


Abstract
Saquinavir (SQV) is a protease inhibitor widely used for the treatment of human immunodeficiency virus (HIV) infection. We profiled SQV metabolism in mice using a metabolomic approach. Thirty SQV metabolites were identified in mouse feces and urine, of which 20 are novel. Most metabolites observed in mice were recapitulated in human liver microsomes. Among these novel metabolites, one α-hydroxyaldehyde produced from SQV N-dealkylation was noted and verified for the first time. Meanwhile, the corresponding product (3S)-N-tert-butyldecahydro-isoquinoline-3-carboxamide and its further metabolites were identified in mouse urine. The α-hydroxyaldehyde pathway was confirmed by using semicarbazide as a trapping reagent as well. Using recombinant cytochrome P450 (CYP450) isoenzymes and Cyp3a-null mice, CYP3A was identified as the dominant enzyme contributing to the formation of α-hydroxyaldehyde. This study enhances our knowledge of SQV metabolism, which can be used for predicting drug-drug interactions and further understanding the mechanism of adverse effects associated with SQV.
Introduction
Saquinavir (SQV) is the first protease inhibitor approved by the U.S. Food and Drug Administration (FDA) for the treatment of human immunodeficiency virus (HIV) infection. SQV alone suffers from a low bioavailability, as ∼4% in the fed and down to 1% in the fasted state (Noble and Faulds, 1996). In general, the low bioavailability is attributed to the combined effects of limited absorption (approximately 30%) and extensive metabolism mediated by CYP3A in the liver and intestine (Vella and Floridia, 1998). Moreover, SQV has been identified as a substrate of P-glycoprotein, an efflux transporter (Kim et al., 1998). Because of its low bioavailability, SQV is commonly used in combination with low-dose ritonavir (RTV) (Plosker and Scott, 2003), an antiretroviral drug that has a potent inhibitory effect on both CYP3A and P-glycoprotein (Eagling et al., 1997). Both the SQV and the RTV (SQV/r)-boosted regimens have favorable tolerability profiles against HIV infection, with high effectivity and selectivity (Vella and Floridia, 1998; Plosker and Scott, 2003). However, various adverse effects related to SQV have been reported in clinical practice, such as diarrhea (20%), nausea (11%), and life-threatening liver injuries (5.7%) (Sulkowski et al., 2000). The exact mechanisms of the SQV-related adverse events are largely unknown.
At present, the generation of reactive metabolites is still regarded as an important cause of drug-induced toxicity (Baillie, 2006; Guengerich and MacDonald, 2007; Argikar et al., 2010). The reactive metabolites, such as aldehyde, epoxide, and quinone, can bring about various adverse effects. (O’Brien et al., 2005; Tang and Lu, 2010). For instance, the toxic metabolite atropaldehyde from felbamate is responsible for felbamate-induced hepatotoxicity (Dieckhaus et al., 2002). Atropaldehyde inhibits the detoxifying enzymes aldehyde dehydrogenase and glutathione S-transferase, which results in an accumulation of reactive species and toxicity (Kapetanovic et al., 2002).
To date, investigations of the metabolism of SQV have been mainly confined to in vitro studies. Fitzsimmons and Collins (1997) identified seven metabolites from incubations in human liver microsomes (HLM) and human intestine microsomes, including six monohydroxylated SQV metabolites and one dihydroxylated SQV. More recent studies have revealed monohydroxylated, dihydroxylated, and monohydroxylated plus dehydrogenated SQV metabolites from hepatic microsomal incubation (Eagling et al., 2002). Unfortunately, limited information is available on the bioactivation of SQV.
In this study, we profile the metabolism of SQV in mice by use of a metabolomic approach, which has been proven to be a powerful tool in the study of drug metabolism (Chen et al., 2006; Li et al., 2010, 2011b). Using this approach, we identified 10 known and 20 novel SQV metabolites; one of these, α-hydroxyaldehyde, a potential reactive metabolite, was observed and characterized. CYP3A4 was determined as the primary enzyme involved in the formation of this aldehyde. These findings could be used for prediction of drug-drug interactions and in further studies of SQV-related adverse effects from the aspect of metabolic bioactivation.
Materials and Methods
SQV [(2S)-N-[(2S,3R)-4-[(3S)-3-(tert-butylcarbamoyl)-decahydroisoquinolin-2-yl]-3-hydroxy-1-phenylbutan-2-yl]-2-(quinolin-2-ylformamido)butanediamide] and RTV (1,3-thiazol-5-ylmethyl N-[(2S,3S,5S)-3-hydroxy-5-[(2S)-3-methyl-2-{[methyl({[2-(propan-2-yl)-1,3-thiazol-4-yl]methyl})carbamoyl]amino}butanamido]-1,6-diphenylhexan-2-yl]carbamate) were supplied by the National Institutes of Health AIDS Research and Reference Reagent Program. Semicarbazide and NADPH were obtained from Sigma-Aldrich (St. Louis, MO). Recombinant human CYP450 (EasyCYP Bactosomes) and HLM were purchased from XenoTech (Lenexa, KS). All solvents for liquid chromatography and mass spectrometry were of the highest grade commercially available.
Animal Treatments and Sample Preparation.
Wild-type (WT) and Cyp3a-null mice were employed in this study. All mice (2–4 months old, male) were maintained under a standard 12-hour dark/light cycle with water and chow provided ad libitum. Handling was in accordance with study protocols approved by the University of Kansas Medical Center Institutional Animal Care and Use Committee. WT and Cyp3a-null mice were treated (PO) with SQV (150 mg/kg), and they were housed separately in metabolic cages for 18 hours.
Urine and feces were collected for analysis. Briefly, urinary samples were prepared by mixing 40 µl of urine with 160 µl of 50% acetonitrile and were centrifuged at 16,000g for 10 minutes. Feces were homogenized in water (100 mg feces in 1000 µl of H2O). Subsequently, 200 µl of acetonitrile was added to 200 µl of the resulting mixture, followed by centrifugation at 16,000g for 10 minutes. The supernatant was transferred to a new Eppendorf vial for a second centrifugation step (16,000g for 10 minutes). Each supernatant was transferred to an autosampler vial, and 5.0 µl was injected to a system (Waters Corporation, Milford, MA) combining ultraperformance liquid chromatography (UPLC) and quadrupole time-of-flight mass spectrometry (QTOFMS) for analysis.
Metabolism of SQV In Vitro.
Incubations were conducted in 1X phosphate-buffered saline (1X PBS, pH 7.4) containing 50 µM SQV, 0.2 mg HLM, or 2 pmol of each cDNA-expressed P450 enzyme (control, CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4) in a final volume of 190 µl. After 5 minutes of preincubation at 37°C, the reaction was initiated by adding 10 µl of 20 mM NADPH (final concentration 1.0 mM) and continued for 30 minutes with gentle shaking. Incubations in the absence of NADPH served as controls. Coincubations of SQV (50 µM) and RTV (2.0 µM) in HLM were performed to determine the role of CYP3A in the metabolism of SQV. Reactions were terminated by 200 µl of ice-cold acetonitrile, vortexing for 30 seconds, and centrifugation at 16,000g for 10 minutes. Each supernatant was transferred to an autosampler vial, and 5.0 µl was injected to the UPLC-QTOFMS system for metabolite analysis. Incubations were conducted in duplicate for cDNA-expressed P450 enzyme, and in triplicate for HLM and inhibitory experiments.
Trapping α-Hydroxyaldehyde (M22′) Using Semicarbazide.
The αhydroxyaldehyde (M22′) was trapped with semicarbazide in our study. The experiments were performed in 1X PBS containing 50 µM SQV, 0.2 mg HLM, and 10 µl of 50 mM semicarbazide (dissolved in H2O, final concentration 2.5 mM) in a final volume of 190 µl. After 5 minutes of preincubation at 37°C, the reaction was initiated by the addition of 10 µl of 20 mM NADPH (final concentration 1.0 mM) and continued for 45 minutes with gentle shaking. Incubations without NADPH were conducted as controls. The reactions were quenched by adding 200 µl of ice-cold acetonitrile. The mixture was vortexed for 1 minute and centrifuged at 16,000g for 10 minutes. The supernatant was transferred to an autosampler vial, and 5.0 µl was injected to the UPLC-QTOFMS system for analysis.
UPLC-QTOFMS Analyses.
The separation of SQV and its metabolites was achieved using a 100 mm × 2.1 mm (Acquity 1.7 µm) UPLC ethylene bridged hybrid (BEH) C-18 column (Waters, Milford, MA). The flow rate of the mobile phase was 0.3 ml/min with a gradient ranging from 2% to 98% aqueous acetonitrile containing 0.1% formic acid in a 10-minute run. QTOFMS was operated in a positive mode with electrospray ionization. The source temperature and desolvation temperature were set at 120°C and 350°C, respectively. Nitrogen was applied as the cone gas (10 l/h) and desolvation gas (700 l/h). Argon was applied as the collision gas. QTOFMS was calibrated with sodium formate and monitored by the intermittent injection of lock mass leucine enkephalin in real time, generating a reference ion at m/z 556.2771. The capillary voltage and the cone voltage were set at 3.5 kV and 35 V, respectively. The structures of metabolites of SQV were elucidated by tandem mass spectrometry fragmentation with collision energy ramp ranging from 10 to 40 V.
Data Analysis.
Mass chromatograms and mass spectra were acquired by MassLynx software (Waters) in centroid format from m/z 50 to 1000. Centroid and integrated mass chromatographic data were processed by MarkerLynx software (Waters) to generate a multivariate data matrix. The corresponding data matrices were then exported into SIMCA-P+12 (Umetrics, Kinnelon, NJ) for multivariate data analysis. Orthogonal projection to latent structures-discriminant analysis (OPLS-DA) was conducted on Pareto-scaled data. For the chemometric analysis, urine matrix data were processed from m/z 50 to 320, and feces data from m/z 320 to 900.
Results
Profile SQV Metabolism in Mice Using a Metabolomic Approach.
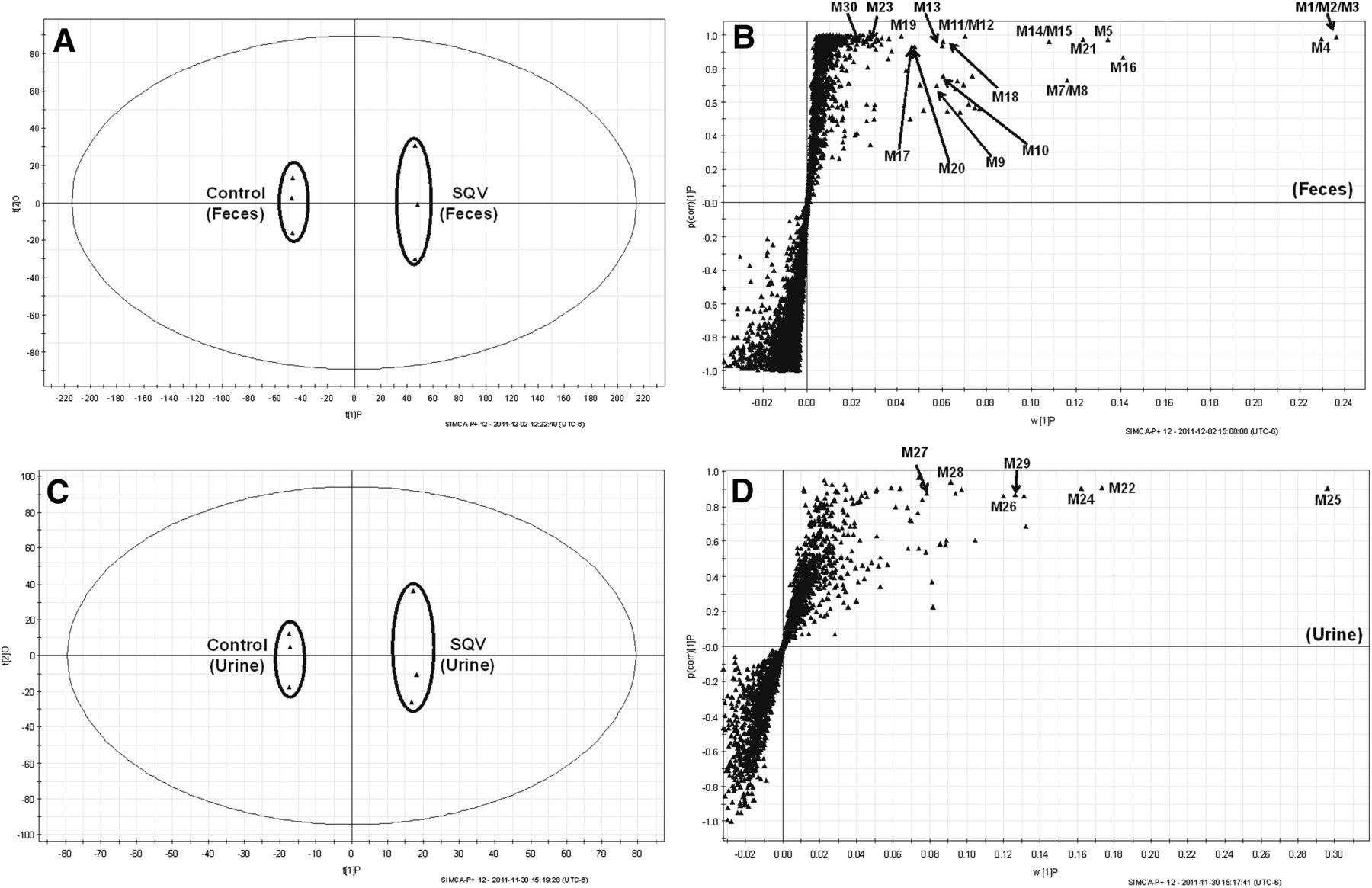
The results of the chemometric analysis on the ions produced by the UPLC-QTOFMS assay of control and SQV-treated mouse urine and feces are shown in Fig. 1. The supervised OPLS-DA analysis revealed two clusters (Fig. 1, A and C) corresponding to the control and SQV-treated groups in the score plots of the feces and urine, individually. The S-plots (Fig. 1, B and D) generated from OPLS-DA display an ion contribution to group separation in the feces and urine, respectively. The top-ranking ions were identified as SQV and its metabolites, which were marked in the S-plots (Fig. 1, B and D).

Metabolomic analysis of control and SQV-treated mouse urine and feces. Wild-type (WT) mice (n = 3) were treated with 150 mg/kg SQV (PO). Urine and feces were collected continuously over the 18 hours after treatment for analysis. (A, C) Separation of control and SQV-treated mouse feces and urine in OPLS-DA score plots, respectively. The t[1]P and t[2]O values represent the score of each sample in principal component 1 and 2, respectively. (B, D) Loading S-plots generated by OPLS-DA analysis of metabolome in SQV-treated mouse feces and urine. The x-axis is a measure of the relative abundance of ions, and the y-axis is a measure of the correlation of each ion to the model. These loading plots represent the relationship between variables (ions) in relation to the first and second components present in the OPLS-DA score plot. SQV and its metabolites were labeled in S-plots. The number of ions (metabolite identification) shown is in accord with Table 1.
SQV and its metabolites were found in both feces and urine, but mainly in feces (M1–M21, M23, and M30, Table 1). The metabolites M22 and M24–M29 with small molecular weights were excreted in the urine (Table 1). The information associated with SQV metabolites is summarized in Table 1.
Summary of SQV metabolites in mouse feces and urine.
The relative abundance of metabolites in mouse feces and urine is presented in Fig. 2. Overall, 30 SQV metabolites were identified in mice, including 10 known metabolites (M1–M3, M5–M9, M14, and M15) and 20 novel metabolites (Table 1). Twenty metabolites were also recapitulated in the incubation of SQV in HLM, and their relative abundance is shown in Supplemental Fig. 1. Among these novel SQV metabolites, one potential reactive α-hydroxyaldehyde (M22′) was trapped in HLM, and its further metabolite α-hydroxyacid (M23) was uncovered and verified in mouse feces.
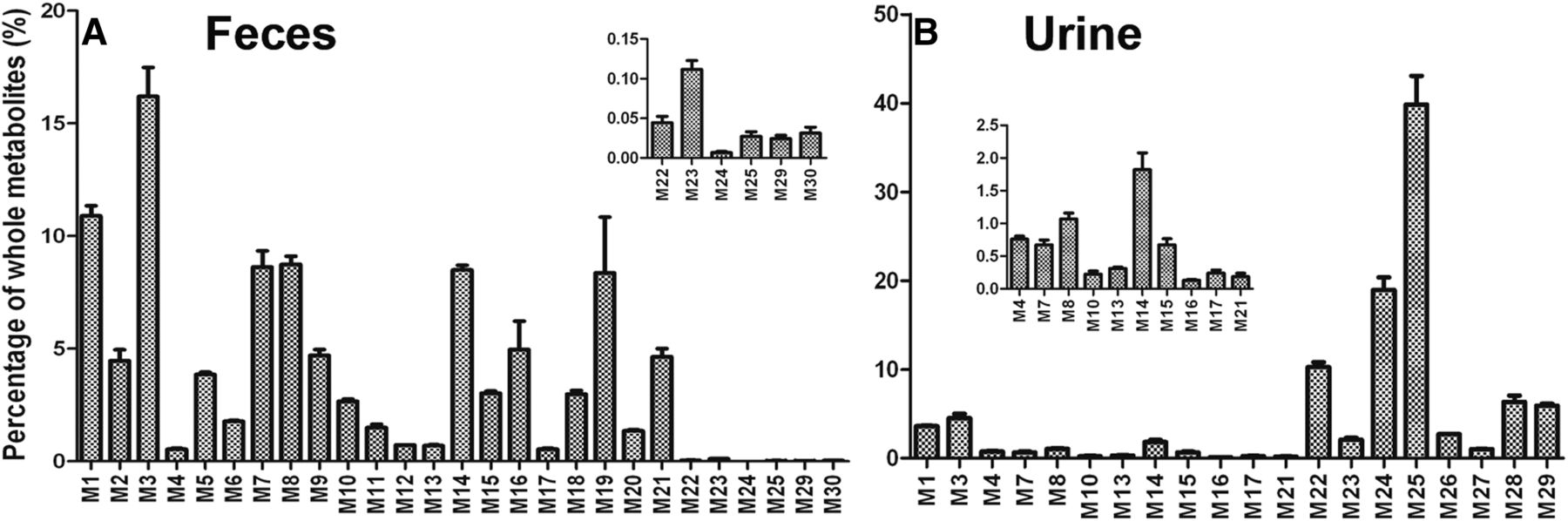
Relative abundance of metabolites in mouse feces and urine. Urine and feces from mice were collected continuously over the 18 hours after treatment for analysis (150 mg/kg, PO). The relative quantification was conducted based on the peak area. The overall abundance of metabolites was set as 100% in each sample. The data are expressed as mean (n = 3). (A) Relative abundance of metabolites in feces. (B) Relative abundance of metabolites in urine.
Identification of α-Hydroxyaldehyde (M22′) in SQV Metabolism.
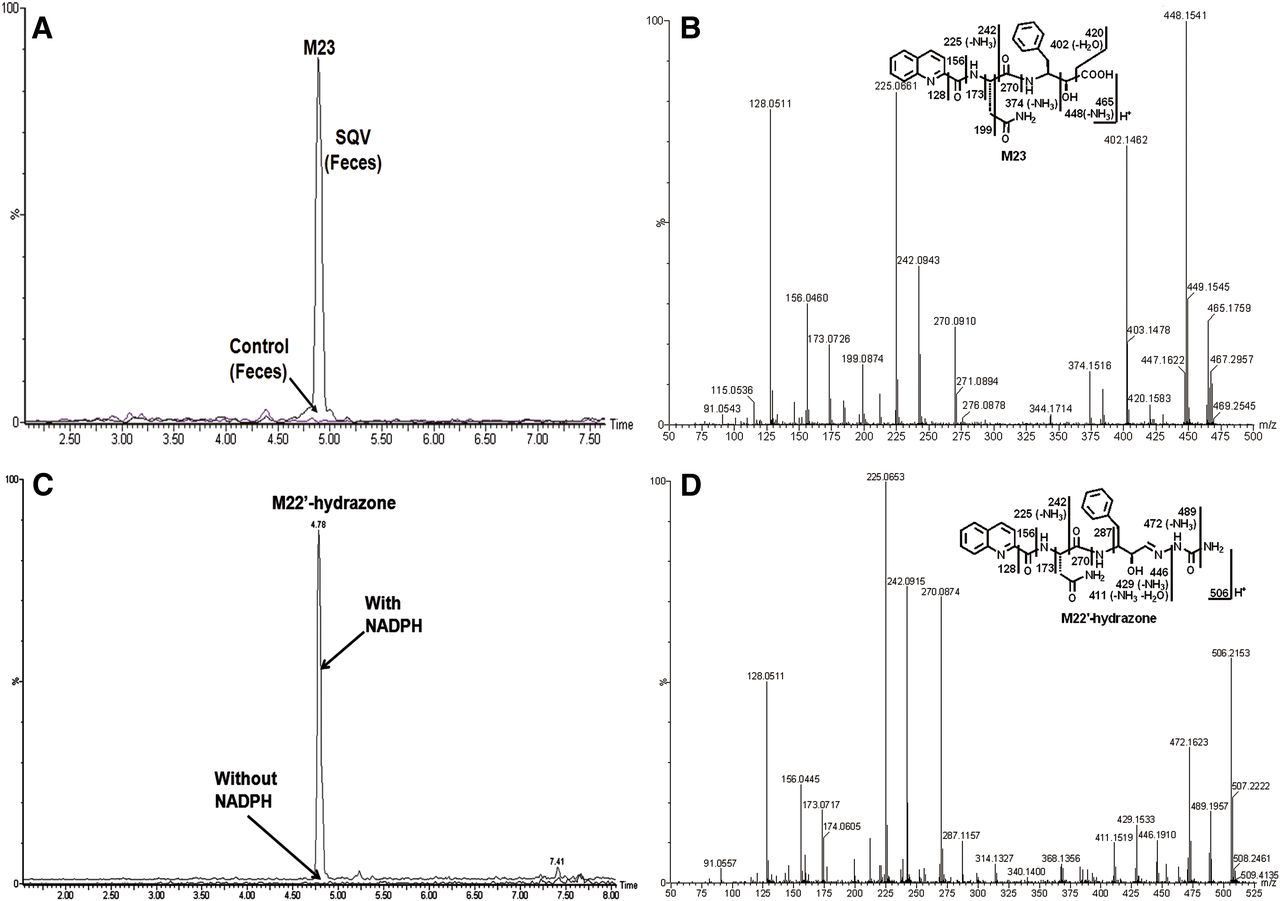
In SQV-treated mouse feces, a novel α-hydroxyacid (M23) was observed and characterized (Fig. 3, A and B). The formation of M23 indicated that its precursor, α-hydroxyaldehyde (M22′), was produced in the metabolism of SQV. Metabolite M23 was eluted at 4.86 minutes (Fig. 3A), having a protonated molecule [M+H]+ at m/z = 465 Da (Fig. 3B). The tandem mass spectrometry (MS/MS) analysis of M23 produced the major fragment ions at m/z 448 (loss of NH3), 402 (loss of COOH and H2O), 242, 225, and 128 (quinolinyl). The other fragment ions are interpreted in the inlaid structural diagram in Fig. 3B. The α-hydroxyaldehyde (M22′) was recapitulated and trappable in HLM using semicarbazide. The proposed mechanism of M22′ formation and the trapping strategy are shown in Fig. 4.

Identification of α-hydroxyaldehyde (M22′) in HLM and its further metabolite M23 in mouse feces. Feces from mice were collected continuously over the 18 hours after treatment for analysis (150 mg/kg, PO). In vitro studies were performed to trap M22′ using semicarbazide. Triplicate incubations were conducted in 1X PBS (pH 7.4) containing SQV (50 µM), HLM (1.0 mg/ml), semicarbazide (2.5 mM), with or without NADPH (1.0 mM). The metabolites were analyzed using UPLC-QTOFMS. Structural elucidations were performed based on accurate mass measurement (mass errors less than 10 ppm) and MS/MS fragmentations. MS/MS fragmentation was conducted with collision energy ramping from 10–40 V. The major daughter ions are interpreted in the inlaid structural diagrams. (A) The chromatograms of metabolite M23 in feces. (B) The MS/MS of M23. (C) The chromatogram of metabolite M22′-hydrazone in HLM. (D) The MS/MS of M22′-hydrazone.

A scheme of M22′ formation, trapping, and its further metabolism. M22′ was generated by dealkylation accompanied by M22, and it was trapped using semicarbazide in HLM. In mice, M22′ was further metabolized to acid M23.
Our studies demonstrate that the formation of M22′ is NADPH dependent (Fig. 3C). The formed M22′-hydrazone with semicarbazide, eluted at 4.78 minutes (Fig. 3C), has a protonated molecule [M+H]+ at m/z = 506 Da (Fig. 3D). The fragmental ions at m/z 489 and 446 suggest the formation of hydrazone. The major fragmental ions at m/z 472, 429, 270, 242, 225, 156, and 128 (quinolinyl) are interpreted in the inlaid structural diagram in Fig. 3D.
Identification of (3S)-N-(tert-Butyl)Decahydroisoquinoline-3-Carboxamide (M22).
Theoretically, the formation of α-hydroxyaldehyde M22′ should be accompanied by a dealkylated SQV metabolite (M22) (Fig. 4). As expected, M22 was observed in SQV-treated mouse urine (Table 1). The metabolite M22 was also detected in the SQV incubations with HLM. The chromatogram of M22 in SQV-treated mouse urine is presented in Fig. 5A. M22, eluted at 4.19 minutes, has a protonated molecule [M + H]+ at m/z = 239 Da. The fragment ions of M22, at m/z 183, 138, and 56, are interpreted in the inlaid diagram in Fig. 5B.

Identification of M22 and its further metabolites M24 and M25 in mouse urine. Urine and feces from mice were collected continuously over the 18 hours after treatment for analysis (150 mg/kg, PO). The metabolite elucidation conditions are described in Fig. 3. (A) Chromatograms of metabolites M22, M24, and M25. (B) MS/MS analysis of M22. (C) MS/MS analysis of M24. (D) MS/MS analysis of M25.
Identification of Monohydroxylated-M22 Metabolites (M24 and M25).
In mice, monohydroxylated M22 metabolites could be produced from two possible pathways: 1) M22 is further oxidized to M24 and M25; (2) dealkylation of monohydroxylated SQV generates M24 and M25 accompanied by α-hydroxyaldehyde M22′ (Fig. 6).

Proposed mechanism for the formation of M22′ and the accompanied metabolites M24–M28. M22′ can be generated from dealkylation of SQV or mono/dihydroxylated SQV. M22 and its further metabolites (M24–M28) were parallel to the formation of M22′. Alternatively, M24–M28 can be produced by further oxidation of M22, which originally generated from dealkylation of SQV.
M24, eluted at 2.87 minutes (Fig. 5A), has a protonated molecule [M + H]+ at m/z = 255 Da, 16 Da higher than that of M22. The fragment ions of M24, at m/z 199, 154, and 136, are interpreted in the inlaid diagram Fig. 5C. Compared with the MS/MS analysis of M22, the ion at m/z 154 is 16 Da higher than the ion at 138 generated from M22, which suggests that the oxidation occurred in the encircled unit (Fig. 5C).
M25 was eluted at 3.55 minutes (Fig. 5A), having a protonated molecule [M + H]+ at m/z = 255 Da. The fragment ions of M25 at m/z 237 (loss of H2O), 199, 154, and 136 are interpreted in the inlaid diagram in Fig. 5D. M25 has similar fragment patterns to those of M24. Therefore, the oxidation happened in the same encircled unit but at a different site.
In addition, three dihydroxylated-M22 metabolites—M26, M27, and M28—were detected in SQV-treated mouse urine. Their possible mechanism of formation, chromatograms, and identification are presented in Fig. 6 and Supplemental Fig. 2.
Role of CYP3A in the Formation of α-Hydroxyaldehyde (M22′).
Aldehyde is not easily detected by mass spectrometry (MS). The formation of the aldehyde M22′ is proportional to the generation of M22 and its further metabolites, which can be readily detected by MS. Accordingly, M22 was employed to evaluate the role of P450 in the formation of the α-hydroxyadehyde M22′ in vitro.
The incubation of SQV with different human cDNA-expressed P450s (control, CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4) revealed that CYP3A4 is the primary enzyme contributing to the formation of M22 (Supplemental Fig. 3A). The role of CYP3A4 in the formation of M22 was further verified by coincubation of RTV, a potent CYP3A4 inhibitor, in HLM. The formation of M22 was suppressed up to 85% by RTV at 2 µM (Supplemental Fig. 3B).
To further confirm the role of CYP3A in the formation of α-hydroxyaldehyde (M22′), Cyp3a-null mice were used. The production of α-hydroxyaldehyde (M22′) is positively related to the formation of these metabolites (M22–M25), which were used to evaluate the contribution of CYP3A in the formation of M22′. The relative abundances of M22, M23, M24, and M25 in the urine of Cyp3a-null mice were 16%, 7%, 8%, and 40%, respectively, of these metabolites in WT mice (Supplemental Fig. 4). These data suggest that CYP3A plays an important role in the formation of M22′ both in vitro and in vivo.
Discussion
In this study, a liquid chromatography–mass spectrometry (LC-MS)-based metabolomic approach was employed to profile SQV metabolism in mice. We identified 30 SQV metabolites, including 10 previously reported metabolites (M1–M3, M5–M9, M14, and M15) (Fitzsimmons and Collins, 1997; Eagling et al., 2002) and 20 novel metabolites. SQV and its metabolites were primarily excreted in feces and small portions in urine (Table 1), which is in agreement with previous studies. In hepatic and intestinal microsomes, SQV was mainly metabolized to mono/dihydroxylated metabolites. In our study, novel trihydroxylated (M10), dehydrogenated (M11 and M12), di/trihydroxylated plus dehydrogenated (M4 and M17–M21), and tridehydrogenated (M13) metabolites were detected and identified in mouse feces (Table 1). Amide hydrolysis produced two carboxylic acids (M16 and M29) and one detectable primary amine (M30), which were identified for the first time. Interestingly, the observation of α-hydroxyacid (M23) in feces and the corresponding M22 in urine indicated the generation of α-hydroxyaldehyde (M22′) in the metabolism of SQV.
M22′, α-hydroxyaldehyde, is a potential reactive metabolite from the metabolism of SQV in mice. The further metabolite M23, α-hydroxyacid, was detected instead of M22′ in mouse feces. In HLM, the existence of α-hydroxyaldehyde was confirmed by the formation of hydrazone with semicarbazide (Figs. 3 and 4). Previous studies have suggested that α-hydroxyaldehyde can form a Schiff base with free amino groups on the protein. The unstable α-hydroxyimine can be transformed into a stable 1-amino-2-keto protein adduct by intramolecular Amadori rearrangement, which then results in toxicity (Spahn-Langguth and Benet, 1992; Tang and Lu, 2010).
Acyl glucuronides of drugs have been regarded as reactive metabolites because of the final production of α-hydroxyaldehyde (Bailey and Dickinson, 2003). The α-hydroxyaldehyde was generated by the scission of the hemiacetal ether bond, which was formed through acyl migration from 1- to the 2-, 3-, or 4-position at the glucuronic acid moiety. For instance, benoxaprofen, zomepirac, indoprofen, and alclofenac (nonsteroidal anti-inflammatory drugs) were withdrawn from market due to unacceptably high incidence of adverse drug reactions (Wolf et al., 1984). All these drugs are metabolized in humans to acyl glucuronides, but the exact mechanisms are still unclear.
We have demonstrated that CYP3A4 is the dominant enzyme responsible for the formation of α-hydroxyaldehyde M22′ both in vitro (Supplemental Fig. 3) and in vivo (Supplemental Fig. 4). To this end, cotreating SQV with the inducers of CYP3A4 will increase the formation of reactive metabolite M22′, which may augment the toxicity of SQV. Rifampicin (RIF), a first-line antituberculosis (anti-TB) drug, is a potent CYP3A4 inducer in vivo (Backman et al., 1998). For HIV/TB coinfected patients, the World Health Organization requires first treating patients with anti-TB drugs for 2 weeks before initiating anti-HIV treatment. Pharmacokinetic studies in patients with HIV/TB coinfection suggest that RIF can significantly decrease the area under a curve from 0 to 24 hours of SQV and RTV by 39.5% and 42.5%, respectively (Ribera et al., 2007). In a recent comparative clinical study, all healthy volunteers who were pretreated with RIF followed by treatment with SQV/r experienced more severe elevation of hepatic transaminases than those who received SQV/r alone (Schmitt et al., 2009). In our previous study, RTV was also bioactived to α-ketoaldehydes via CYP3A-mediated thiazole ring opening (Li et al., 2011a). Consequently, the possible mechanism of the toxicity related to SQV/r is that RIF induces CYP3A expression, which increases the formation of reactive metabolites of SQV/r and then leads to severe liver injury. Alternatively, SQV/r may exacerbate the hepatotoxic potential of RIF by inhibiting the normal metabolic breakdown of RIF, because long-term treatment with RIF may cause liver toxicity as well (Corpechot et al., 2006). Taken together, our data and clinical findings suggested that SQV/r should be given with caution in patients receiving CYP3A4 inducers.
The primary amine M30 from amide hydrolysis may be also related to SQV toxicity. Previous studies have suggested that primary amine can be further oxidized to nitroso compounds, which can irreversibly inhibit enzymes, leading to drug-drug interaction or toxicity (Tang et al., 2008). However, M30 was not observed in HLM, so more work is needed to determine its formation in humans and the role of M30 in SQV toxicity.
In summary, we extensively investigated the metabolism of SQV in mice and in HLM by the use of LC-MS-based metabolomics. Our study tripled the number of known metabolites of SQV (Fig. 7) and for the first time identified two potentially toxic metabolites, including α-hydroxyaldehyde and one primary amine. Additionally, we determined CYP3A4 as the primary enzyme contributing to the formation of α-hydroxyaldehyde in vitro and in vivo. Further studies are needed to investigate the importance of these potential reactive metabolites in SQV-related adverse effects.

Summary of putative structures of SQV metabolites in mice. All structures were determined based on the exact mass (mass error less than 10 ppm) and MS/MS fragments. MS/MS fragmentation was conducted with collision energy ramping from 10–40 V.
Acknowledgments
The authors thank the National Institutes of Health AIDS Research and Reference Reagent Program for providing the HIV protease inhibitors, Dr. Curtis D. Klaassen for providing the Cyp3a-null mice, and Dr. Hartmut Jaeschke for scientific suggestions.
Authorship Contributions
Participated in research design: Li, Ma.
Conducted experiments: Li, Lu.
Contributed the new reagents or analytic tools: Li.
Performed data analysis: Li, Ma.
Wrote or contributed to the writing of manuscript: Li.
Footnotes
- Received September 17, 2013.
- Accepted November 8, 2013.
This work was supported by the National Institutes of Health National Center for Research Resources [Grant COBRE 5P20-RR021940]; the National Institute of General Medical Sciences [Grant 8P20-GM103549-7]; and the National Institute of Allergy and Infectious Diseases [Grant AI095425].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- CYP450
- cytochrome P450
- HIV
- human immunodeficiency virus
- HLM
- human liver microsomes
- LC
- liquid chromatography
- MS
- mass spectrometry
- MS/MS
- tandem mass spectrometry
- OPLS-DA
- orthogonal projection to latent structures-discriminant analysis
- PBS
- phosphate-buffered saline
- PO
- by mouth
- QTOFMS
- quadrupole time-of-flight mass spectrometry
- RIF
- rifampicin
- RTV
- ritonavir
- SQV
- saquinavir
- SQV/r
- saquinavir and ritonavir
- TB
- tuberculosis
- UPLC
- ultraperformance liquid chromatography
- WT
- wild type
- Copyright © 2013 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}