


Abstract
Freshly isolated hepatocytes are considered the gold standard for in vitro studies of hepatic drug disposition. To ensure a reliable supply of cells, cryopreserved human hepatocytes are often used. ABC-superfamily drug efflux transporters are key elements in hepatic drug disposition. These transporters are often considered lost after isolation of hepatocytes. In the present study, the expression and activity of ABC transporters BCRP, BSEP, P-gp, MRP2, MRP3, and MRP4 in human and rat cryopreserved hepatocytes were investigated. In commercially available human cryopreserved hepatocytes, all drug efflux transporters except human BCRP (hBCRP) exhibited similar expression levels as in fresh liver biopsies. Expression levels of hBCRP were 60% lower in cryopreserved human hepatocytes than in liver tissue, which could lead to, at most, a 2.5-fold reduction in hBCRP-mediated efflux. Fresh rat hepatocytes showed significantly lower levels of rat BCRP compared with liver expression levels; expression levels of other ABC transporters were unchanged. ABC transporters in human cryopreserved cells were localized to the plasma membrane. Functional studies could demonstrate P-gp and BCRP activity in both human cryopreserved and fresh rat hepatocytes. Inhibiting P-gp–mediated efflux by elacridar in in vitro experiments significantly decreased fexofenadine efflux from hepatocytes, resulting in an increase in apparent fexofenadine uptake. The results from the present study clearly indicate that ABC transporter-mediated efflux in freshly isolated as well as cryopreserved rat and human hepatocytes should be taken into account in in vitro experiments used for modeling of drug metabolism and disposition.
Introduction
Isolated hepatocytes are used to determine many properties of a candidate drug, its intrinsic clearance (CLint), and metabolic pathways, and increasingly also to assess drug-drug interactions mediated by drug-metabolizing enzymes and/or drug transporters (Soars et al., 2007a, 2009; Zhao, 2008; Chiba et al., 2009; McGinnity et al., 2009; Brown et al., 2010; Giacomini et al., 2010).
Cryopreserved human hepatocytes are used as a convenient and reliable source of cells in drug discovery research, whereas for preclinical species freshly isolated hepatocytes are commonly available (Chiba et al., 2009; Giacomini et al., 2010). Characterization of both freshly isolated and cryopreserved hepatocytes from a number of species has been reported, focusing on drug-metabolizing enzymes (Chiba et al., 2009; Soars et al., 2009). The studies have concluded that the drug-metabolizing enzymes are retained on the same level in cryopreserved hepatocytes as in freshly isolated cells (Floby et al., 2009).
Several ABC efflux transporters such as P-glycoprotein (P-gp), multidrug-resistance protein 2 (MRP2), breast cancer–resistance protein (BCRP), and the bile salt efflux pump (BSEP) are important for physiological functions of the liver but are also exporting organic compounds, including drugs, from the hepatocytes. These are all localized to the bile-facing canalicular apical membrane of the hepatocytes and are important for biliary secretion of bile salts and drugs (Klaassen and Aleksunes, 2010). Additionally, some ABC transporters such as MRP3 and MRP4 are expressed on the basolateral membrane and export compounds into the bloodstream (Klaassen and Aleksunes, 2010). MRP3 and MRP4 have been indicated to play a central role in basolateral efflux of sulfate- and glucoronide-conjugated drug metabolites (Zamek-Gliszczynski et al., 2006) and have been shown to be involved in basolateral efflux of fexofenadine and rosuvastatin (Tian et al., 2008a; Pfeifer et al., 2013).
Fexofenadine, rosuvastatin, and digoxin are examples of drug compounds being eliminated, in whole or in part by bile secretion in humans through efflux via ABC transporters (Cvetkovic et al., 1999; Roberts et al., 2002; Kitamura et al., 2008; Yang et al., 2009). Erythromycin is an example of a drug that is cleared in the human liver by the concerted action of the ABC transporter P-gp (human P-gp, hP-gp) and cytochrome P450 3A4 (CYP3A4) (Kurnik et al., 2006). Fexofenadine and erythromycin are selective substrates of P-gp, whereas rosuvastatin is predominantly effluxed via BCRP (Morrissey et al., 2012). These compounds are thus valuable probes for the study of efflux transporter activity and of transporter-enzyme interplay and were extensively used in the present study together with selective transporter inhibitors.
The fate of ABC efflux transporters in isolated and cryopreserved hepatocytes has been debated. Some reports suggest that cryopreserved hepatocytes have lost most of their ABC transporter function as a result of a rapid and partial transporter internalization after isolation of the hepatocytes (Oude Elferink et al., 1993; Bow et al., 2008). No quantification of the extent of transporter loss was attempted.
Other results have pointed to the presence and activity of several major efflux transporters in freshly isolated hepatocytes as well as cryopreserved hepatocytes from both rats and humans (Lam and Benet, 2004; Lam et al.,, 2006; Li et al., 2008, 2009a, 2009b). In the study by Di et al. (2012), significant P-gp activity in cryopreserved hepatocytes was inferred from in vitro–in vivo extrapolation (IVIVE) of hepatic clearance (CLhepatic) for P-gp substrates from human liver microsomes and cryopreserved hepatocytes. However, in IVIVE of hepatic drug clearance using freshly isolated or cryopreserved hepatocytes, the activity of ABC transporters is most often considered lost after hepatocyte isolation (Zamek-Gliszczynski et al., 2013).
The aim of the present study was to investigate the applicability of cryopreserved hepatocytes for drug disposition predictions. We specifically focused on efflux transporters and investigated the function and localization of these in human and rat cryopreserved hepatocytes. The abundance of efflux transporters was also investigated in isolated hepatocytes and compared with levels in liver tissue. The impact of ABC transporter-mediated efflux on in vitro uptake and metabolism measurements was assessed. Apart from BCRP, no tested ABC efflux transporter (BSEP, P-gp, MRP2, MRP3, and MRP4) showed significant downregulation in human cryopreserved or rat freshly isolated hepatocytes. The efflux transporters exhibited activity that significantly affected in vitro measurements of drug disposition.
Materials and Methods
Nomenclature.
As recommended by Hagenbuch and Stieger (2013), this article follows the gene and protein nomenclature guidelines of the HUGO Gene Nomenclature Committee (Gray et al., 2013) and the International Committee for Standardized Genetic Nomenclature in Mice (http://www.informatics.jax.org/mgihome/nomen/2009_gene.shtml) (2009). Human genes are written in capital letters (e.g., ABCG2), whereas rodent genes are given in upper and lower case (e.g., Abcg2). Protein names for both human and rodent proteins are written in capital letters (e.g., BCRP). In the interest of clarity, human protein names are prefixed with “h,” whereas rat proteins are prefixed “r” (e.g., hBCRP, rBCRP). Protein names without prefix refer to the protein in general, not belonging to any particular species (e.g., a BCRP inhibitor).
Chemicals.
Chemicals were purchased from Sigma-Aldrich (St. Louis, MO), unless otherwise specified.
Animals.
Male Sprague-Dawley rats were acquired from Harlan Laboratories B.V. (Venray, The Netherlands). Rats were housed up to three per cage in transparent plastic cages. All animals were acclimatized for 2 weeks prior to studies. The room was lit in a 12-hour light-dark cycle. Bedding material consisted of aspen wood chips, and plastic tunnels and aspen chewing blocks were provided. The rats had free access to food and water. All animal experiments were approved by the local animal ethics committee (Stockholms Södra djurförsöksetiska nämnd) and were conducted in compliance with national guidelines for the care and use of laboratory animals.
Preparation of Hepatocytes.
Rat hepatocytes were isolated according to a modified collagenase perfusion method previously described (Bissell and Guzelian, 1980) from 7- to 8-week-old male Sprague-Dawley rats anesthetized with Isoflurane (Abbott Scandinavia AB, Kista, Sweden). Shortly, the liver was perfused in situ using a peristaltic pump, first with 37°C calcium-free buffer containing 0.5 M EGTA, followed by prewarmed William’s E medium, supplemented with 10 mM HEPES (Gibco; Life Technologies, Paisley, U.K.), pH 7.4 (WE). To the buffer was added 0.16 mg/ml collagenase type XI. After completed perfusion, the organ was cut from its ligaments and transferred to a prewarmed collagenase solution. Released from connective tissue, the hepatocytes were washed to obtain a homogenized cell suspension without debris from extracellular matrix. The suspension was filtered through gauze, repeatedly centrifuged at 50 × g for 2 minutes, and resuspended in new Krebs-Henseleit (KHL) buffer supplemented with 10 mM HEPES (Gibco), pH 7.4, until a homogenous suspension of cells was obtained. Cell number and viability were determined using a Bürker counting chamber and the Trypan blue exclusion method. Viability typically exceeded 90%. Cell preparations with viability of less than 80% were discarded.
Handling of Human and Rat Cryopreserved Hepatocytes.
Human: lots Hu4035, Hu4037, and Hu4152 (CellzDirect; Life Technologies) and lots OZL, SQJ, and REL (Celsis In Vitro Technologies, Baltimore, MD); donor information is listed in Supplemental Table 1. The lots represented a mix of ages and genders and displayed average P450 metabolism. Rat: derived from male Sprague-Dawley rats (CellzDirect), 7–9 weeks of age; only two lots were available. Cryopreserved hepatocytes were thawed in a 37°C water bath immediately after removal from the −150°C freezer and transferred to a 50 ml vial of WE at 37°C. Centrifugation was done according to the manufacturer’s instructions, and the cell pellet was resuspended in a small volume of KHL. Cell number and viability were determined as described for rat hepatocytes.
For direct efflux measurements, hepatocytes were plated in collagen I-coated 12-well plates (BD Biocoat; BD Biosciences, Franklin Lakes, NJ). In each well was seeded 0.75 million cells, from a suspension of 106 cells/ml in WE supplemented with 10% fetal calf serum (FCS), 2 mM glutamine, and 0.01% insulin-transferrin-selenium A (Gibco). Prior to 10-minute incubation at room temperature (RT), the plate was shaken carefully to let the cells distribute evenly. Cells were then left to attach to the plates for 90 minutes at 37°C in an incubator containing 5% CO2. The plates were washed three times in 37°C KHL. The distribution of adhered cells was inspected using an inverted microscope after washing. Cells were then preincubated in KHL for 10 minutes before each experiment.
Preparation of Hepatocyte Protein.
From liver tissue, rat liver was handled in WE in a petri dish, cut into smaller pieces, and homogenized with a glass pestle homogenizer containing a cracking buffer, consisting of 5 mM HEPES, 0.5 mM EDTA, and 1% protease inhibitor cocktail (Sigma-Aldrich), pH 7.2. The resulting suspension was transferred to a tube and homogenized thoroughly with a small gauge syringe before centrifugation to remove unhomogenized tissue. Small samples of human liver tissue were taken from an in-house tissue biobank and processed in the same manner (n = 3); donor information is given in Supplemental Table 1. The liver biobank derives from the Karolinska University Hospital (Stockholm, Sweden). Samples were collected from liver resections with approval from the Karolinska University Hospital ethical review board.
From hepatocytes, aliquots of 1 million fresh, snap frozen after isolation, rat hepatocytes collected with the preparation technique described above, were thawed and suspended in 200 μl cracking buffer. Homogenization was performed as described for liver tissue.
From cryopreserved hepatocytes, rat and human cryopreserved hepatocytes (CellzDirect; Celsis In Vitro Technologies), stored at −150°C, were thawed and pelleted by centrifugation at 90 rpm for 3 minutes and resuspended in cracking buffer. Homogenization was performed as described for liver tissue.
Determination of protein concentration was performed with a bicinchoninic acid protein assay kit (Pierce Biotechnology, Thermo Fisher Scientific, Rockford, IL), according to the manufacturer’s instructions. Samples were stored at −80°C until use.
Determination of Transporter Expression by Western Blot.
A total of 10–40 μg protein, prepared as described above, was diluted in dH2O and loading buffer [0.5 ml Laemlli buffer, 50 µl β-mercaptoethanol, and 2.5 µl 1 M dithiothreitol, all from Bio-Rad (Hercules, CA)] and separated by 9% SDS-PAGE, followed by transfer of protein to a polyvinylidene difluoride membrane (Perkin-Elmer, Waltham, MA) at 200 mA for 2 hours. A broad range unstained molecular weight ladder (Bio-Rad) was used as a reference and detected after Ponceau S staining. Ponceau S staining was also scanned and quantified to ensure that equal amounts of protein had been added to each lane.
The membranes were washed 2 × 5 minutes in Tris-buffered saline Tween 20 [0.1% Tween 20 in a buffer consisting of 150 mM NaCl and 25 mM Tris-HCl, pH 7.4 (TBST)] before continuing with blocking for 1 hour in blocking buffer (consisting of 5% nonfat dry milk in TBST with pH adjusted to 7.2 with 1 M Tris base) at room temperature.
Membranes were incubated with primary antibody (primary antibodies used are listed in Supplemental Table 2), diluted 1:500 or 1:1000, at 4°C overnight, followed by washing in TBST for 4 × 15 minutes. Blocking was again performed during 1 hour at room temperature, followed by incubation with a 1:1000 or polyclonal goat anti-mouse IgG conjugated to HRP (Dako, Glostrup, Denmark), depending on the species of the primary antibody. To detect glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a HRP-conjugated primary antibody was used, omitting the need for a secondary antibody (Supplemental Table 2).
Membranes were washed again, first in TBST and thereafter for 2 × 5 minutes in dH2O. Antibodies were chemiluminescently detected using Western blotting ECL Plus reagents (GE Healthcare, Piscataway, NJ), a substrate for HRP, according to the manufacturer’s instruction. The blot was photographed with Amersham hyperfilm ECL (GE Healthcare), developed, and fixed. Protein bands were scanned, and the background was subtracted. To normalize band intensities, the ratio between the transporter band and the band from the housekeeping protein, GAPDH, was calculated.
Immunofluorescence.
Human cryopreserved hepatocytes were allowed to attach to slides (Super Frost; Thermo Fisher Scientific) for 1 hour at room temperature. After washing, adherent cells were fixed in ice-cold 50% acetone for 2 minutes, followed by a 5-minute fixation in 100% acetone. Coverslips were then dried and stored at −20°C until use.
Coverslips were washed 4 × 5 minutes in PBS and blocked with 10% FCS (Gibco) in PBS for 30 minutes at room temperature. They were then washed again and incubated with primary antibody in 1% FCS in PBS for 1 hour at room temperature. Primary antibodies were used at a dilution of 1:100. Cells were washed again and blocked with 10% goat serum (Life Technologies) in PBS for 30 minutes at room temperature. Slides were then incubated with a 1:1000 dilution of goat anti-mouse Alexafluor-conjugated fluorescent secondary antibodies (Life Technologies) in PBS with 10% goat serum and double stained with 4′,6′-diamidino-2-phenylindole (Life Technologies). Coverslips were washed one final time for 4 × 5 minutes in PBS and mounted with Prolong Gold anti-fade mounting media (Life Technologies). Stained cells were photographed using a confocal microscope (DMIRE 2; Leica Microsystems, Wetzlar, Germany).
Liver tissue samples were obtained from donors undergoing surgical liver resection at Uppsala University Hospital (Uppsala, Sweden), as approved by Uppsala Regional Ethical Review Board. Small biopsies from liver resections were snap frozen in ethane cooled by liquid nitrogen in the operating theater immediately after the surgical procedure (n = 3); donor information is given in Supplemental Table 1. Frozen biopsies were stored at −80°C until use. Liver biopsies were cut into 8-μm–thick sections on a cryotome and dried down on slides (Super Frost; Thermo Fisher Scientific). Liver sections were fixed, stained, and visualized, as described for hepatocyte samples above.
Uptake, Efflux, and Metabolic Stability Measurements in Hepatocytes.
To directly measure efflux, plated hepatocytes in 12-well plates were used. To start uptake, KHL was removed and 300 μl 1 µM rosuvastatin diluted in KHL, containing 0.1% dimethyl-sulfoxide, was added to each well. After 30 minutes of preloading, the cells were rapidly washed three times with 37°C KHL, after which 300 μl fresh KHL was added to each well. Efflux from the cells was followed by sampling 200 μl buffer from each well at different time points, 0.5, 1, 2, and 5 minutes. Efflux was quantified at the 1-minute time point, well within the linear range of efflux. Samples were mixed with 2 volumes of an ice-cold stop solution of acetonitrile with 200 nM warfarin (internal standard). Plates containing samples were incubated on ice for at least 10 minutes to ensure complete precipitation of protein. The plate contents were shaken for a few minutes prior to centrifugation at 1200 × g for 10 minutes. An aliquot of 200 μl supernatant from each well was transferred to a 96-well analysis plate. Before analysis by liquid chromatography tandem mass spectrometry (LC-MS/MS), each fraction was diluted in KHL to 25% acetonitrile final concentration.
Uptake measurements in suspended cells were performed using the media loss technique, as described (Soars et al., 2007b). Briefly, hepatocytes (106 cells/ml) were incubated with test compound at a concentration of 1 μM in KHL with the addition of 0.1% dimethyl-sulfoxide. At intervals ranging from 0 and 15 seconds up to 90 minutes, aliquots were transferred to a glass tube loaded into a bench top centrifuge and hepatocytes were pelleted by centrifugation (7000 × g, 15 seconds), after which a sample was taken from the supernatant. Samples were processed and analyzed, as described above.
To assess metabolic elimination of compound, an aliquot of cells and medium was removed from the incubation, lysed, and processed, as above, omitting the centrifugation step.
Uptake of fexofenadine and uptake and metabolism of erythromycin were assessed using the spinning-through-oil method (Petzinger and Fückel, 1992). An advantage with this method is that compound concentrations in cells and medium can be measured separately. The incubation of hepatocytes and test compound was layered on top of a layer of mineral oil (d = 1.02 g/ml; Sigma-Aldrich). Below the oil meniscus, a lower phase consisting of 50 μl 4 M CsCl was found. At the designated time points, the uptake reaction was stopped by spinning the cells through the oil layer (1200 × g, 30 seconds). A sample was taken of the supernatant, after which it, the oil meniscus, and the lower CsCl phase were removed by careful pipetting to not disturb the cell pellet. The cells were then resuspended in 50 μl KHL, lysed, and processed, as described above. Compound concentrations were determined by LC-MS/MS.
The effects of efflux through ABC transporters on drug efflux, drug metabolism, compound uptake, and metabolite efflux were investigated using the P-gp and BCRP inhibitor elacridar (also known as GF120918) or the BCRP inhibitor fumitremorgin C (FTC) in rat fresh and human cryopreserved hepatocytes using terfenadine, fexofenadine, erythromycin, or rosuvastatin as probe substrates (Supplemental Table 3). At the concentrations of 0.5–2 µM used in the experiments, elacridar inhibition of BCRP is minor, less than 20% (Matsson et al., 2009). Fexofenadine and erythromycin were used in P-gp inhibition experiments using the inhibitor elacridar, as these two compounds do not seem to interact with BCRP (Ozawa et al., 2004; Morrissey et al., 2012). Rosuvastatin and fexofenadine efflux from plated hepatocytes was measured in the presence or absence of inhibitors. After addition of terfenadine to hepatocytes, P-gp activity was detected monitoring efflux of the terfenadine metabolite fexofenadine, in the presence or absence of elacridar. The effects on fexofenadine uptake by inhibiting P-gp–mediated efflux with elacridar were then investigated using media-loss and spinning through oil experiments. Spinning through oil experiments with or without elacridar were performed to monitor uptake, metabolism, and efflux of erythromycin and its metabolites. In each reaction, the concentration of the inhibitor was optimized to give maximal effects on the reaction studied without affecting other parameters such as uptake or P450 metabolism of the test compound.
Detection of Test Compounds.
Sample analysis was performed by LC-MS/MS using a Micromass Quattro Micro triple quadrupole (Micromass, Manchester, U.K.) coupled to a gradient pump composed of two Shimadzu LC-10AD VP isocratic pumps (Shimadzu, Kyoto, Japan) and a CTC HTS PAL autosampler (CTC Analytics, Zwingen, Switzerland). The software MassLynx (controlling the LC system and mass spectrometer), which includes QuanLynx (quantification) and QuanOptimize (MS/MS optimization), was used (Waters, Milford, MS). A high purity C18 5 μm 30 × 2.1-mm (Thermo Electron, Waltham, MA) analytical column was used. Chromatography was performed using a generic gradient at a flow rate of 0.4 ml/min. The mobile phase consisted of solvent 2% acetonitrile in 0.1% (v/v) acetic acid in water and 80% acetonitrile in 0.1% (v/v) acetic acid in water. The injection volume was 20 µl, and warfarin was used as internal standard. Other source parameters (e.g., collision energy, cone voltage, ion mode, molecular weight of parent and daughter, etc.) were individually optimized for each compound (see Supplemental Table 4). Responses were detected as the area under the curve, and the relative concentration was determined with the internal standard. Absolute concentrations were calculated from standards spanning from twice the initial experimental concentration to below the limit of quantification. Limit of quantification, signal-to-noise ratio for detection, was set to 5 times. Terfenadine metabolism was monitored by quantification of fexofenadine and erythromycin metabolism by the formation of N-desmethyl-erythromycin.
Determination of Intrinsic Clearance.
The elimination rate constant was calculated from the first-order elimination equation
 (1)
(1)where [S] is concentration of substrate at a given time point, t, after start of the incubation at the initial concentration [S0]. k is the elimination rate constant and was determined by nonlinear regression within a time point interval considered to represent the initial elimination rate either in metabolism or uptake experiments (Soars et al., 2007b). Intrinsic clearance for metabolism or uptake (CLint, met or CLint, uptake) could then be calculated from:
 (2)
(2)where V represents the volume of incubated substrate.
IVIVE of in vitro CLint, uptake values was performed for fexofenadine uptake experiments in the absence or presence of increasing concentrations of elacridar. IVIVE was performed using the well-stirred model including fraction unbound in both in vitro and in vivo systems, as described by Sohlenius-Sternbeck et al. (2012), omitting the regression line correction step employed in that study. The physiologic scaling factors used are listed in Sohlenius-Sternbeck et al. (2012). In vivo blood CL for fexofenadine was determined in house in Sprague-Dawley rats after intravenous administration of a dose of 3 µmol/kg (Lundquist et al., 2014). CLhepatic for fexofenadine was calculated from the blood CL using data on the hepatic fraction of clearance from the literature (Swift et al., 2009). Human pharmacokinetic parameters for erythromycin are from Sun et al. (2010).
Statistical Analysis.
Data were analyzed using Graphpad Prism 5 software (Graphpad Software, La Jolla, CA). Statistical differences were tested using the two-tailed Student’s t test. Probability values (P values) are symbolized by *P < 0.05, **P < 0.01, and ***P < 0.001. P values <0.05 were considered statistically significant.
Results
ABC Efflux Transporter Expression in Human and Rat Hepatocytes.
Western blotting was used to determine expression levels of drug efflux transporters from the ABC superfamily, hBCRP, hBSEP, hMRP2, hMRP3, hMRP4, and hP-gp in human cryopreserved hepatocytes and human liver samples. In total, six different batches of cryopreserved hepatocytes (three obtained from CellzDirect and three from Celsis In Vitro Technologies) and samples from three liver donors were tested. Both cryopreserved hepatocytes and liver samples derived from donors of mixed gender (Supplemental Table 1). No differences could be seen between cryopreserved cells obtained from the different providers. No statistically significant differences of efflux transporter protein levels in cryopreserved human hepatocytes and liver tissue were seen, with the exception of hBCRP, which was approximately 60% lower in cryopreserved hepatocytes compared with levels in fresh liver (Fig. 1).
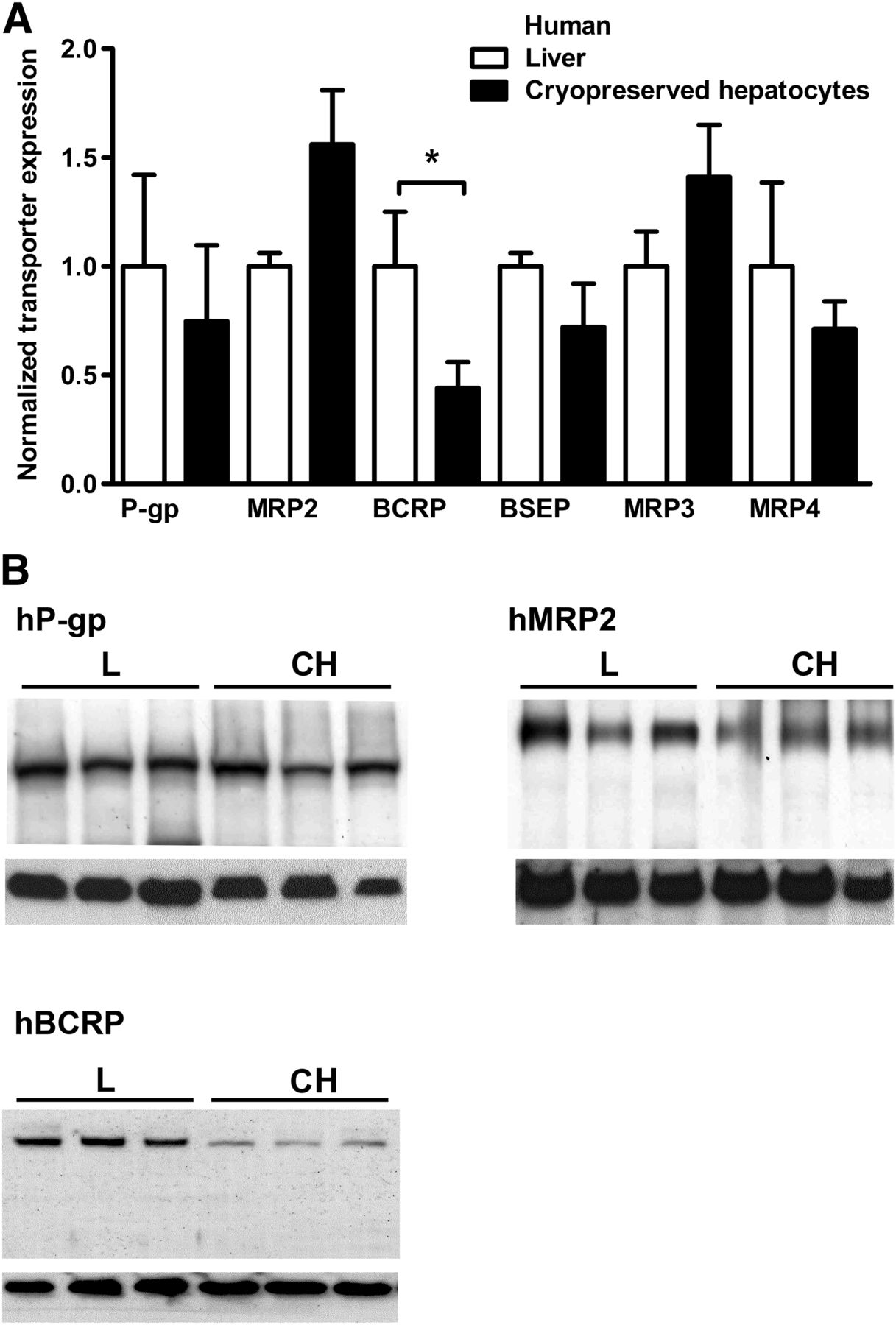
(A) Quantification of ABC transporter expression in human liver and cryopreserved hepatocytes. Transporter protein levels were normalized against GAPDH; expression levels in liver tissue were set to 1. Of the tested transporters, only hBCRP was downregulated (by 60%) after cryopreservation of hepatocytes (mean ± SD, n = 3; *P < 0.05). (B) Representative Western blots of hP-gp, hMRP2, and hBCRP derived from human liver and cryopreserved hepatocytes are shown. Lower panels show the internal standard hGAPDH. Approximate molecular weights were hBCRP, 140 kDa (dimer); hMRP2, 190 kDa; hP-gp, 170 kDa; and hGAPDH, 35 kDa. Black bars denote different sample groups. CH, cryopreserved hepatocytes; L, liver.
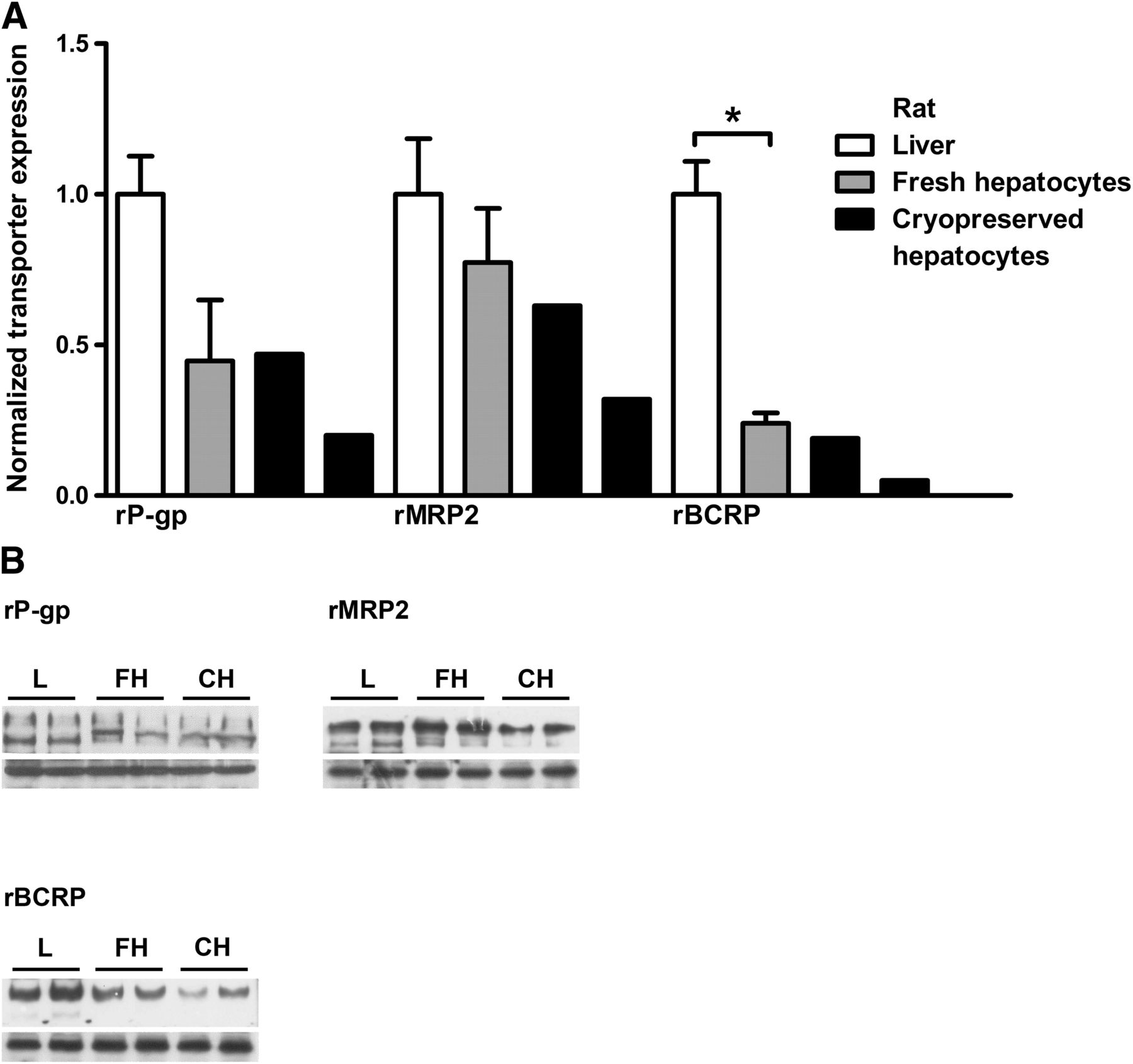
Three batches of fresh rat hepatocytes were prepared from Sprague-Dawley rats. Two batches of cryopreserved rat hepatocytes from the same strain were purchased from CellzDirect. In freshly isolated rat hepatocytes, rBCRP protein levels were significantly lower (75%) than in liver tissue (Fig. 2). In cryopreserved rat hepatocytes, the transporters rBCRP, rMRP2, and rP-gp were 90, 40, and 60% lower, respectively, compared with liver tissue.

(A) Quantification by Western blot of ABC efflux transporter expression in rat liver, freshly isolated hepatocytes, and cryopreserved cells. Transporter protein levels were normalized against GAPDH; expression levels in liver tissue were set to 1. rBCRP showed significant downregulation after isolation of cells of hepatocytes. All three transporters were downregulated after cryopreservation (mean ± SD, n = 3; *P < 0.05; for liver and fresh hepatocytes, for cryopreserved hepatocytes two individual measurements were performed). (B) Western blots of rP-gp, rMRP2, and rBCRP derived from rat liver, fresh hepatocytes, and cryopreserved hepatocytes are shown. Lower panels show the internal standard rGAPDH. Approximate molecular weights were rBCRP, 140 kDa (dimer); rMRP2, 190 kDa; rP-gp, 170 kDa; and rGAPDH, 35 kDa. Black bars denote different sample groups. CH, cryopreserved hepatocytes; FH, fresh hepatocytes; L, liver.
Immunofluorescent microscopy of cryopreserved human hepatocytes indicated that hBCRP, hMRP2, and hP-gp transporters were localized to the plasma membrane (Fig. 3). Some intracellular staining was also evident but was much weaker than the plasma membrane-associated signal. hMRP2 and hP-gp were found in distinct areas of the membrane possibly constituting remaining apical domains. hBCRP, in contrast, showed no clear localization but seemed distributed over the surface of the cryopreserved hepatocytes.

Immunofluorescent detection of hBCRP (A), hMRP2 (B), and hP-gp (C) in human cryopreserved hepatocytes. Right-hand panels show a higher magnification of individual cells. The large majority of transporters seemed to be localized to the plasma membrane, whereas intracellular staining was comparatively weak. hMRP2 and hP-gp were localized to distinct membrane domains reminiscent of intact canalicular membrane domains. hBCRP, in contrast, was found spread over a large membrane area showing no specific localization.
Immunofluorescent staining of human liver tissue sections by the same three primary antibodies toward hBCRP, hMRP2, and hP-gp (Fig. 4) showed clear and specific staining of the apical domains of hepatocytes. Localization of the transporters along the bile canalicular ducts was clearly visible.

Immunofluorescent detection of hBCRP (A), hMRP2 (B), and hP-gp (C) in human liver tissue sections. (D) A negative control section stained, omitting the primary antibody, is shown. Transporters are localized to apical membranes of hepatocytes tracing an outline of the bile canalicular network.
Efflux Transporter Activity in Human Cryopreserved and Rat Fresh Hepatocytes.
To investigate transporter functions in fresh and cryopreserved hepatocytes, the transport of rosuvastatin, a known BCRP substrate, and the mother compound-metabolite pair terfenadine-fexofenadine, both P-gp substrates, were used as probe compounds (Cvetkovic et al., 1999; Kitamura et al., 2008) in combination with the P-gp and BCRP inhibitor elacridar or the BCRP inhibitor FTC.
Figure 5 shows efflux of rosuvastatin from plated human cryopreserved hepatocytes (Fig. 5A) and rat fresh hepatocytes (Fig. 5B). Rosuvastatin efflux was in both cases inhibited by FTC. Efflux from rat cells was 20-fold higher than efflux from human cryopreserved hepatocytes, due to the higher uptake of rosuvastatin and resulting increase in intracellular concentration seen in fresh rat hepatocytes (data not shown).
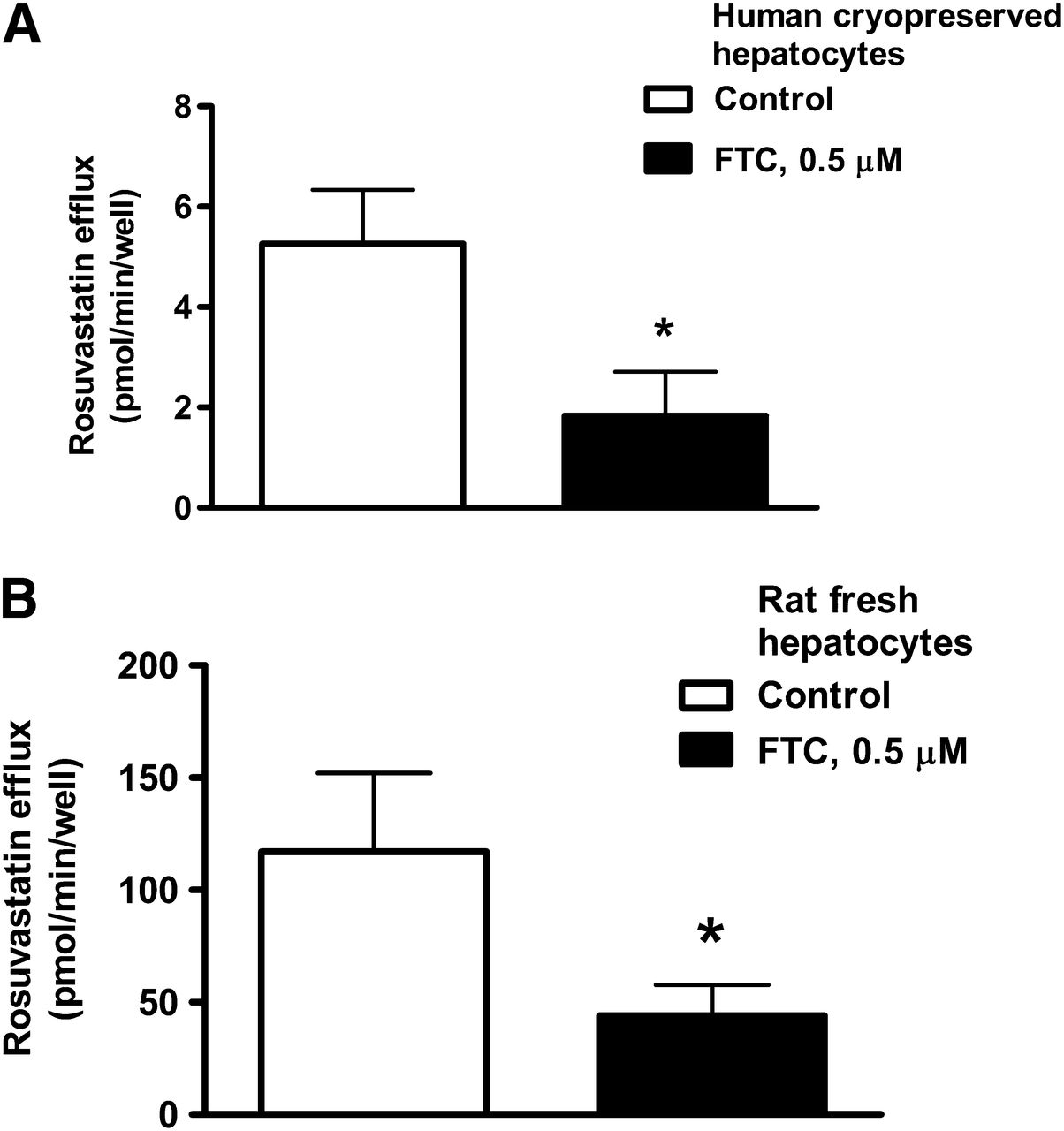
Efflux of rosuvastatin from plated hepatocytes can be inhibited with FTC. (A) Efflux from plated human cryopreserved hepatocytes. (B) Efflux from plated rat fresh hepatocytes (mean ± SD, n = 3; *P < 0.05).
In Figure 6A, the uptake and metabolism of terfenadine in human cryopreserved hepatocytes are shown; uptake was measured using the media loss method and metabolism by metabolic stability measurements (total metabolism, cells + medium) in suspended cells. The compound showed rapid uptake into hepatocytes and moderate metabolism. Adding 1 μM elacridar did not influence the rate of terfenadine metabolism in metabolic stability experiments (Fig. 6B), and the formation of the terfenadine metabolite fexofenadine was not affected (Fig. 6C).

Terfenadine uptake and metabolism and fexofenadine efflux in human cryopreserved hepatocytes. (A) Terfenadine shows rapid uptake (loss from media) and moderate metabolism (metabolic stability assay, cells + medium). (B) Metabolic breakdown of terfenadine is unaffected by 1 μM elacridar (metabolic stability assay, cells + medium). (C) Total formation of fexofenadine (cells + medium) is unaffected by 1 μM elacridar. (D) Efflux of fexofenadine is reduced by 40% upon addition of 1 μM elacridar. (E) Total formation of fexofenadine (cells + medium) in fresh rat hepatocytes, with and without 1 μM elacridar. (F) Efflux of fexofenadine from fresh rat hepatocytes, with and without 1 μM elacridar (mean ± SD, n = 3; **P < 0.01, ***P < 0.001).
Elacridar (1 µM) added to loss from media experiments with terfenadine reduced the efflux of intracellularily formed fexofenadine into the medium by approximately 40%, indicating inhibition of hP-gp–mediated efflux (Fig. 6D). In fresh rat hepatocytes, elacridar did not inhibit the formation of fexofenadine but inhibited the efflux in a similar way as shown in cryopreserved human hepatocytes (Fig. 6, E and F).
Uptake-Efflux Interactions.
To test whether efflux affects the results from loss from media uptake experiments, experiments with freshly isolated rat hepatocytes using the OATP and P-gp substrate fexofenadine were performed. The uptake of fexofenadine in human cryopreserved hepatocytes was studied using the spinning-through-oil technique because the uptake was too slow to be reliably measured in loss from media experiments.
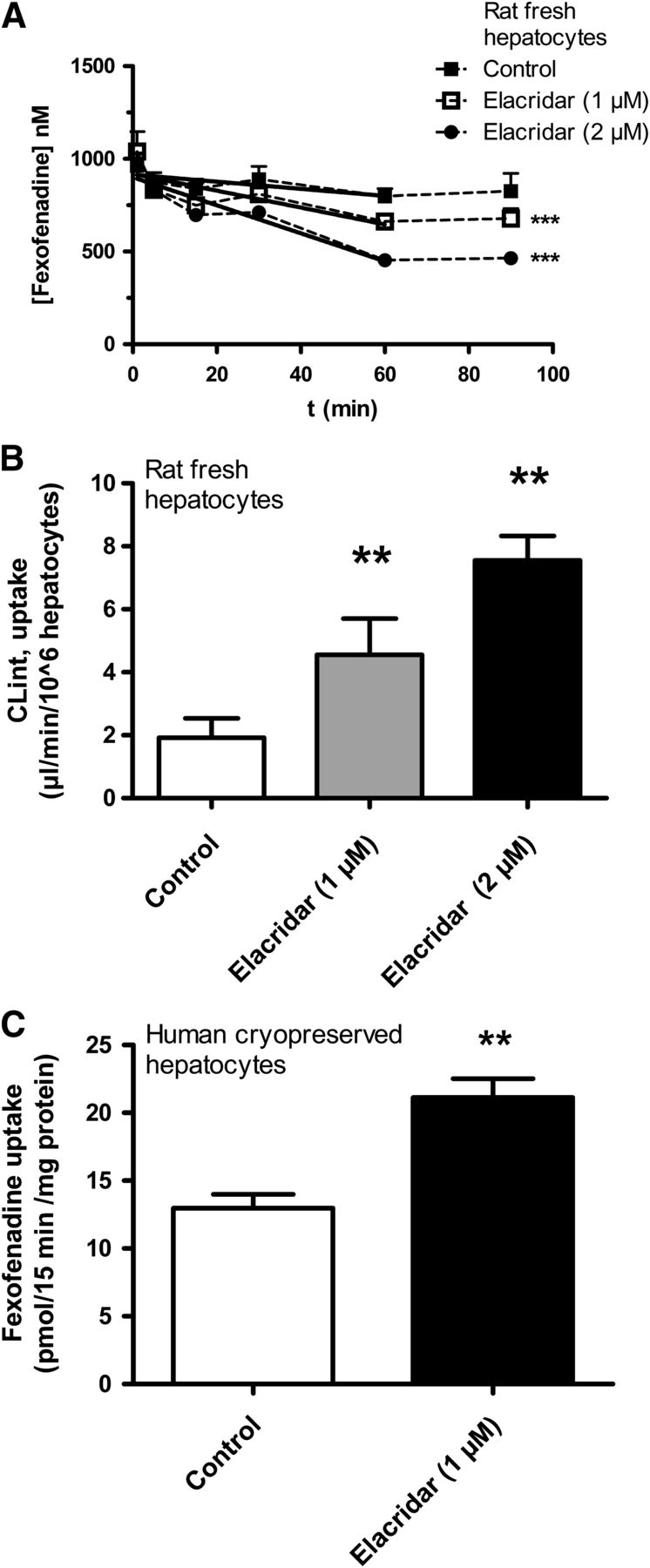
In freshly isolated rat hepatocytes, the measured loss of fexofenadine from the medium increased with increasing concentrations of elacridar up to 2 μM, indicating an active efflux of fexofenadine back to the medium by rP-gp (Fig. 7A). Solid lines represent initial fexofenadine uptake rates in the presence of 0, 1, and 2 μM elacridar (Fig. 7A). The apparent uptake rate increased 4-fold when comparing CLint, uptake in incubations with 2 μM elacridar and control experiments without elacridar (Fig. 7B).

Uptake-efflux interactions of fexofenadine in fresh rat hepatocytes. (A) In loss from media experiments, apparent uptake rates for fexofenadine increased with increasing concentrations of the P-gp inhibitor elacridar. Solid lines represent best fit curves of uptake rates. Steady state medium concentrations of fexofenadine seen after 60 minutes were decreased in a dose-dependent manner by elacridar. (B) Fexofenadine apparent uptake CLint increased 4-fold with increasing concentrations of elacridar from 0 to 2 μM. (C) In human cryopreserved hepatocytes, fexofenadine uptake was measured with the spinning-through-oil method. Fexofenadine uptake was significantly increased by 1 μM elacridar (mean ± SD, n = 3; **P < 0.01, ***P < 0.001).
Fig. 7A also indicates that the medium concentration of fexofenadine reached a steady state after 60 minutes. At this time point, the concentration of fexofenadine left in the medium was reduced with increasing elacridar concentration. Without elacridar, 20% of fexofenadine had been taken up by the cells after uptake completion. After addition of 2 μM elacridar, 55% of fexofenadine had been taken up by the hepatocytes after 60 minutes. We can thus infer that the total intracellular concentration of fexofenadine had more than doubled in the presence of 2 μM elacridar.
Measuring fexofenadine uptake in cryopreserved human hepatocytes with the spinning-through-oil method showed a significant increase in fexofenadine uptake after addition of 1 μM elacridar (Fig. 7C).
CLint, uptake measurements shown in Fig. 7B were used for IVIVE of rat hepatic fexofenadine CL (Table 1). The higher CLint, uptake values measured in the presence of elacridar led to predicted in vivo CLhepatic closer to observed in vivo values.
IVIVE of fexofenadine and erythromycin CLhepatic utilizing in vitro CLint, uptake measurements demonstrating the impact of elacridar
Metabolism-Efflux Interactions.
Effects of efflux inhibition on drug metabolism in human cryopreserved hepatocytes were investigated using erythromycin as a probe substrate together with 0.5 μM P-gp inhibitor elacridar.
In Figure 8, A and B, the results from a metabolic stability experiment with erythromycin are shown using human cryopreserved hepatocytes in suspension. The addition of 0.5 μM elacridar to the incubation did not change the total (cells + medium) metabolism of erythromycin significantly; neither was the total (cells + medium) formation rate of the main metabolite N-desmethyl-erythromycin affected.

Metabolism-efflux interactions of erythromycin in human cryopreserved hepatocytes. (A) Total metabolic breakdown (cells + medium) of erythromycin was unaffected by addition of 0.5 μM elacridar (metabolic stability assay). (B) Addition of 0.5 μM elacridar did not change the formation rate of the major metabolite N-desmethyl-erythromycin significantly (cells + medium, metabolic stability assay). (C) No change in extracellular erythromycin concentrations could be seen after addition of elacridar to spinning-through-oil experiments. (D) Efflux of the metabolite N-desmethyl-erythromycin was diminished by addition of elacridar (spinning-through-oil experiments). (E) Elacridar (0.5 μM) increased uptake and intracellular concentrations of erythromycin in human cryopreserved hepatocytes (spinning-through-oil experiments). (F) Intracellular levels of the metabolite N-desmethyl-erythromycin increased 2-fold in the presence of elacridar (s-t-o) (mean ± SD, n = 3; *P < 0.05, **P < 0.01, ***P < 0.001). AU, arbitrary units.
The extracellular concentration of erythromycin in spinning-through-oil uptake experiments with human cryopreserved hepatocytes was not affected by elacridar (Fig. 8C), whereas extracellular levels of N-desmethyl-erythromycin were reduced (Fig. 8D).
Measurements of intracellular levels showed that the net uptake of erythromycin was increased by approximately 2-fold after addition of elacridar (Fig. 8E). In parallel with this, intracellular concentrations of N-desmethyl-erythromycin more than doubled in the presence of the inhibitor (Fig. 8F).
Uptake of erythromycin was used for IVIVE of human hepatic CL (Table 1). The higher uptake rates measured in the presence of elacridar significantly improved the IVIVE.
The experiment on human cryopreserved hepatocytes shown in Figure 8 could be confirmed with fresh rat hepatocytes, reproducing the above findings of intracellular retention of erythromycin and its metabolite after P-gp inhibition by elacridar (data not shown).
Discussion
In the present study, protein levels of most tested efflux transporters, with the exception of hBCRP, were present at similar levels in human liver and cryopreserved hepatocytes. Our data correspond well with LC-MS/MS quantification of hepatocyte transporters by Li (et al., 2009a,b) showing retention of hMRP2, hBCRP, and hBSEP in human liver, fresh and cryopreserved hepatocytes. Li et al. (2009a,b) detected lower levels of rMRP2 in isolated and cryopreserved rat hepatocytes and a loss of rBCRP in cryopreserved cells, whereas in the present study rBCRP showed the largest reduction of the transporters studied in isolated as well as in cryopreserved hepatocytes as compared with fresh liver tissue levels.
The levels of the basolateral efflux transporters hMRP3 and hMRP4 were unchanged in cryopreserved human hepatocytes, compared with liver tissue. Basolateral efflux of drugs and conjugated drug metabolites is thus likely to be present at physiologic levels in in vitro experiments with isolated hepatocytes.
Retention of hepatocyte canalicular membrane domains after cell isolation has been demonstrated previously (Rippin et al., 2001; Tietz et al., 2005) and is likely representing the localization of hP-gp and hMRP2 observed in the present study. In contrast, hBCRP was found to be spread over large areas of the cell surface in isolated cells. This is reminiscent of the redistribution of some canalicular membrane markers such as alkaline phosphatase over the cell surface demonstrated by Groothuis et al. (1981). It seems that BCRP is both the least stable and more prone to lateral diffusion compared with the other tested ABC transporters. BCRP exists as a multimer of several “half-transporters,” whereas the other tested ABC transporters consist of single polypeptide chains, possibly making BCRP more prone to disassociation and breakdown (Klaassen and Aleksunes, 2010). Another explanation could be that P-gp, BSEP, and MRP2 are anchored to the cytoskeleton and are localized to stable caveolin-1 containing membrane microdomains (Hegedüs et al., 2003; Ortiz et al., 2004; Ismair et al., 2009). Such stabilizing interactions have not been demonstrated for BCRP, and their absence might make the transporter more sensitive to degradation. Some intracellular staining for hBCRP, hMRP2, and hP-gp was detected that may be internalized transporters or new transporter proteins that have yet to be exported to the membrane.
Bow et al. (2008) showed that rP-gp and rMRP2 in hepatocytes immediately after isolation were localized to remaining canalicular structures of the hepatocyte membrane. When culturing hepatocytes for 1 hour, a rapid but partial internalization of the transporters was observed, which was taken as evidence that transporter-mediated drug efflux is inactive in primary hepatocytes. However, little effort has been made to quantify the impact of this possible internalization on in vitro efflux activity. The fold reduction of efflux activity due to a loss of transporter proteins can be calculated as the inverse of the remaining transporter activity (Zamek-Gliszczynski et al., 2009). A 60% loss of BCRP activity in human cryopreserved or rat fresh hepatocytes would thus give a 2.5-fold (1/0.4) decrease in in vitro efflux and increase in intracellular exposure of a BCRP selective substrate, less if other efflux transporters are also involved. This is within the error margins deemed acceptable in IVIVE. BCRP expression in the human population is variable, showing a span of 2-fold around the mean, only slightly smaller than the reduction seen in cryopreserved samples (Prasad et al., 2013). This indicates that the remaining BCRP level is close to the levels in the lower range of the human population. In rat cryopreserved hepatocytes, effects will be larger, 10-, 1.7-, and 2.5-fold reductions in in vitro efflux for selective rBCRP, rMRP2, and rP-gp substrates, respectively.
Several other studies have suggested that efflux transporter protein and activity are retained to some extent in isolated or cryopreserved hepatocytes. Protein levels similar to liver expression levels of rBCRP, rBSEP, rP-gp, and rMRP2 have been detected in short-term cultures of isolated rat hepatocytes (Rippin et al., 2001; Tchaparian et al., 2011). Efflux activity in rat isolated hepatocytes has been shown for rP-gp using the inhibitor elacridar and the substrate erythromycin (Lam and Benet, 2004; Lam et al., 2006). Li and coworkers (2008) could demonstrate BCRP activity in human fresh and cryopreserved hepatocytes, but not rat cells, using the selective inhibitor FTC, and P-gp activity using efflux of 3,3-diethyloxacarbocyanine iodide (Li et al., 2008; Matsson et al., 2009). Full or partial retention of BCRP, BSEP, MRP2, and P-gp protein in isolated rat and human hepatocytes has been demonstrated by proteomics, but these measurements might include internalized transporters (Li et al., 2008, 2009a, 2009b). Retention of efflux transporter activity in human cryopreserved hepatocytes has recently been inferred from comparisons of CLint determinations from hepatocytes and microsomes for efflux transporter substrates (Di et al., 2012). These findings correspond well with the present study in which P-gp activity in rat fresh and human cryopreserved hepatocytes could be detected by inhibiting efflux of fexofenadine and rosuvastatin with the inhibitor elacridar and FTC, respectively. In contrast to Bow et al. (2008), only limited internalization of efflux transporters was seen in our experiments on human cryopreserved hepatocytes. Data in the present study are in agreement with studies suggesting that only a fraction of ABC transporters is internalized, leaving substantial active efflux in freshly isolated as well as cryopreserved hepatocytes (Li et al., 2009a, 2009b; Di et al., 2012).
Efflux transporter activity may reduce intracellular concentrations of certain drugs and thus have an impact on both uptake rate estimations and drug metabolism in the hepatocytes, often called transporter-enzyme interplay. In vivo, in the liver, uptake and efflux transporters work in unison, which results in a vectorial flux of compounds from blood to bile. When performing in vitro hepatocyte uptake studies with suspended hepatocytes, the cell polarization is lost. Compound that is taken up by uptake transporters is effluxed back into the same medium compartment by active ABC transporters that in this manner diminishes measurements of apparent uptake (Benet, 2009). In the present study, measured CLint, uptake for fexofenadine in rat hepatocytes was increased after addition of the P-gp inhibitor elacridar. Fexofenadine is also effluxed by MRP2, so efflux inhibition by elacridar was likely not complete (Matsushima et al., 2008; Tian et al., 2008b; Matsson et al., 2009). Fexofenadine shows low permeability, low metabolism, and high solubility, and its hepatic CL is most likely rate limited by uptake transporters (Benet, 2009). Scaling rat in vivo hepatic CL for fexofenadine from hepatocyte CLint, uptake data resulted in a large underprediction. Predictions of fexofenadine hepatic CL were considerably improved using CLint, uptake generated in the presence of elacridar, demonstrating that in vitro uptake measurements can be perturbed by efflux transporter activity retained in the hepatocytes used. Scaling of human CLhepatic from erythromycin uptake in human cryopreserved hepatocytes was improved in a similar manner using the higher uptake rates measured in the presence of elacridar. Human hepatic CL was underpredicted 4- to 5-fold, presumably due to reduced uptake of erythromycin in human cryopreserved hepatocytes as a consequence of the large downregulation of OATPs seen in human cryopreserved hepatocytes (Kimoto et al., 2012; Lundquist et al., 2014).
Uptake transporters are rate determining for the clearance of many drugs, but IVIVE of CLhepatic from hepatocyte uptake data generally underpredicts the in vivo CL (Chiba et al., 2009; Watanabe et al., 2009; Giacomini et al., 2010). Modeling of in vivo CL from rat and human hepatocyte uptake data in recent studies needed inclusion of empirical scaling factors ranging from 2 to 67 to predict in vivo hepatic CL within 2-fold (Gardiner and Paine, 2011; Menochet et al., 2012). One reason for this underprediction may be that the studies did not take any active efflux transporters in the cell models into account when predicting CLhepatic. Additionally, loss of OATPs and other uptake transporters when isolating and cryopreserving rat and human hepatocytes may further add to the underprediction of hepatic CL (Kimoto et al., 2012; Lundquist et al., 2014).
In the present study, no increase in erythromycin metabolism could be detected in experiments using cryopreserved human hepatocytes after addition of elacridar. The addition of elacridar slightly diminished the total amount of N-desmethyl-erythromycin metabolite formed. However, the intracellular levels of both erythromycin and the N-desmethyl-erythromycin metabolite increased after P-gp inhibition, which is in accordance with the results by Lam et al. (2006). Measurements of metabolite concentrations in the extracellular medium showed that P-gp inhibition had reduced the efflux of the metabolite that previously was identified as a P-gp substrate (Lam et al., 2006). The increase in intracellular metabolite levels is thus probably to a large extent a result of inhibited efflux of the metabolite and not a result of increased metabolism.
In conclusion, the present study shows that the four ABC efflux transporters, P-gp, BCRP, MRP2, and BSEP, important for biliary drug clearance, are retained in human cryopreserved and freshly isolated rat hepatocytes. The transporters exhibit activity that clearly affects the drug disposition in the experimental system and should thus be considered when hepatocyte drug metabolism and transport kinetics are studied.
Acknowledgments
The technical assistance of Annelie Bengtsson, Jessie Dahlström, and Eva Floby is gratefully acknowledged. We thank Dr. Mike Baldwin, University of California, San Francisco, for generously providing us with the MRP4 antibody.
Authorship Contributions
Participated in research design: Lundquist, Englund, Skogastierna, Lööf, Andersson.
Conducted experiments: Lundquist, Englund, Skogastierna, Johansson, Lööf.
Contributed new reagents or analytic tools: Johansson.
Performed data analysis: Lundquist, Englund, Skogastierna, Johansson, Lööf.
Wrote or contributed to the writing of the manuscript: Lundquist, Hoogstraate, Afzelius, Andersson.
Footnotes
- Received August 26, 2013.
- Accepted January 6, 2014.
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- ABC
- ATP-binding cassette
- BCRP
- breast cancer–resistance protein
- BSEP
- bile salt efflux pump
- CL
- clearance
- CLhepatic
- hepatic clearance
- CLint
- intrinsic clearance
- FCS
- fetal calf serum
- FTC
- fumitremorgin C
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- h
- as prefix, human
- HRP
- horseradish peroxidase
- IVIVE
- in vitro–in vivo extrapolation
- KHL
- Krebs-Henseleit buffer
- LC-MS/MS
- liquid chromatography tandem mass spectrometry
- MRP
- multidrug resistance-associated protein
- OATP
- organic anion-transporting polypeptide
- PBS
- phosphate-buffered saline solution
- P-gp
- P-glycoprotein
- P450
- cytochrome P450
- r
- as prefix, rat
- TBST
- Tris-buffered saline solution, 0.1% Tween 20
- WE
- William’s E medium
- Copyright © 2014 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}