


Abstract
In our study, we tested the hypothesis whether valproic acid (VPA) in therapeutic concentrations has potential to affect expression of CYP3A4 and MDR1 via constitutive androstane receptor (CAR) and pregnane X receptor (PXR) pathways. Interaction of VPA with CAR and PXR nuclear receptors was studied using luciferase reporter assays, real-time reverse transcriptase polymerase chain reaction (RT-PCR), electrophoretic mobility shift assay (EMSA), and analysis of CYP3A4 catalytic activity. Using transient transfection reporter assays in HepG2 cells, VPA was recognized to activate CYP3A4 promoter via CAR and PXR pathways. By contrast, a significant effect of VPA on MDR1 promoter activation was observed only in CAR-cotransfected HepG2 cells. These data well correlated with up-regulation of CYP3A4 and MDR1 mRNAs analyzed by real-time RT-PCR in cells transfected with expression vectors encoding CAR or PXR and treated with VPA. In addition, VPA significantly up-regulated CYP3A4 mRNA in primary hepatocytes and augmented the effect of rifampicin. EMSA experiments showed VPA-mediated augmentation of CAR/retinoid X receptor α heterodimer binding to direct repeat 3 (DR3) and DR4 responsive elements of CYP3A4 and MDR1 genes, respectively. Finally, analysis of specific CYP3A4 catalytic activity revealed its significant increase in VPA-treated LS174T cells transfected with PXR. In conclusion, we provide novel insight into the mechanism by which VPA affects gene expression of CYP3A4 and MDR1 genes. Our results demonstrate that VPA has potential to up-regulate CYP3A4 and MDR1 through direct activation of CAR and/or PXR pathways. Furthermore, we suggest that VPA synergistically augments the effect of rifampicin in transactivation of CYP3A4 in primary human hepatocytes.
Valproic acid (VPA) is an effective broad-spectrum anticonvulsant used in the treatment of primary generalized tonic-clonic, absence, and partial seizures (Tanaka, 1999). VPA has recently been identified as an inhibitor of histone deacetylase (HDAC) with potential antitumorous activity that has been studied in several clinical trials (Göttlicher et al., 2001; Blaheta et al., 2002, 2005; Kramer et al., 2003; Raffoux et al., 2005).
VPA is well known to affect mechanisms that control drug disposition such as activity of hepatic biotransformation enzymes or drug binding to plasma proteins (Rogiers et al., 1995; Wen et al., 2001; Perucca, 2006). Generally, VPA is thought to be an inhibitor rather than an inducer of drug-metabolizing enzymes (Perucca, 2006). However, this point of view is currently not supported satisfactorily by comprehensive data published in the literature. Regarding metabolizing enzymes of the cytochrome P450 family, Wen et al. (2001) have demonstrated, using in vitro methods, that VPA in therapeutically relevant concentrations competitively inhibits only human CYP2C9 catalytic activity. On the other hand, there is clear evidence that VPA has potential to up-regulate expression and activity of several rodent and human genes encoding proteins involved in drug disposition. It was found that prolonged exposure of rats to VPA results in the self-inducing metabolism of the agent (Fisher et al., 1991). Moreover, Rogiers et al. (1992, 1995) have found VPA to be a potent inducer of genes of the rat Cyp2b subfamily, in particular, Cyp2b1 and Cyp2b2. Recently, Eyel et al. (2006) have found out that VPA does not affect expression of rat Cyp3a2, an ortholog of human CYP3A4, whereas valpromide, the primary amide of VPA that reveals no HDAC-inhibitory activity, was shown to induce this gene by a nonspecific mechanism. In addition, these authors have suggested that VPA induces expression and activity of human P-glycoprotein (MDR1), a member of the ATP-binding cassette family of drug transporters, in tumor cell lines by the mechanism of HDAC inhibition (Eyal et al., 2006). These summarized data indicate that VPA is capable of altering expression and activity of various cytochromes P450 differently. Moreover, VPA has potential to induce the MDR1 gene.
We assumed that alteration of expression of these genes could be caused by interaction of VPA with constitutive androstane receptor (CAR; N1I3) and human pregnane X receptor (PXR; SXR; NR1I2). PXR and CAR are ligand-activated nuclear receptors that act as heterodimers with retinoid X receptor α (RXRα) and up-regulate the transcription of their target genes, such as CYP3A4 and MDR1, by interaction with specific promoter-binding motifs (Goodwin et al., 1999). PXR has been shown to be activated by many structurally and chemically diverse ligands. Examples of human PXR activators include xenobiotics such as rifampicin (Bertilsson et al., 1998; Pascussi et al., 2000), the endobiotic lithocholic acid (Staudinger et al., 2001; Xie et al., 2001), the progesterone metabolite 5-pregnane-3,20-dione (Jones et al., 2000), and the herbal compound hyperforin (Moore et al., 2000a). In the case of CAR, the experimental substance CITCO ((6-(4-chlorophenyl) imidazo [2,1-b][1,3] thiazole-5-carbaldehyde O-(3,4-dichlorobenzyl) oxime), the antimalarial artemisinin drugs, and 5-pregnane-3,20-dione have been recognized as its ligands (Jones et al., 2000; Maglich et al., 2003; Burk et al., 2005b). The barbiturate drug phenobarbital activates CAR indirectly through the increase of CAR translocation from the cytoplasm to the nucleus (Honkakoski et al., 1998; Kawamoto et al., 1999; Goodwin and Moore, 2004). On the other hand, several competitive inverse agonists of CAR have also been discovered such as androstanol, androstenol, and clotrimazole (Forman et al., 1998; Moore et al., 2000b).
In this study, we examined whether VPA controls expression of CYP3A4 and MDR1 (ABCB1) genes at the transcriptional level via activation of PXR and CAR pathways. This hypothesis was investigated in the human Caucasian hepatocyte carcinoma (HepG2) and human colon carcinoma (LS174T) cells using several reporter luciferase constructs with major promoter-regulatory sequences of CYP3A4 and MDR1. In addition, using real-time RT-PCR, we investigated CYP3A4 mRNA expression in primary human hepatocytes treated with VPA. Moreover, we studied whether VPA augments binding of CAR/RXRα to several response elements of CYP3A4 (DR3, ER6) and MDR1 (DR4) using the electrophoretic mobility shift assay (EMSA).
Our data indicate that VPA is capable of transactivating both CYP3A4 and MDR1 via interaction with the CAR pathway; however, VPA-mediated activation of PXR pathway controls only CYP3A4 gene expression. Moreover, we demonstrate that VPA synergizes with rifampicin in transactivation of CYP3A4 in primary human hepatocytes.
Materials and Methods
Cell Lines and Chemicals. The human Caucasian hepatocyte carcinoma HepG2 and human colon adenocarcinoma LS174T cell lines were purchased from the European Collection of Cell Cultures (Salisbury, UK) and were used within 25 passages after delivery and maintained in antibiotic-free Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum (FCS), 1% sodium pyruvate, and 1% nonessential amino acids (Sigma-Aldrich, St. Louis, MO). The latter cell line has been previously shown to have highly inducible expression of CYP3A4 and MDR1 (Geick et al., 2001).
Androstenol (5α-androst-16-en-3α-ol), rifampicin, VPA (2-propylpentanoic acid), and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich. CITCO was purchased from BIOMOL Research Laboratories (Plymouth Meeting, PA). DMSO or water was used as a solvent of VPA when indicated.
Plasmids. A chimeric p3A4-luc reporter construct containing the basal promoter (–362/+53) with proximal PXR response element and the distal xenobiotic responsive enhancer module (–7836/–7208) of the CYP3A4 gene 5′-flanking region inserted to pGL3-Basic reporter vector was described by Goodwin et al. (1999). Plasmid p-7975(Δ7012–1804) harboring the enhancer of MDR1 gene 5′-flanking region (MDR1E) and the basal promoter were constructed with pGL3-Basic reporter vector as described by Geick et al. (2001), herein referred to as pMDR1-luc. pMDR1E-SV40-luc reporter plasmid was constructed by insertion of the enhancer region of MDR1 gene from –7881 to –7809 into pGL3-Promoter plasmid (Promega, Madison, WI) in NheI/BglII cloning sites upstream of a heterologous promoter SV40. CYP2B6 phenobarbital responsive enhancer module (PBREM) reporter gene construct (pPBREM-SV40-luc) containing two DR4-type motifs (NR1 and NR2) was prepared from pGL3-Promoter plasmid by insertion of the region from –1733 to –1683 of CYP2B6 promoter into NheI/BglII cloning sites upstream of a heterologous promoter SV40. Constructed plasmids were sequenced using a Big Dye Terminator Cycle Sequencing Method (Applied Biosystems, Foster City, CA). The expression plasmid for human PXR receptor, pSG5-PXR, was kindly provided by Dr. S. Kliewer (University of Texas, Dallas, TX). The human CAR expression plasmid pCR3-CAR was kindly provided by Dr. M. Negishi (National Institute of Environmental Health Sciences, Research Triangle Park, NC). The expression plasmid pSG5-hRXRα encoding hRXRα cDNA was a generous gift from Dr. C. Carlberg (University of Kuopio, Kuopio, Finland).
Transient Transfection and Luciferase Gene Reporter Assays. All transfection assays were carried out using Lipofectamine2000 transfection reagent (Invitrogen, Carlsbad, CA) in cells cultivated in the phenol red-free medium containing 10% charcoal-stripped FCS, 1% sodium pyruvate, and 1% nonessential amino acids according to the manufacturer's instruction.
HepG2 cells (2 × 105 per well) were seeded into 48-well plates and cotransfected with a luciferase reporter construct (0.4 μg/well) and expression plasmid encoding either CAR or PXR (50 ng/well) 24 h later. Subsequently, cells were maintained in medium supplemented with 10% charcoal/dextranstripped fetal bovine serum containing VPA at appropriate concentrations for 24 h. Luminescence activity was determined with a Genios Plus luminometer (Tecan, Grödig, Austria) in cell lysate using a commercially available luciferase detection system (Promega). In addition, all experiments were performed in parallel using empty pGL3-Basic and pGL3-Promoter luciferase reporter constructs containing no responsive elements to distinguish nonspecific CAR- and PXR-independent effect of VPA on reporter plasmids used. Luminescence of these samples was taken as background that was subtracted in the final calculation from luminescence of cell samples transiently transfected with reporter plasmids containing response elements of tested genes. Resulting data are presented as ratio of luminescence of treated cell samples to control. Luminescence of each sample has been normalized to its protein concentration determined with the BCA system (Pierce, Rockford, IL).
Real-Time RT-PCR Analysis of CYP3A4 mRNA. LS174T cells (1.2 × 105 per well) were seeded into 24-well plates and cultivated for 24 h. Then cells were transfected with CAR or PXR expression plasmids (400 ng/well) and appropriate cell samples were exposed to VPA at a concentration of 500 μM for 48 h. Total RNA was isolated using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. The following primers were used for CYP3A4: forward primer 5′-TTCAGCAAGAAGAACAAGGACAA-3′, reverse primer 5′-GGTTGAAGAAGTCCTCCTAAGC-3′); for MDR1, forward primer 5′-TGCTCAGACAGGATGTGAGTTG-3′, reverse primer 5′-AATTACAGCAAGCCTGGAACC-3′; and for housekeeping genes HPRT (hypoxanthine-guanine phosphoribosyl transferase), forward primer 5′-CTGGAAAGAATGTCTTGATTGTGG-3′, reverse primer 5′-TTTGGATTATACTGCCTGACCAAG-3′ and B2M (β2-microglobulin), forward primer 5′-CGTGTGAACCATGTGACTTTGTC-3′, reverse primer 5′-CATCTTCAAACCTCCATGATGC-3′). cDNA was prepared from 1 μg of total RNA with MMLV transcriptase (Finnzymes, Espoo, Finland) using oligo(dT)15VN primer (Generi-Biotech, Hradec Králové, Czech Republic) and porcine RNase inhibitor (TaKaRa BIO, Shiga, Japan). Real-Time PCR analysis was performed on an iCycler (Bio-Rad, Hercules, CA). cDNA (40 ng of reverse-transcribed RNA) was amplified with HotStar Taq polymerase (QIAGEN, Valencia, CA) under the following conditions: 3 mM MgCl2, 0.2 mM deoxynucleoside-5′-triphosphate, 0.3 μM each primer, 0.025 U/μl polymerase, SYBR Green I in 1:100,000 dilution, and fluorescein (10 nM). The temperature profile was 95°C for 14 min; 50 times 95°C for 15 s, 60°C for 20 s, 72°C for 30 s, 72°C for 15 s; melting curve program 70–95°C. All samples were run in triplicates simultaneously with negative controls. The processing of real-time amplification curves was performed on iCycler software version 4.6 (Bio-Rad). Pfaffl′s method was applied for relative quantification of gene expression normalized to endogenous control (housekeeping) gene (Pfaffl, 2001). Results are presented as the means of at least three experiments.
Isolation and CYP3A4 mRNA Analysis in Primary Cell Culture of Human Hepatocytes. Human tissue samples were obtained according to protocols approved by the local ethics committee of the Palacky University in Olomouc (Czech Republic) complying with the current Czech legislation. Hepatocytes were prepared from lobectomy segments resected from adult patients for medical reasons unrelated to our research program. The tissue encompassing the tumor was dissected by a surgeon and sent for anatomopathological studies, whereas the remaining tissue was used for hepatocyte preparation. No information on the patients was available to us, apart from age, sex, and the reason for surgery. Human liver samples used in this study were obtained from two donors: donor 1, a woman, 69 years old, tumor metastasis; and donor 2, a woman, 46 years old, Caroli syndrome. Hepatocytes were isolated as described previously (Pichard-Garcia et al., 2002). After isolation, the cells were plated on collagen-coated culture dishes at density 1.4 × 105 cells/cm2. Culture medium was enriched for plating with 2% FCS (v/v) as described previously (Isom et al., 1985). The medium was replaced with serum-free medium the day after, and the cells were allowed to be stabilized for an additional 48 to 72 h before the treatment with VPA (500 μM) or rifampicin (10 μM) or their combination. Cells were maintained at 37°C and 5% CO2 in a humidified incubator. The effect of VPA on CYP3A4 mRNA expression was tested in periods of 24 and 48 h. The effect of rifampicin and the combination of VPA and rifampicin were studied in a period of 24 h. In the latter experiments, DMSO (0.1%) was used as solvent of both VPA and rifampicin. The level of CYP3A4 mRNA expression was analyzed using real-time RT-PCR according to the protocol mentioned above.
Functional Analysis of CYP3A4 Activity. LS174T cells (6 × 105 per well) were seeded into 12-well plates and cultivated for 24 h. Subsequently, cells were transfected with CAR or PXR expression plasmids (1.2 μg/well) and then exposed to VPA at a concentration of 600 μM for 48 h. After treatment, cells were washed with phosphate-buffered saline and cultivated in serum-free Opti-MEM medium (Invitrogen-Gibco) containing testosterone at a final concentration of 150 μM for 2 h at 37°C. Then, cells were washed with phosphate-buffered saline and lysed with 200 μl of SDS (1%). CYP3A4 activity was detected in total cellular lysate (150 μl) using an established method based on the measuring of a prototypic CYP3A4-mediated testosterone 6β-hydroxylation activity (Guengerich et al., 1986) and modified for the analysis of the metabolite in cell lysate. For the determination of CYP3A4 activity, the metabolite was extracted to dichloromethane. Subsequently, the solvent was evaporated and the sample dissolved in the mobile phase. The concentration of the testosterone metabolite was determined using a Shimadzu Class VP HPLC system (Shimadzu, Tokyo, Japan). Final activity was normalized to sample protein concentration in cell lysate that was measured using the BCA assay according to the manufacturer's instructions (Pierce).

Effect of VPA on CAR- and PXR-mediated transactivation of CYP3A4 promoter. HepG2 cells were transiently transfected with p3A4-luc reporter construct containing the basal promoter (–362/+53) with proximal PXR response element and the distal xenobiotic responsive enhancer module (–7836/–7208) of CYP3A4 (0.4 μg/well) and either pCR3-CAR or pSG5-PXR expression vector (50 ng) using Lipofectamine2000 transfection reagent according to the manufacturer's instructions. Transfected HepG2 cells were maintained in medium containing VPA at the indicated concentrations for 24 h. Luciferase activities are normalized to protein concentration and expressed as -fold activation of nontreated cells transfected with p3A4-luc. All means ± S.D. were calculated from quadruplicates of a representative experiment and analyzed using ANOVA followed by Dunnett's test. #, p < 0.05; ###, p < 0.001: statistically different from nontreated cells transfected with p3A4-luc. *, p < 0.05; **, p < 0.01: statistically different from VPA-nontreated cells cotransfected with either pCR3-CAR or pSG5-PXR.
EMSA. Human CAR and RXRα were translated in vitro using TNT Quick Coupled Transcription/Translation Systems (Promega, Southampton, UK). The nuclear fraction was isolated from HepG2 cells transfected with hRXRα expression vector using CelLytic NuCLEAR Extraction Kit (Sigma). The following double-stranded 5′-biotinylated oligonucleotides of specific response elements of both CYP3A4 and MDR1 promoters were used as probes: CYP3A4 DR3 sense, 5′-GAATGAACTTGCTGACCCTCT-3′; CYP3A4 DR3 antisense, 5′-AGAGGGTCAGCAAGTTCATTC-3′; CYP3A4 ER6 sense, 5′-ATATGAACTCAAAGGAGGTCAGTG-3′; CYP3A4 ER6 antisense, 5′-CACTGACCTCCTTTGAGTTCATAT-3′; MDR1 DR4 sense, 5′-CATTGAACTAACTTGACCTTGC-3′; and MDR1 DR4 antisense, 5′-GCAAGGTCAAGTTAGTTCAATG-3′. The oligonucleotides were synthesized at Generi-Biotech.
The EMSA was performed according to the protocol published previously with slight modifications (Frank et al., 2003). The binding reactions were performed in a total volume of 20 μl and contained equal amounts (∼3 μg) of CAR and RXRα, 10× binding buffer (100 mM Tris, 500 mM KCl, 10 mM dithiothreitol) (Pierce), poly(dI · dC) Double Strand (GE Healthcare, Little Chalfont, Buckinghamshire, UK), and 1% Nonidet P-40. VPA was tested at final concentrations of 100, 500, and 1000 μM. CITCO, an agonist of human CAR, was used at the concentration of 6.25 μM and androstenol at the concentration of 10 μM. DMSO was used as a solvent of the compounds at maximal final concentration of 0.1%. For supershift experiments, 1 μg of the anti-RXRα rabbit polyclonal IgG antibody was added to the reaction mixture (Santa Cruz Biotechnology, Inc., Santa Cruz, CA). Reactions were preincubated on ice for 10 min before the addition of oligonucleotide probe (20 fmol). Samples were maintained at room temperature for an additional 20 min, and then protein/DNA complexes were resolved on a 5% (w/v) nondenaturing polyacrylamide gel (acrylamide/bisacrylamide 29:1 w/v) in 0.5× Tris borate-EDTA buffer (450 mM Tris, 450 mM boric acid, 10 mM EDTA). The gels were electrophoresed at 100 V for an hour at room temperature. Next, the gels were blotted at 380 mA for 30 min at 4°C and then analyzed by the Chemiluminescent Nucleic Acid Detection Module (Pierce).
EMSA Quantification. EMSA reactions were exposed to X-ray film (Foma Bohemia a.s., Hradec Králové, Czech Republic) and bands were quantified using computerized densitometry using LabImage densitometry analysis software (Kapelan Bio-Imaging Solutions, Halle, Gemany).
Statistics. One-way ANOVA followed by Dunnett's multiple comparison post hoc test or unpaired Student's t test was used for statistical analysis of data using GraphPad Prism software (GraphPad Software Inc., San Diego, CA). Two-way ANOVA with interaction was used to analyze the synergistic effect of VPA and rifampicin.
Results
Effect of VPA on CAR- and PXR-Mediated Activation of CYP3A4 Promoter in HepG2 Cells Transiently Transfected withp3A4-lucLuciferase Reporter Construct. First, we examined whether VPA affects activation of p3A4-luc reporter construct through CAR and PXR. We observed statistically significant concentration-dependent increase in CAR- and PXR-mediated transcriptional activation of p3A4-luc in HepG2 cells after 24 h exposure to VPA (Fig. 1). VPA at the concentration of 500 μM doubled the activation of p3A4-luc in cells cotransfected with CAR expression vector (Fig. 1). In HepG2 cells cotransfected with PXR, we observed even a 4-fold increase in activation of p3A4-luc after treatment with VPA (500 μM) (Fig. 1). In contrast, VPA did not significantly affect transcriptional activation of p3A4-luc in HepG2 cells, which were not cotransfected with either CAR or PXR (Fig. 1). We observed that expression of both CAR and PXR in HepG2 cells in the absence of an exogenous ligand resulted in statistically significant activation of p3A4-luc reporter, which is in agreement with published data and indicates involvement of endogenous activators of the nuclear receptors in HepG2 cells (Fig. 1) (Goodwin et al., 1999, 2002).

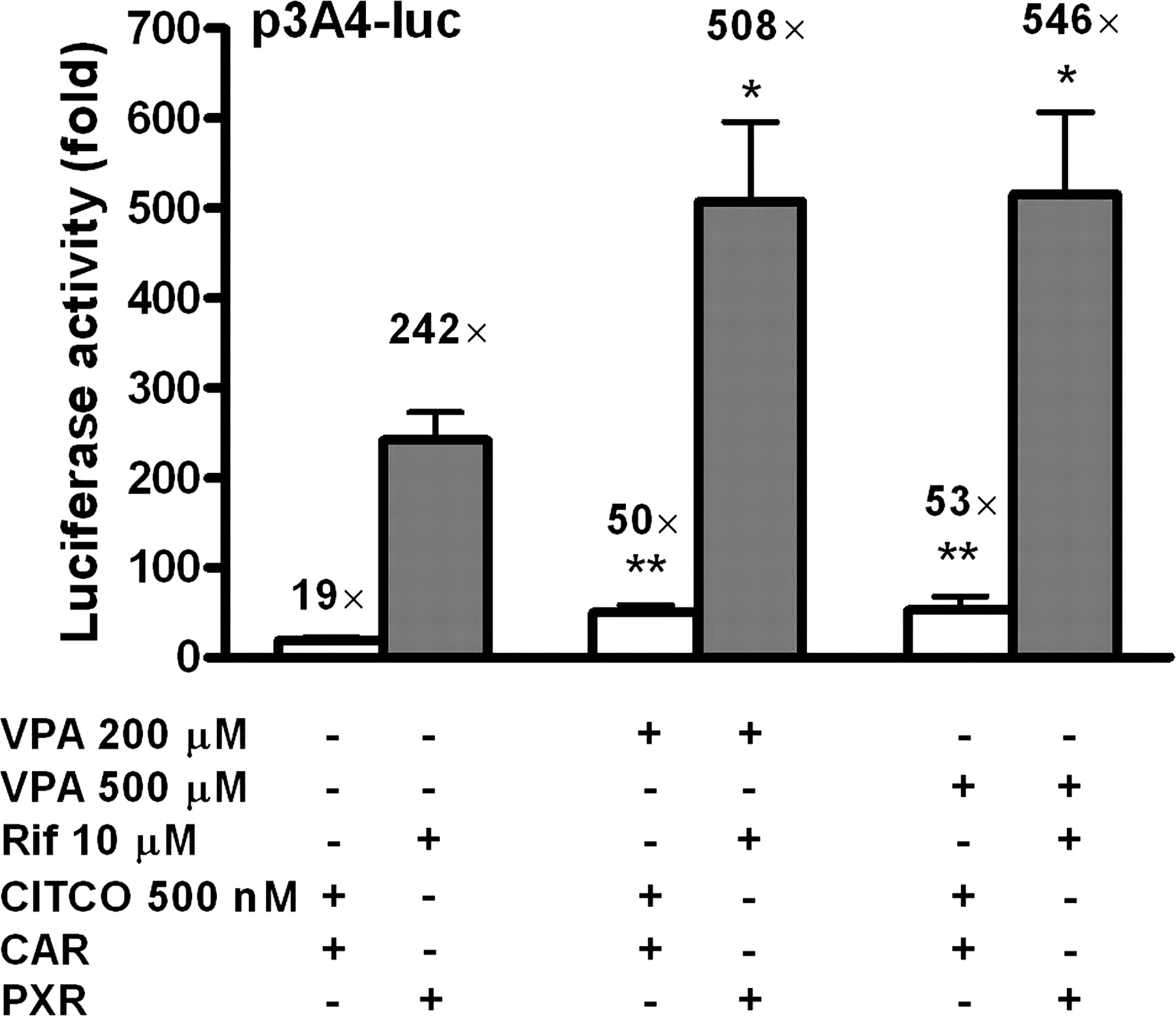
Transactivation of CYP3A4 promoter by VPA in combination with CITCO or rifampicin. HepG2 cells were transiently transfected with p3A4-luc reporter construct (0.4 μg/well) and pCR3-CAR or pSG5-PXR expression plasmid (50 ng) using Lipofectamine2000 transfection reagent according to the manufacturer's instructions and subsequently exposed to VPA in combination with CITCO (500 nM) or rifampicin (Rif; 10 μM), prototype ligands of CAR and PXR, respectively, for 24 h. All means ± S.D. were calculated from quadruplicates of a representative experiment and analyzed using ANOVA followed by Dunnett's test. Data are presented as -fold activation of nontreated cells transfected only with p3A4-luc. *, p < 0.05; **, p < 0.001: statistically different from cells cotransfected with either pCR3-CAR or pSG5-PXR and treated with either CITCO or rifampicin.
Transactivation of CYP3A4 Promoter by VPA in Combinationwith CITCO or Rifampicin. In these experiments, we examined whether VPA has the ability to affect CAR- and PXR-mediated activation of p3A4-luc construct caused by prototypic ligands CITCO (500 nM) in CAR-expressing cells and rifampicin (10 μM) in PXR-expressing cells. VPA at tested concentrations in combination with CITCO (500 nM) significantly augmented CAR-mediated activation of p3A4-luc compared with CAR-expressing cells treated with CITCO (p < 0.01) (Fig. 2). Similarly, we observed a more pronounced effect of VPA in combination with rifampicin (10 μM) on transcriptional activation of p3A4-luc via PXR compared with the effect of rifampicin alone (p < 0.05) (Fig. 2).
Effect of VPA on CAR- and PXR-Mediated Activation of MDR1 Promoter in HepG2 Cells Transiently Transfected withpMDR1-lucor HeterologouspMDR1E-SV40-lucLuciferase Reporter Constructs. Regarding MDR1, two different luciferase reporter constructs were used in our study. First, we used pMDR1-luc construct containing the distal enhancer region with a DR4 responsive motif common to both PXR- and CAR-mediated transactivation of MDR1 gene, and the native basal promoter from –1803 to +281. Using this reporter construct, we observed a similar effect of VPA on activation of MDR1 promoter in cells cotransfected with PXR or CAR in comparison with mock-transfected HepG2 cells (Fig. 3A). On the other hand, rifampicin (10 μM) and CITCO (1 μM) significantly activated (∼2-fold, p < 0.05) the pMDR1-luc construct in HepG2 cells cotransfected with PXR or CAR expression plasmids under the experimental conditions used. This indicates that VPA probably has potential to transactivate this luciferase construct independently on PXR and CAR pathways through a nonspecific mechanism, which is likely related to activation of MDR1 basal promoter (Morrow and Nakagawa, 1994).
In light of our previous observations demonstrating that heterologous pMDR1E-SV40-luc is more responsive to both PXR- and CAR-mediated activation than pMDR1-luc reporter, we performed a transfection assay with pMDR1E-SV40-luc reporter construct containing the MDR1 enhancer upstream of viral SV-40 promoter. Using the plasmid, we observed that VPA at a concentration of 500 μM activates significantly only CAR-mediated transcription of pMDR1E-SV40-luc reporter construct (p < 0.001) (Fig. 3B). The effect of VPA on activation of pMDR1E-SV40-luc plasmid was comparable to the effect of CITCO (1 μM, 2-fold activation) and rifampicin (25 μM, 2.3-fold activation, p < 0.05) in nuclear receptor-transfected cells.
Finally, we used androstenol, an inverse agonist of CAR, to confirm that VPA activates pMDR1E-SV40-luc through CAR. Androstenol (10 μM) significantly repressed CAR-mediated transcriptional activation of pMDR1E-SV40-luc in cells exposed to VPA at a concentration of 500 μM (p < 0.05) (Fig. 3C). In contrast, we observed no effect of androstenol on pGL3-Basic or pGL3-Promoter construct activities in cells cotransfected with CAR expression vector (data not shown).
Effect of VPA on CAR- and PXR-Mediated Activation of CYP2B6 PBREM in HepG2 Cells Transiently Transfected withpPBREM-SV40-lucLuciferase Reporter Construct. To elucidate the discrepancy between CAR- and PXR-mediated activation of MDR1 promoter by VPA, we analyzed luciferase activity of pP-BREM-SV40-luc construct in cells transiently transfected with CAR or PXR expression vectors and exposed to VPA (500 μM). pPBREM-SV40-luc construct contains two DR4-type responsive elements of the PBREM of the CYP2B6 gene, which has high ability to interact with both CAR and PXR (Goodwin et al., 2001; Faucette et al., 2007). We detected significant up-regulation of pPBREM-SV40-luc activity in HepG2 cells cotransfected with both CAR and PXR expression plasmids (p < 0.001; Fig. 4). Moreover, the activation of the construct was further significantly augmented in cells coexpressing CAR and exposed to VPA (5.8-fold increase, p < 0.001; Fig. 4). Similarly, VPA increased significantly activation of pPBREM-SV40-luc in PXR-expressing cells (2-fold, p < 0.05). Under the same experimental conditions, CITCO (1 μM) increased activation of the plasmid 2.1-fold in the cells expressing CAR and rifampicin (10 μM) 2.4-fold in PXR-transfected cells. Nevertheless, we also detected comparable activation of the luciferase construct by VPA in cells with no exogenous PXR (p < 0.05) (Fig. 4), which documents that exogenous PXR has minor or no effect on activation of pPBREM-SV40-luc by VPA. Thus, we observed a similar pattern in activation of pPBREM-SV40-luc and pMDR1E-SV40-luc by VPA (Figs. 3B and 4). In both cases, we demonstrated that VPA increases CAR-mediated, but not PXR-mediated transactivation of the constructs.
Analysis of VPA-Mediated Up-Regulation of CYP3A4 and MDR1 mRNAs in LS174T Cells. To evaluate the ability of VPA to induce CYP3A4 and MDR1 mRNA levels by means of CAR and PXR activation, real-time RT-PCR was used. CYP3A4 and MDR1 mRNAs were quantified in samples of LS174T cells transfected with expression plasmids encoding studied nuclear receptors and exposed to VPA for 48 h. Significant effect of VPA on CYP3A4 mRNA level was observed in cells transfected with both CAR and PXR nuclear receptors yielding a 4.2-fold and 3.8-fold (p < 0.001 and p < 0.01) induction, respectively (Fig. 5A). VPA also significantly up-regulated CYP3A4 mRNA in LS174T cells, which were not cotransfected with any nuclear receptor (p < 0.01) (Fig. 5A). This might be in agreement with high expression of endogenous PXR in LS174T cells, which could at least partly participate in up-regulation of CYP3A4 mRNA by VPA (data in Fig. 1) (Burk et al., 2005b). In the case of MDR1, we observed a similar profile of VPA-mediated MDR1 mRNA up-regulation. Contrary to transfection assay results, we detected a statistically significant effect of VPA on MDR1 mRNA expression in PXR-transfected LS174T cells (2.0-fold increase, p < 0.05). However, the MDR1 mRNA level was not statistically significantly different from the level in VPA-treated cells (Fig. 5B). Thus, we suppose that the increase in MDR1 mRNA level after treatment with VPA is caused mostly by the mechanism of HDAC inhibition described previously rather than through activation of the PXR pathway by VPA (Morrow and Nakagawa, 1994; Jin and Scotto, 1998; Xiao and Huang, 2005). It is noteworthy that we observed statistically significant (p < 0.05) up-regulation of MDR1 mRNA after cotransfection of LS174T cells with CAR expression vector and treatment with VPA (Fig. 5B). On the other hand, cotransfection of LS174T cells with expression plasmid for either PXR or CAR without exposure to VPA did not result in statistically significant up-regulation of CYP3A4 and MDR1 mRNAs (Fig. 5, A and B). Under the same experimental conditions, treatment of LS174T cells with rifampicin (10 μM) resulted in a 12-fold increase in CYP3A4 mRNA and a 7-fold increase in MDR1 mRNA.

Effect of VPA on CAR- and PXR-mediated activation of reporter constructs of MDR1 gene. HepG2 cells were transiently transfected with pCR3-CAR or pSG5-PXR expression plasmid (50 ng) and appropriate reporter construct (0.4 μg/well) using Lipofectamine2000 transfection reagent according to the manufacturer's instructions. Transfected HepG2 cells were maintained in medium containing the tested compounds at the indicated concentrations for 24 h. A, effect of VPA on CAR- and PXR-mediated transactivation of pMDR1-luc construct containing the enhancer region of MDR1 gene (–7975/–7013) and the basal promoter from –1803 to +231. B, VPA-mediated transcriptional activation of pMDR1E-SV40-luc construct containing the enhancer region of MDR1 gene and viral promoter SV40 in cells coexpressing exogenous CAR or PXR. C, influence of androstenol (Andol), an inverse agonist of CAR, on CAR-mediated transcriptional activation of pMDR1E-SV40-luc in HepG2 cells treated with VPA. HepG2 cells were transiently transfected with pCR3-CAR (40 ng) and pMDR1E-SV40-luc construct (0.4 μg/well) using Lipofectamine2000. Control and VPA-treated cells were exposed to DMSO (0.1%) to eliminate the influence of the androstenol solvent. Data are presented as -fold activation of nontreated cells transfected only with the appropriate reporter construct, pMDR1E-luc or pMDR1E-SV40-luc. All means ± S.D. were calculated from quadruplicates of a representative experiment. ###, p < 0.001: statistically different from nontreated cells transfected only with reporter construct. *, p < 0.05; **, p < 0.01; ***, p < 0.001: statistically different from VPA-nontreated cells cotransfected with either pCR3-CAR or pSG5-PXR expression vectors. ≠≠, p < 0.01: statistically different from cells transfected with reporter plasmid and exposed to VPA.
Determination of CYP3A4 Catalytic Activity in LS174T Cells Exposed to VPA (600 μM). CYP3A4 induction was subsequently investigated employing functional assay based on determination of specific CYP3A4-mediated testosterone 6β-hydroxylation activity measured in LS174T cell lysate. As shown in Fig. 6, only cells transfected with plasmid encoding PXR yielded statistically significant increase in CYP3A4 catalytic activity after 48 h exposure to VPA at a concentration of 600 μM (p < 0.05).

Effect of VPA on CAR- and PXR-mediated activation of the PBREM region of the CYP2B6 promoter. HepG2 cells were transiently transfected with pCR3-CAR or pSG5-PXR expression plasmid (50 ng) and pPBREM-SV40-luc reporter construct containing two DR4-type motifs of CYP2B6 PBREM (0.4 μg/well) using Lipofectamine2000 transfection reagent according to the manufacturer's instructions. Transfected HepG2 cells were maintained in medium containing VPA (500 μM) for 24 h. ###, p < 0.001: statistically different from nontreated cells transfected only with reporter construct; *, p < 0.05; ***, p < 0.001: statistically different from VPA-nontreated cells cotransfected with either pCR3-CAR or pSG5-PXR expression vectors.
VPA Induces CYP3A4 mRNA in Primary Human Hepatocytes and Synergizes with the Effect of Rifampicin. To determine whether VPA induces CYP3A4 mRNA and whether it influences the induction by rifampicin, primary cultures of human hepatocytes were treated individually or in combination with VPA (500 μM) and/or rifampicin (10 μM). CYP3A4 mRNA was significantly up-regulated in the hepatocytes exposed to VPA for 48 h taken from both donor 1 (14.1-fold, p < 0.01) (Fig. 7A) and donor 2 (3.8-fold, p < 0.01) (Fig. 7B). As shown in Fig. 7C, by using the unpaired Student's t test, we revealed a statistically significant increase of CYP3A4 mRNA compared with control treated with DMSO (0.1%) in cell samples treated with rifampicin (10 μM) for 24 h (donor 1, 20.1-fold and donor 2, 13.6-fold, respectively; p < 0.001). In cells cultivated in medium containing VPA (500 μM) and vehicle (DMSO, 0.1%) used for elimination of rifampicin solvent effect, we also detected significant induction of CYP3A4 mRNA relative to control (DMSO, 0.1%) (p < 0.01 and p < 0.05, respectively). Furthermore, when VPA (500 μM) and rifampicin (10 μM) were added simultaneously into medium and hepatocytes were cultivated for 24 h, up-regulation of CYP3A4 was synergistic rather than additive (62- and 34-fold). Using two-way ANOVA, this synergy was found to be statistically significant at the level of p < 0.01 (Fig. 7C).

Analysis of VPA-mediated up-regulation of CYP3A4 and MDR1 mRNAs. LS174T cells were transfected with pCR3-CAR or pSG5-PXR expression plasmids (400 ng/well) using Lipofectamine2000 transfection reagent according to the manufacturer's instructions and exposed to VPA (500 μM) for 48 h. mRNA expression of tested genes was determined using real-time RT-PCR and normalized to HPRT housekeeping gene. The effect of VPA (500 μM) on CYP3A4 (A) and MDR1 (B) mRNA expression is presented as -fold increase to control nontreated cells. Data are the means ± S.D. of three individual cell samples. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Examination of Interactions between CAR/RXRα Heterodimer or CAR Monomer and Responsive Elements of CYP3A4 and MDR1 Promoters in the Presence of VPA using a Ligand-Dependent EMSA. The interactions between CAR/RXRα heterodimers and nuclear receptor-binding motifs of CYP3A4 (DR3, ER6) and MDR1 enhancer (DR4) in the presence of VPA were examined via EMSA using in vitro translated CAR and RXRα proteins. CAR forms a complex with RXRα, which binds DR3, DR4, and ER6 responsive elements of CYP3A4 and MDR1 even in the absence of a ligand (Goodwin et al., 1999, 2002; Geick et al., 2001; Burk et al., 2005a). This complex formation can be enhanced by ligand binding to CAR, which can be revealed using EMSA (Frank et al., 2004). We detected that VPA at the tested concentrations of 500 and 1000 μM increased the formation of CAR/RXRα complex with the DR3 responsive element of CYP3A4 promoter (Fig. 8, A, lane 8; and B, lanes 6 and 7). Figure 8A shows that using recombinant CAR and RXRα proteins, VPA moderately (by ∼30%) augmented the CAR/RXRα/DR3 complex in EMSA experiments (Fig. 8A, lane 8). In the next experiments, we used nuclear fraction from HepG2 cells transfected with expression vector encoding RXRα. VPA at the concentration of 500 μM augmented the complex 1.8-fold and at the concentration of 1000 μM increased 4.2-fold the binding of CAR/RXRα to DR3 (Fig. 8B, lanes 6 and 7). Similarly, as shown in Fig. 8D (lane 6), we detected increased (∼1.5-fold) binding of CAR/RXRα heterodimer to oligonucleotide containing the DR4 motif of MDR1 enhancer in the presence of VPA (1000 μM). Consistently, CITCO, an agonist of human CAR, augmented binding of CAR/RXRα complex to DR3 response elements (Fig. 8, A, lane 5; and B, lane 3) (Maglich et al., 2003). The inverse agonist of CAR, androstenol (10 μM), decreased formation of CAR/RXRα complex with response element DR3 in samples treated with CITCO (500 nM) or VPA (500 and 1000 μM) (Fig. 8, A–D). Interestingly, we observed that VPA at tested concentrations augments interaction of recombinant CAR monomer with the DR3 motif of CYP3A4 promoter (Fig. 8C, lane 2). On the other hand, we did not detect any significant and reproducible effect of VPA on interaction of CAR/RXRα heterodimer with the ER6 response element of the CYP3A4 gene (data not shown). RXRα alone did not reveal any interaction with tested response elements (data not shown). Similarly, we did not detect any complex after incubation of nuclear fraction of HepG2 cells and recombinant RXRα with DR3 oligonucleotide under the conditions used (data not shown). These data suggest that VPA could directly interact with CAR/RXRα heterodimer and CAR monomer in binding with DR3 and DR4 response elements of CYP3A4 and MDR1.
Unfortunately, there is no report in the literature that introduces reliable ligand-dependent EMSA assay with recombinant PXR at present. Therefore, we could not analyze the interaction of VPA with PXR/RXRα complex at present, using the EMSA method.

Determination of CYP3A4 catalytic activity. LS174T cells were transfected with expression plasmids encoding CAR or PXR. Subsequently, cells were treated with VPA (600 μM) and specific CYP3A4-mediated testosterone 6β-hydroxylation activity was detected after 48 h. The concentration of the 6β-testosterone metabolite was determined using the HPLC system in cell lysate. Final activity was normalized to sample protein concentration. Bars indicate the means ± S.D. calculated from three samples. *, p < 0.05.
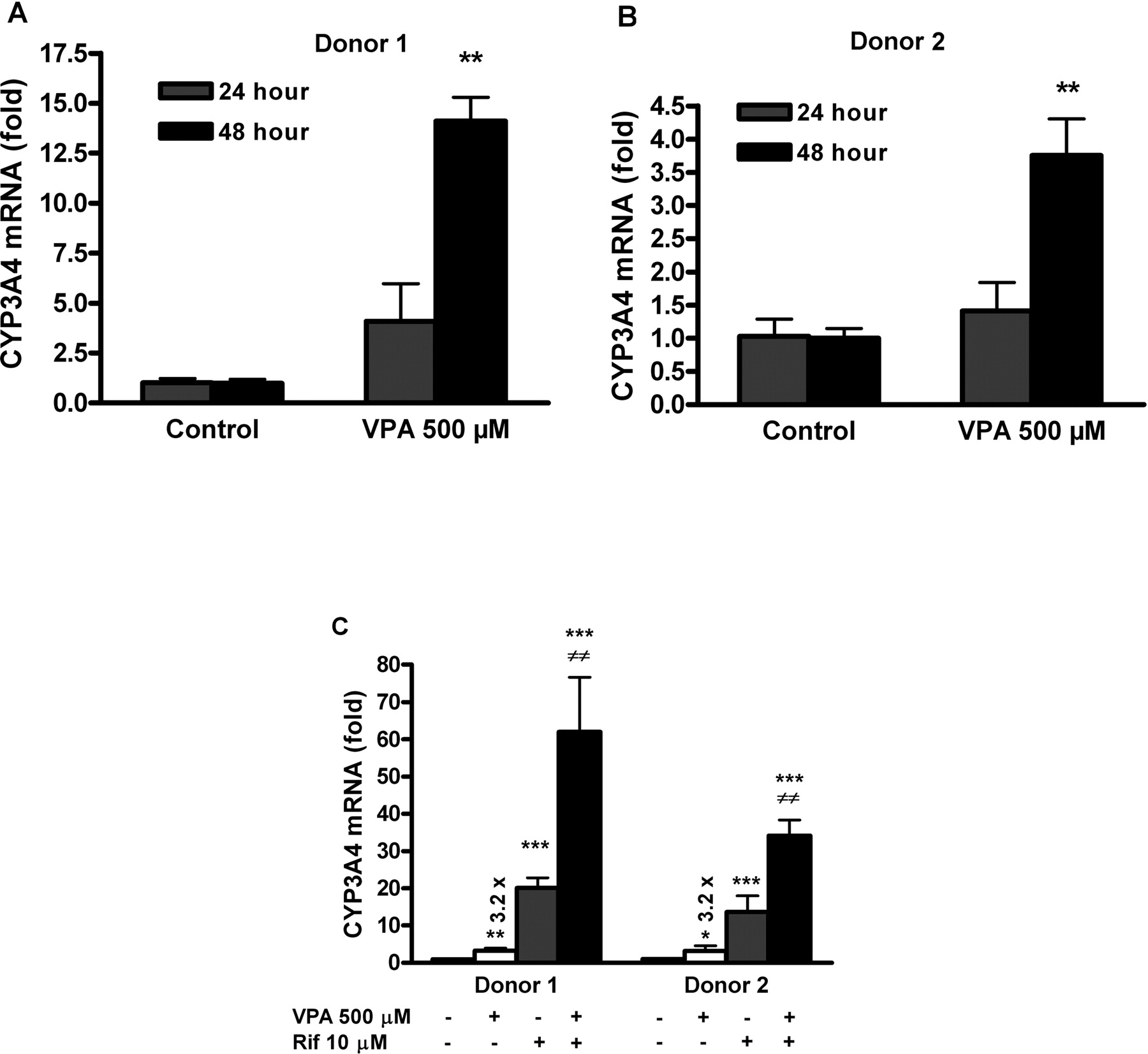
VPA synergizes with rifampicin (Rif) in induction of CYP3A4 mRNA in primary human hepatocytes. Primary human hepatocyte cultures were cultivated together with VPA and/or rifampicin either individually or in combination, and then assayed using real-time RT-PCR. CYP3A4 mRNA expression is normalized to HPRT housekeeping gene and presented as -fold increase to control nontreated cells. Values represent the means ± S.D. of three independent wells. A and B, 48-h exposure of primary hepatocytes to VPA (500 μM) up-regulates CYP3A4 mRNA (p < 0.01). This effect of VPA was analyzed using unpaired Student's t test. C, rifampicin, VPA or a combination of these two drugs significantly up-regulates CYP3A4 mRNA to control (0.1% DMSO-treated cells) with p values of * p < 0.05; **, p < 0.01; and ***, p < 0.001 (unpaired Student's t test) after 24 h of treatment. When given in combination, the induction is in both cases statistically synergistic rather than additive, at ≠≠, p < 0.01 (analyzed by two-way ANOVA with interaction).
Discussion
VPA, an effective broad-spectrum anticonvulsant, has been shown to affect expression of a large number of genes (Bosetti et al., 2005). Some of these recognized alterations in gene expression have been attributed to HDAC inhibitory activity of VPA (Chen et al., 1999; Phiel et al., 2001; Werling et al., 2001; Eyal et al., 2006). In this study, we show for the first time (to our knowledge) that VPA is capable, in a clinically relevant range of concentrations <1000 μM (Davis et al., 1994; Wen et al., 2001; Centorrino et al., 2003; Allen et al., 2006), of up-regulating CYP3A4 and MDR1 gene expression also, by a different molecular mechanism, via direct activation of the CAR pathway. Moreover, we demonstrate that VPA can increase CYP3A4 gene expression and activity through activation of PXR nuclear receptor as well.
Ligand-activated nuclear receptors CAR and PXR up-regulate expression of target genes at the transcriptional level through interaction with specific promoter response elements. Therefore, to examine the potential effect of VPA on CAR- and PXR-mediated transcriptional regulation of CYP3A4 and MDR1 genes, we first used transient transfection experiments with luciferase reporter constructs containing a relevant regulatory promoter sequence of tested genes (Goodwin et al., 1999, 2001, 2002; Burk et al., 2005a). Using transient transfection, we established that the reporter constructs appeared in cells separately from the chromosomal DNA and nucleosome structure, which guaranteed that the detected effect of VPA on activation of luciferase constructs is independent of chromatin remodeling caused by VPA-mediated HDAC inhibition.

The ability of CAR/RXRα heterodimer to bind CAR response elements of CYP3A4 (DR3, ER6) and MDR1 (DR4) in the presence of VPA was investigated using EMSA as described under Materials and Methods. A, the complex formation of CYP3A4 DR3 with CAR/RXRα was examined with in vitro translated wild-type CAR and RXRα proteins in the presence of solvent (water or DMSO 0.1% where indicated), the CAR agonist CITCO (6.25 μM; lanes 5 and 7), the inverse agonist of CAR, androstenol (Andol) (10 μM; lanes 6, 7, and 9), VPA (1000 μM; lanes 3, 8, and 9), or CITCO-androstenol and VPA-androstenol combinations (lanes 7 and 9). B, combined gel shift/supershift experiments were performed using in vitro translated wild-type CAR protein and the nuclear fraction of HepG2 cells transfected with RXRα expression vector. The concentration-dependent effect of VPA (100, 500, or 1000 μM; lanes 5–7) on CAR/RXRα complex was studied. The band supershift was performed with anti-RXRα antibody (1 μg; lane 8). C, effect of VPA on CAR monomer interaction with CYP3A4 DR3. The experiments were performed using in vitro translated CAR without RXRα and with VPA (1000 μM) and androstenol (10 μM) as an inverse agonist of CAR. D, the capability of VPA (1000 μM; lane 6) to affect CAR/RXRα heterodimer interaction with MDR1 DR4 response element was tested using in vitro translated wild-type CAR and RXRα proteins. CITCO (6.25 μM) and androstenol (10 μM) were used as a known ligand and inverse agonist of CAR, respectively. Representative gels are shown. Band signal intensity was quantitated by densitometry and data are expressed as -fold increase (fold) relative to their vehicle controls (DMSO or water).
We demonstrate that VPA induces CAR- and PXR-mediated transactivation of p3A4-luc construct containing both DR3 and ER6 responsive elements of CYP3A4 required for CAR- and PXR-mediated regulation of the gene (Goodwin et al., 1999) (Fig. 1). Interestingly, we found out that the combination of VPA with prototypic ligands causes far more pronounced transactivation of the p3A4-luc construct (Fig. 2). These data indicate a synergistic effect of VPA and rifampicin on transactivation of CYP3A4 via PXR, which could play an important role during cotreatment with these drugs.
In additional experiments, we examined activation of two different luciferase reporter constructs containing MDR1 enhancer and either native or viral SV40 basal promoter (Burk et al., 2005a). We did not detect any significant CAR- or PXR-dependent effect of VPA on activation of MDR1 gene promoter in HepG2 cells transfected with pMDR1-luc reporter construct, although an increase in transcriptional activation of this reporter construct was observed in cells exposed to VPA (Fig. 3A). On the other hand, using pMDR1E-SV40-luc reporter construct containing the MDR1 enhancer upstream of the SV40 viral promoter, VPA was identified as an activator of the MDR1 enhancer via the CAR pathway (Fig. 3B). Thus, by using pMDR1E-SV40-luc construct, we eliminated the potential interference of additional transcriptional factors that bind relevant sites in the basal promoter of MDR1 gene such as NF-Y, Sp1, AP-1, NK-κB, C/EBPβ, and so on (Scotto, 2003), and analyzed specific CAR-/PXR-mediated activation of the MDR1 enhancer by VPA. Moreover, the reporter plasmid lacks the inverted CCAAT motif (Y-box), which was shown to be essential for activation of MDR1 promoter in transient transfection reporter assays by several other HDAC inhibitors (Jin and Scotto, 1998). Despite this fact, significant activation of pMDR1E-SV40-luc reporter construct by VPA was observed also in the absence of cotransfected CAR or PXR, which indicates that VPA can also activate MDR1 enhancer of pMDR1E-SV40-luc reporter construct (Fig. 3B). We suppose that endogenous CAR or PXR is not involved in the phenomenon because their functional expression in HepG2 cells is very low. However, we cannot exclude potential up-regulation of various transcriptional factors by VPA treatment, which could lead to transactivation of the reporter construct.
Subsequently, interaction of VPA with PXR transactivation pathway has been studied in more detail using another gene reporter construct, pPBREM-SV40-luc, containing two DR4-type motifs of the CYP2B6 PBREM region upstream of SV40 viral promoter, which were recently demonstrated to have a high affinity to both PXR and CAR (Faucette et al., 2007). These experiments have revealed significant CAR-mediated, but no PXR-mediated, activation of pPBREM-SV40-luc reporter construct by VPA, which is in agreement with the findings obtained using pMDR1-SV40-luc reporter construct. Thus, we suggest a different effect of VPA on CAR- and PXR-mediated transcriptional activation of heterologous reporter constructs pPBREM-SV40-luc and pMDR1-SV40-luc in comparison with p3A4-luc construct (Figs. 1, 3B, and 4). We hypothesize that this discrepancy can be explained considering the report of Masuyama et al. (2005), who demonstrated a ligand- and promoter-specific fashion of PXR-mediated transcription of CYP3A4 and MDR1 genes.
CAR- and PXR-mediated regulation of CYP3A4 and MDR1 gene expression was subsequently analyzed by real-time RT-PCR in samples of LS174T cells (Fig. 5, A and B). mRNA expression of the studied genes was normalized to both HPRT and B2M housekeeping genes to eliminate a false interpretation potentially caused by non-specific VPA-mediated up-/down-regulation of these housekeeping genes. We observed the effect of coexpressed CAR and PXR on VPA-mediated up-regulation of CYP3A4 mRNA in LS174T cells (Fig. 5A), which well correlates with the reporter experiments. In the case of MDR1, the mRNA level was significantly up-regulated in LS174T cells transfected with CAR expression vector, whereas the VPA-mediated increase of MDR1 mRNA in PXR-expressing cells was not statistically different from that of nontransfected cells exposed to VPA (Fig. 5B). Considering data from gene reporter assay with pMDR1E-SV40-luc (Fig. 3A) and pPBREM-SV40-luc (Fig. 4), we assume that the observed influence of VPA on MDR1 mRNA expression in LS174T cells transfected with PXR should instead be attributed to HDAC inhibitory activity of VPA, which was previously demonstrated to cause MDR1 gene up-regulation (Eyal et al., 2006). Thus, we hypothesize that the total induction of MDR1 mRNA expression in LS174T cells transfected with CAR and PXR might be a sum of several VPA effects such as inhibition of HDAC (Eyal et al., 2006) and interaction of VPA with CAR or PXR. On the other hand, we suppose that the effect of VPA on CYP3A4 mRNA up-regulation is mediated by a transcription factor(s), since HDAC inhibition did not result in induction of human CYP3A4 or its rat homolog Cyp3a2 mRNA (Rodríguez-Antona et al., 2003; Eyal et al., 2006). Nevertheless, we hypothesize that HDAC inhibitory activity of VPA synergistically augments the effect of rifampicin in PXR-mediated up-regulation of CYP3A4 mRNA in primary human hepatocytes (Fig. 7C).
The interaction of VPA with CAR was investigated in more detail using EMSA, in which interaction of VPA with CAR/RXRα heterodimer or CAR monomer (Frank et al., 2003) and responsive elements of both CYP3A4 and MDR1 promoters were examined. In the presence of VPA, we detected augmented binding of CAR/RXRα heterodimer to the DR3 responsive element of CYP3A4 and to the DR4 responsive element of MDR1 (Fig. 7, A–D). Binding of CAR/RXRα to the ER6 element of CYP3A4 was not affected by VPA (data not shown), which might correlate with low affinity of CAR/RXRα complex to bind the ER6 motif and with minor role of ER6 in CAR-mediated CYP3A4 gene transactivation (Goodwin et al., 2002). Interestingly, we also observed increased binding of the CAR monomer to the DR3 motif of CYP3A4 promoter in the presence of VPA (Fig. 7C, lane 2). Androstenol decreased the binding of CAR/RXRα heterodimer and CAR monomer to DR3 and CAR/RXRα heterodimer to DR4 in samples containing VPA (Fig. 7, A, C, and D), which provides us additional evidence that VPA transactivates CYP3A4 and MDR1 through CAR. Interestingly, we observed a difference in the VPA-activated CAR/RXRα heterodimer binding to the DR3 responsive element in EMSA experiments performed using recombinant CAR and RXRα proteins (Fig. 7A, lane 8) in comparison with the experiments in which recombinant RXRα was replaced with nuclear extract from HepG2 cells transfected with expression vector encoding RXRα (Fig. 7B, lane 7). Binding of CAR/RXRα complex with recombinant in vitro translated RXRα to the DR3 motif was only slightly augmented in the presence of VPA (∼30%); however, using a nuclear fraction of HepG2 cells transfected with RXRα, we observed a more than 4-fold effect of VPA on the CAR/RXRα/DR3 complex formation (Fig. 7B, lane 7). Based on this finding, we hypothesize that the nuclear fraction of HepG2 cells could contain another transcription cofactor involved in regulation of CAR/RXRα binding to DR3 in the presence of VPA.
Finally, we analyzed CYP3A4 enzymatic activity in LS174T cells exposed to VPA (600 μM) for 48 h. As shown in Fig. 6, significant increase of CYP3A4 activity was detected only in VPA-treated LS174T cells transfected with PXR expression vector (p < 0.05). We did not detect any alteration of CYP3A4 enzymatic activity in non-transfected LS174T cells, although LS174T cells express PXR (our unpublished data; Burk et al., 2005b). With respect to this fact, we suppose that CYP3A4 catalytic activity can be increased only in cells rich in PXR protein.
In conclusion, we show that VPA mediates transactivation of CYP3A4 and MDR1 genes via direct interaction with CAR nuclear receptor. Furthermore, we present that VPA up-regulates CYP3A4 and its catalytic activity through PXR. With respect to the published data on the effect of VPA in regulation of MDR1 gene expression, we suggest that interaction of VPA with the CAR signaling pathway together with VPA-mediated HDAC inhibition is the mechanism involved in up-regulation MDR1 gene. Because MDR1 plays an important role in the multidrug resistance (MDR) phenomenon, we assume that administration of VPA could lead to an increase in tumor resistance against many anticancer drugs, which are transported by P-glycoprotein. Finally, our data indicate that VPA in clinically relevant concentrations during treatment of schizoaffective disorders, acute mania, and refractory epilepsy, when VPA plasma concentrations range within 500 to 1000 μM, could cause drug-drug interactions with coadministered drugs that are metabolized by CYP3A4.
Footnotes
-
This work was supported in part by grants from The Grant Agency of the Charles University in Prague (94/2005/C), the Czech Science Foundation (170/53/75301), and the Ministry of Education of the Czech Republic (MSM 6198959216).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.014456.
-
ABBREVIATIONS: VPA, valproic acid; HDAC, histone deacetylase; CITCO, (6-(4-chlorophenyl) imidazo [2,1-b][1,3] thiazole-5-carbaldehyde O-(3,4-dichlorobenzyl) oxime); NR, nuclear receptor; RT-PCR, reverse transcriptase polymerase chain reaction; PXR, pregnane X receptor; CAR, constitutive androstane receptor; RXRα, retinoid X receptor α (9-cis retinoic acid receptor-α); FCS, fetal calf serum; DMSO, dimethyl sulfoxide; DR, direct repeat; ER, everted repeat; HPRT, hypoxanthine-guanine phosphoribosyl transferase; B2M, β2-microglobulin; PBREM, phenobarbital responsive enhancer module; EMSA, electrophoretic mobility shift assay; ANOVA, analysis of variance.
- Received December 20, 2006.
- Accepted March 23, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}