μM corticosteroid treatment are in agreement with previous reports (Meunier et al., 2000; Pascussi et al., 2001; Ringel et al., 2002). This differential effect of supplemental dexamethasone on CYP3A and CYP2D6 induction in response to corticosteroids is probably attributable to differences in the mechanisms of induction of these enzymes.
We have previously shown that the pattern of induction of CYP3A activity in HepaRG cells by cortisol (in the absence of supplemental dexamethasone) is similar to that observed in SCHH (Zhang et al., 2015). Therefore, we determined if the same was true with respect to CYP2D6 induction. Indeed, this was the case (Fig. 7). However, the Emax (relative to control = 1) of CYP2D6 induction (both mRNA and activity) by the corticosteroids was lower in HepaRG cells compared with SCHH.
Although this is the first report that CYP2D6 is induced by endogenous and exogenous corticosteroids, there is a body of evidence that CYP2D6 is induced during human pregnancy (Wadelius et al., 1997). For instance, oral clearance of the CYP2D6 substrate metoprolol is increased by ∼3-fold during the third trimester (Hogstedt et al., 1985; Ryu et al., 2015). Likewise, oral clearance of clonidine and paroxetine (also CYP2D6 substrates) is increased by ∼2-fold during pregnancy (Buchanan et al., 2009; Ververs et al., 2009; Claessens et al., 2010). Also the urinary ratio of dextromethorphan to dextrorphan in pregnant women decreases in a gestational age–dependent manner, indicating a ∼2-fold increase in CYP2D6 activity (Tracy et al., 2005). The plasma concentration of cortisol during pregnancy increases with gestational age and is ∼1 μM during the third trimester (Zhang et al., 2015). Our data also show significant induction of CYP2D6 activity in SCHH and HepaRG cells by 1 μM cortisol when dexamethasone is absent as a medium supplement. Thus, our data suggest that CYP2D6 may be induced in pregnancy owing to the elevated cortisol concentration present in the plasma. Others have suggested that CYP2D6 is induced in pregnant CYP2D6-humanized mice as a result of elevated hepatic concentrations of all-trans retinoic acid (Koh et al., 2014). It is possible that multiple mechanisms may operate in human pregnancy to induce CYP2D6 activity. Interestingly, whereas cortisol induced CYP2D6 and CYP3A4 proteins in HepaRG, it did not induce other enzymes such as CYP1A2 and cytochrome P450-reductase (data not shown). Therefore, in the absence of dexamethasone supplementation, cortisol is not a nonspecific inducer of all hepatic enzymes. Moreover, since in vivo CYP1A2 activity is repressed during pregnancy (Tracy et al., 2005), mechanisms other than elevation in cortisol plasma concentrations probably caused this repression.
CYP3A and CYP2D6 activities are both induced during pregnancy (Tracy et al., 2005). On the basis of human hepatocytes and HepaRG cell data, we have speculated that elevated plasma concentrations of cortisol are the probable cause for the induction of CYP3A activity during pregnancy (Zhang et al., 2015). Here we report that induction of CYP2D6 activity during pregnancy may also be caused by these elevated cortisol plasma concentrations. Such parallel in vitro observations suggest possible similarity in the mechanisms of induction of these two proteins. Low concentrations of glucocorticoids are known to induce CYP3A expression and activity via the glucocorticoid receptor, whereas supraphysiological concentrations of glucocorticoids induce CYP3A4 via the pregnane X receptor (Pascussi et al., 2001). Our observation that CYP2D6 mRNA was induced by corticosteroids and rifampin suggests a possible role of glucocorticoid receptor or pregnane X receptor or both in the induction of CYP2D6 activity. Studies to investigate the molecular mechanisms by which CYP2D6 is induced by corticosteroids are underway in our laboratory.
In conclusion, our data show for the first time that CYP2D6 is inducible by both endogenous and exogenous corticosteroids. These findings, if confirmed in vivo, have direct implications for CYP2D6 mediated drug-drug interactions and call for re-evaluation of regulatory guidelines on screening for CYP2D6 induction by xenobiotics. Moreover, our data call for further studies to determine if cortisol is a causative factor in the in vivo induction of CYP2D6 activity during pregnancy. Detailed studies will need to be conducted to determine the molecular mechanisms by which CYP2D6 is induced by corticosteroids. In this regard, HepaRG appear to be an excellent model to elucidate the molecular mechanisms by which corticosteroids induce CYP2D6 activity.
Acknowledgments
The authors thank Robert D. Pelletier for providing dextrorphan-d3, Dr. Bhagwat Prasad for his help in optimizing the CYP2D6 proteomics assay, and Gabriela Patilea-Vrana for her help in statistical analysis.
Authorship Contributions
Participated in research design: Farooq, Kelly, Unadkat.
Conducted experiments: Farooq.
Performed data analysis: Farooq.
Wrote or contributed to the writing of the manuscript: Farooq, Kelly, Unadkat.
Footnotes
- Received December 29, 2015.
- Accepted March 9, 2016.
This work was supported by the National Institutes of Health National Institute on Drug Abuse [Grant P01DA032507].
Abbreviations
- CYP
- cytochrome P450
- EC50
- half maximal effective concentration
- E0
- baseline CYP2D6 activity
- Emax
- maximal CYP2D6 activity
- EC50
- half maximal effective concentration
- SCHH
- sandwich cultured human hepatocytes
- UPLC
- ultra-performance liquid chromatography
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}