


Abstract
The hormonal regulation of human CYP2C18 and CYP2C19, which are expressed in a male-specific manner in liver and kidney in a mouse transgenic model, was examined. The influence of prepubertal castration in male mice and testosterone treatment of female mice was investigated, as was the effect of continuous administration of growth hormone (GH) to transgenic males. Prepubertal castration of transgenic male mice suppressed the expression of CYP2C18 and CYP2C19 in liver and kidney to female levels, whereas expression was increased for the endogenous female-specific mouse hepatic genes Cyp2c37, Cyp2c38, Cyp2c39, and Cyp2c40. Testosterone treatment of female mice increased CYP2C18 and CYP2C19 expression in kidney, and to a lesser extent in liver, but was without effect in brain or small intestine, where gene expression was not gender-dependent. Continuous GH treatment of transgenic males for 7 days suppressed hepatic expression of CYP2C19 (>90% decrease) and CYP2C18 (∼50% decrease) but had minimal effect on the expression of these genes in kidney, brain, or small intestine. Under these conditions, continuous GH induced all four female-specific mouse liver Cyp2c genes in males to normal female levels. These studies indicate that the human CYP2C18 and CYP2C19 genes contain regulatory elements that respond to the endogenous mouse hormonal profiles, with androgen being the primary regulator of male-specific expression in kidney, whereas the androgen-dependent pituitary GH secretory pattern is the primary regulator of male-specific expression in liver in a manner that is similar to the regulation of the endogenous gender-specific hepatic genes.
Hepatic cytochrome P450 (P450)-dependent metabolism of steroids and xenobiotics is sexually dimorphic in rodents and some other species (Waxman and O'Connor, 2006), whereas in humans gender differences are much less pronounced. Metabolism by gender-specific P450 enzymes could lead to gender-dependent susceptibility to chemical toxicants and carcinogenicity (Aldridge et al., 2003). Some reports have suggested gender differences in activity of CYP2C19 (Xie et al., 1997) and CYP2E1 (Kim and O'Shea, 1995), but such differences have been difficult to reproduce by others (Kim and O'Shea, 1995; Hägg et al., 2001; Bebia et al., 2004). CYP3A4, the most important drug-metabolizing P450 enzyme in human liver, is more highly expressed in women than in men, as revealed by in vivo measurements of the metabolism of probe drug and by examination of CYP3A4 protein and RNA levels in a large collection of human livers (Wolbold et al., 2003; Diczfalusy et al., 2008). The regulation of the gender-dependent expression of CYP3A4 has been studied in a transgenic CYP3A4/CYP3A7 humanized mouse line, where CYP3A4 mRNA and protein are expressed in livers of adult female mice but not male mice, and where continuous infusion of recombinant growth hormone (GH) in transgenic male mice increased hepatic CYP3A4 mRNA and protein to female levels (Cheung et al., 2006).
The sexually dimorphic, ultradian rhythm of circulating GH levels has been shown to regulate the gender-dependent expression of hepatic monooxygenases in rats and mice (Waxman and O'Connor, 2006). In males, the pulsatile GH-secretion pattern activates and stimulates nuclear translocation of signal transducer and activator of transcription (STAT5b) (Waxman et al., 1995), which is required for gender-specific expression of >1000 genes in mouse liver (Clodfelter et al., 2006). The gender-dependent differences in plasma GH profiles first emerge at puberty but are set, and ultimately regulated, by gonadal steroid imprinting during the neonatal period (Jansson et al., 1985, Jansson et al., 1985). Sex differences in plasma GH profiles are most dramatic in rodents, but significant male-female differences in the regulation of pituitary GH release also exist in humans (Veldhuis et al., 2001).
In addition to GH, gonadal hormones are essential for the differentiation of many sexually dimorphic P450s. Whereas estrogens induce the female-specific gene CYP2C12 in female rat liver (Dannan et al., 1986), androgens are required for expression of the male-specific genes CYP2A2 (Waxman et al., 1988), CYP2C11 (Morgan et al., 1985; Waxman et al., 1985), CYP2C13 (McClellan-Green et al., 1989), CYP3A2 (Waxman et al., 1985; Ribeiro and Lechner, 1992), and CYP4A2 (Sundseth and Waxman, 1992) in male rat liver. Androgens and estrogens act on the hypothalamic-pituitary axis, which controls the sexually dimorphic pattern of GH secretion, and thereby influence the expression of the gender-dependent hepatic P450 enzymes in an indirect manner (Mode and Norstedt, 1982; Jansson et al., 1985, Jansson et al., 1985). In contrast, androgens act directly on the kidney to regulate gender-dependent P450 expression (Henderson and Wolf, 1991; Imaoka et al., 1992; Sundseth and Waxman, 1992).
Differences in diets, drinking, and smoking habits, and medication history are all known to affect P450 enzyme expression and/or activity in humans (Bomsien et al., 2006; Thum et al., 2006). These environmental factors can be controlled in animals maintained under defined dietary and environmental conditions, suggesting that animal models, including transgenic mice, may be useful for investigation of the gender-specific expression and hormonal regulation of human P450 enzymes. Thus, recently we found strong gender differences in human P450 gene and enzyme expression in a humanized CYP2C18/CYP2C19 transgenic mouse line, where adult male mice expressed much higher CYP2C18 and CYP2C19 mRNA levels in liver and kidney compared with that in female mice (Löfgren et al., 2008).
We considered it of interest to investigate to what extent GH and androgens are responsible for the sexually dimorphic expression of the human genes CYP2C18 and CYP2C19. We examined the influence of castration and continuous GH treatment of male mice and continuous testosterone treatment of female mice in the transgenic mouse model. Our findings show that exogenous GH administration and prepubertal castration suppress CYP2C18 and CYP2C19 expression in liver and kidney of transgenic male mice and stimulate hepatic expression of four endogenous female specific Cyp2c genes. Testosterone administration to female mice stimulates the expression of CYP2C18 and CYP2C19 in both liver and kidney. These results provide insight into the sexually dimorphic regulation of CYP2C18 and CYP2C19 expression, suggesting that this animal model can be a valuable tool for studying the regulation of these human CYP genes.
Materials and Methods
Chemicals, Enzymes, and Other Reagents. Alzet micro-osmotic pumps, model 1007D, were obtained from Scanbur AB (Sollentuna, Sweden). Recombinant rat GH was obtained from the National Hormone and Peptide Program (Torrance, CA). Oligonucleotide polymerase chain reaction (PCR) primers were purchased from Invitrogen (Paisley, UK). Testosterone 60-day release pellets and 60-day placebo control pellets were obtained from Innovative Research of America (Sarasota, FL). All of the other laboratory chemicals were of analytical grade and obtained from commercial suppliers.
Animals and Treatments.CYP2C18/CYP2C19 hemizygous transgenic mice (Löfgren et al., 2008) and wild-type C57BL/6OlaHsd littermates were housed at the Karolinska Institute in Stockholm and were treated according to research protocols approved by the Swedish Ethical Application Committee. The mice were housed on wood shavings in plastic cages, with water and commercial mouse diet supplied ad libitum.
In the first study, transgenic male mice were castrated at ∼3 weeks before puberty, which occurs between 6 and 9 weeks of age. Transgenic male littermates underwent sham surgery at the same time. Transgenic female littermates had a 15-mg testosterone pellet (60-day release) or a placebo pellet (60-day release) inserted subcutaneously in the skin of the neck at 3 weeks of age. All of the animals were necropsied approximately 8 weeks after surgery, at 11 weeks of age. In addition, untreated wild-type littermates of both genders were euthanized at the same age.
In the second study, male transgenic mice received continuous infusion with recombinant rat GH. An Alzet 1007D micro-osmotic pump designed to deliver the solution at a rate of 0.5 μl/h was implanted subcutaneously at the back of the neck. Adult (8–11 weeks old) transgenic male mice received the osmotic infusion of recombinant rat GH (660 ng/h) dissolved in buffer (91.5%, 30 mM NaHCO3/0.15 M NaCl, pH 10.3; 8.5%, 0.5 M NaHCO3; and rat albumin at a final concentration of 100 μg/ml) or infusions with buffer only. Transgenic female mice and placebo-treated males were used as controls. All the transgenic male mice were killed either 4 days or 7 days after osmotic pump implantation. The female transgenic controls and placebo-treated controls were killed at the same time. For both studies at least four animals were included in each group. Three- and 9-week-old mice used for preliminary measurements of S-mephenytoin 4′-hydroxylation were either reared at the facilities of AstraZeneca (Södertälje, Sweden) or purchased as wild-type C57BL/6 mice (therefore, not genotyped) from B&K Universal Limited (Grimston, East Yorkshire, UK).
PCR Genotyping. Mice were genotyped to confirm the inserted human CYP2C18/CYP2C19 gene segment. Genotyping was performed on genomic DNA (gDNA) extracted from tissues obtained from either a small tail section or a 2- to 3-mm diameter ear punch using a commercially available kit (DNeasy Tissue kit; QIAGEN, Hilden, Germany). Amplification of interleukin 2 (internal control) and CYP2C19 in the extracted gDNA was performed in a 25-μl reaction volume containing 2.5 μl of 10× HiFi PCR MasterMix (ABgene House, Surrey, UK), 400 nM each primer, and 1 μl of gDNA. The mice used for S-mephenytoin hydroxylation measurements were genotyped to confirm the presence of both the CYP2C18/CYP2C19 transgene and the bacterial artificial chromosome 3′ and bacterial artificial chromosome 5′ ends of the inserted transgene. Sequences of the gene-specific oligonucleotide PCR primers are shown in Table 1. The PCR thermoprofile consisted of an initial denaturation for 2 min at 94°C followed by 33 cycles of 94°C for 10 s, 55°C for 30 s, and 72°C for 40 s, followed by a 7-min extension at 72°C. The amplified products were visualized by ethidium bromide/agarose gel electrophoresis.
Primer sequences used for real-time PCR and genotyping
Analysis ofS-Mephenytoin Metabolism. Microsomes from 3- and 9-week-old, wild-type and transgenic mice were prepared from individual male and female mouse livers as described previously (Löfgren et al., 2008) but were ultimately pooled by volume because of the limited amount of microsomal proteins obtained. Metabolism of S-mephenytoin to its major CYP2C19-dependent metabolite 4′-hydroxymephenytoin was measured as described previously (Löfgren et al., 2008) with the exception that a final S-mephenytoin solvent concentration of 1% methanol was used in each microsomal incubation. Michaelis-Menten kinetics was assumed, and apparent Km and Vmax values were estimated for all the groups using Lineweaver-Burke plots. S-Mephenytoin concentrations ranged from 10 to 250 μM, and time curves ranged from 0 to 20 min. The intrinsic clearance (CLint) was calculated as the Vmax divided by the Km.
Total RNA Isolation and cDNA Synthesis for Real-Time PCR. Mice were killed by cervical dislocation at 9 to 12 weeks of age. Portions of liver, kidney, brain, or small intestine were placed in RNAlater (QIAGEN) according to the manufacturer's recommendations. Total RNA was extracted from 40 to 100 mg of tissue using a commercially available kit (Rneasy; QIAGEN), and DNA was removed by DNase digestion (QIAGEN). The concentration of the extracted RNA was determined by absorption at 260 nm, and the RNA was then reverse-transcribed into first-strand cDNA using 0.5 μg of total RNA, 4 μl of 5× reaction buffer, 5 μM oligo(dT)18, 0.5 mM dNTPs, 10 mM DTT, 1 μl of RNaseOut (Invitrogen, Carlsbad, CA), and 200 U of Superscript II RNase reverse transcriptase (Invitrogen) in a 20-μl reaction volume. Reactions were incubated at 42°C for 60 min followed by inactivation at 70°C for 10 min. Reactions were also run in the absence of reverse transcriptase to address the possibility of gDNA contamination.
Primer Design for Real-Time PCR. The primer pairs used for the amplification of the endogenous murine Cyp2c29, Cyp2c37, Cyp2c38, and Cyp2c40 transcripts were targeted to sequences showing relatively low homology among both murine Cyp2c isoforms and the CYP2C18 and CYP2C19 transgene using a multiple sequence alignment (ClustalW). Multiple potential primers for real-time PCR were evaluated with the following criteria: observation of a single melting curve peak, visualization of a single amplicon of the appropriate length after agarose gel electrophoresis, direct sequencing of amplicons, and amplification efficiencies >95%. Primer sequences are shown in Table 1.
Real-Time Quantitative PCR. Real-time PCR reaction mixtures (25 μl) contained 12.5 μl of 2× SYBR Green Master Mix (Applied Biosystems, Foster City, CA) or 12.5 μl of TaqMan MasterMix (Applied Biosystems); cDNA (0.25 μl in the castration study or 0.125 μl in the GH study) and the appropriate primer pairs (400 nM) or 1.25 μl of TaqMan Gene Expression Assay (Applied Biosystems) specific for the mRNA studied. SYBR Green assays were performed using isoform-specific primers (see Table 1). TaqMan Gene Expression Assays with the ID numbers Mm 00656110_gH, Mm00663066_gH, Mm02602271_mH, Mm01205031_mH, and Mm01197220_mH were used for detection of Cyp2c37, Cyp2c50, Cyp2c54, Cyp2c55, and Cyp2c70, respectively. Because of the absence of detectable amplification of Cyp2c65 in any liver-derived cDNA sample, this isoform was not studied any further. Murine β-actin was quantified using a VIC-labeled TaqMan Endogenous Control Assay (Applied Biosystems).
PCR was performed using Applied Biosystems 7500 Standard Real-Time PCR system with the following PCR conditions: activation of polymerase at 95°C for 10 min, followed by 40 amplification cycles with denaturation at 95°C for 15 s, and annealing and extension at 60°C for 1 min. The specificity of the SYBR Green assays was monitored by melting curve analysis of each amplification product. The optimal combination of genes used to normalize transcript expression level was determined using qBase version 1.3.5 (Hellemans et al., 2007).
In the first study (castration/testosterone treatment), all the transcripts were analyzed in triplicate. Murine Cyp2c expression levels were normalized using the geometric mean of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and β-actin, whereas GAPDH and hypoxanthine guanine phosphoribosyltransferase (HPRT) were used for normalization of CYP2C18 and CYP2C19 transcripts. A subsequent survey of established gender-specific genes (Holloway et al., 2006) was normalized using the geometric mean of GAPDH, HPRT, and β-actin. For the GH treatment study, we chose to focus on Cyp2c18/Cyp2c19, the sexually dimorphic murine Cyp2c isoforms, and two male and female specific genes identified in the previous survey. Transcripts were analyzed in duplicate because duplicate samples were considered to be sufficient based on the variation observed in the castration study. Transcript levels were normalized using the geometric mean of GAPDH and HPRT. Relative mRNA expression levels were determined using qBase version 1.3.5 (Hellemans et al., 2007).
Statistical Analysis. All the genotype- and treatment-associated differences in mRNA expressions between treatment groups were compared with a Student's t test using Sigma Stat version 2.03 (SPSS Inc., Chicago, IL).
Results
4′-Hydroxymephenytoin Formation. Initially, a developmental study was carried out in which the ontogenecity of CYP2C19-dependent catalytic activity was examined in wild-type and transgenic CYP2C18/CYP2C19 mice. As shown in Table 2, a clear decrease in S-mephenytoin CLint occurred between 3 and 9 weeks of age in female mice and wild-type males but not in transgenic CYP2C18/CYP2C19 male mice. This decrease could be linked to an age-dependent increase in Km for both genders and genotypes and a concomitant decrease in Vmax in females only (both genotypes).
Apparent enzyme kinetics parameters of S-mephenytoin metabolism in pooled hepatic microsomes from 3- and 9-week-old wild-type and CYP2C18/CYP2C19 transgenic mice
Thus, this preliminary study indicates that the gender-dependent alterations in CYP2C18 and CYP2C19 gene expression may occur at puberty; therefore, we chose to castrate male mice and begin testosterone treatment of female mice before the onset of puberty. The hormonal regulation of CYP2C18 and CYP2C19 was then further studied in mature animals.
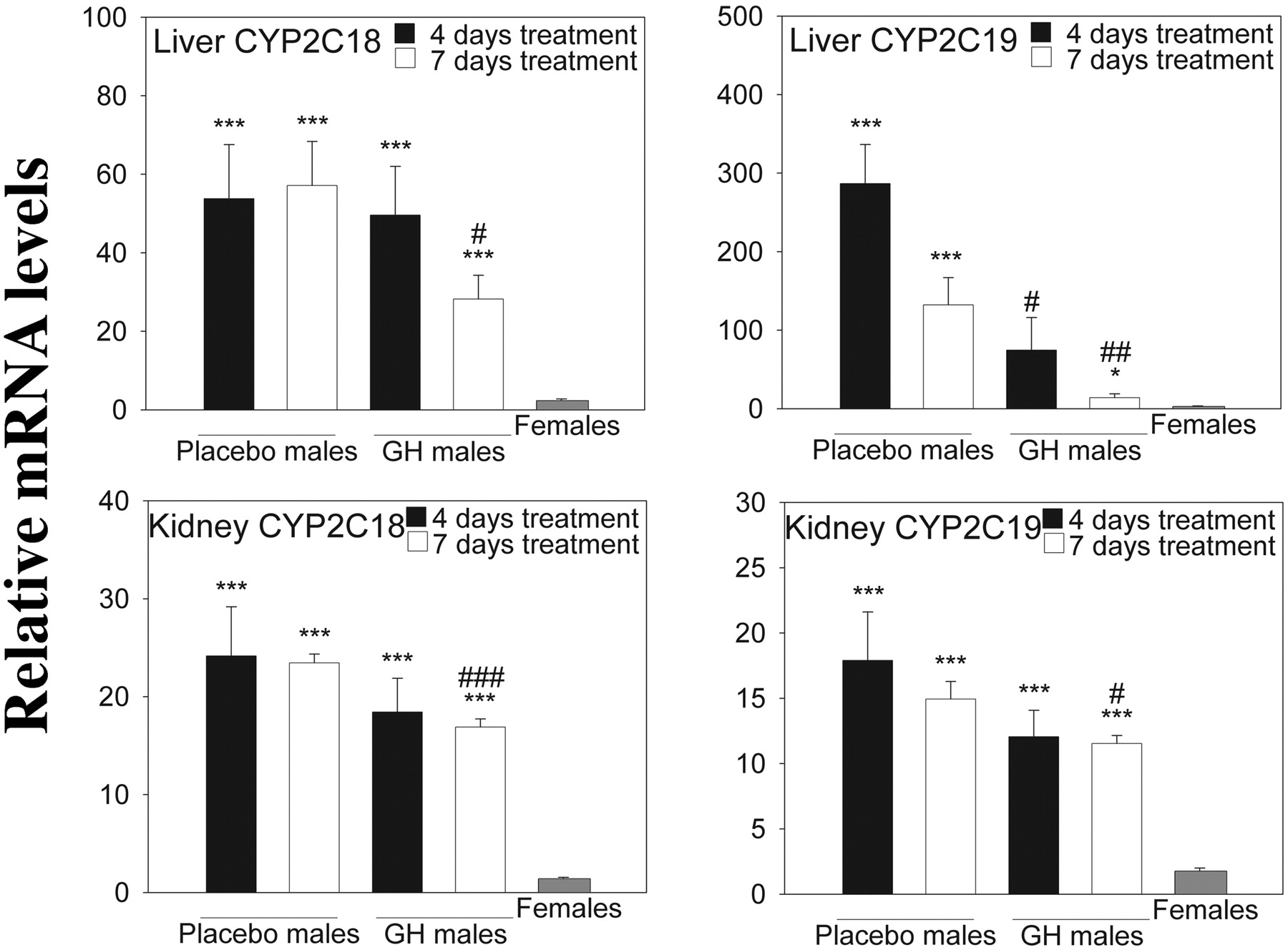
Effect of Male Castration and Female Testosterone Treatment on CYP2C Expression. Real-time PCR using gene-specific primers was used to monitor the expression of CYP2C18, CYP2C19, and of nine endogenous mouse Cyp2c genes in adult male mice castrated at 3 weeks of age. As shown in Fig. 1, castration dramatically decreased the expression of CYP2C18 and CYP2C19 in adult male mouse liver compared with that in the sham-operated control group (p ≤ 0.001). Both genes were expressed at much lower levels in both placebo and testosterone-treated females, but testosterone-treated females had a slightly higher hepatic expression compared with placebo females (p ≤ 0.05 for both genes). The same trend was observed in kidney (Fig. 1), but here the difference in expression between sham males and castrated males was smaller for CYP2C19 compared with CYP2C18. The effects of testosterone treatment in female mice were much more pronounced in the kidneys than in the liver, and for CYP2C19 testosterone increased the expression to that of untreated males. No gender- or treatment-related differences were seen for CYP2C18 or CYP2C19 in either brain or small intestine (data not shown).
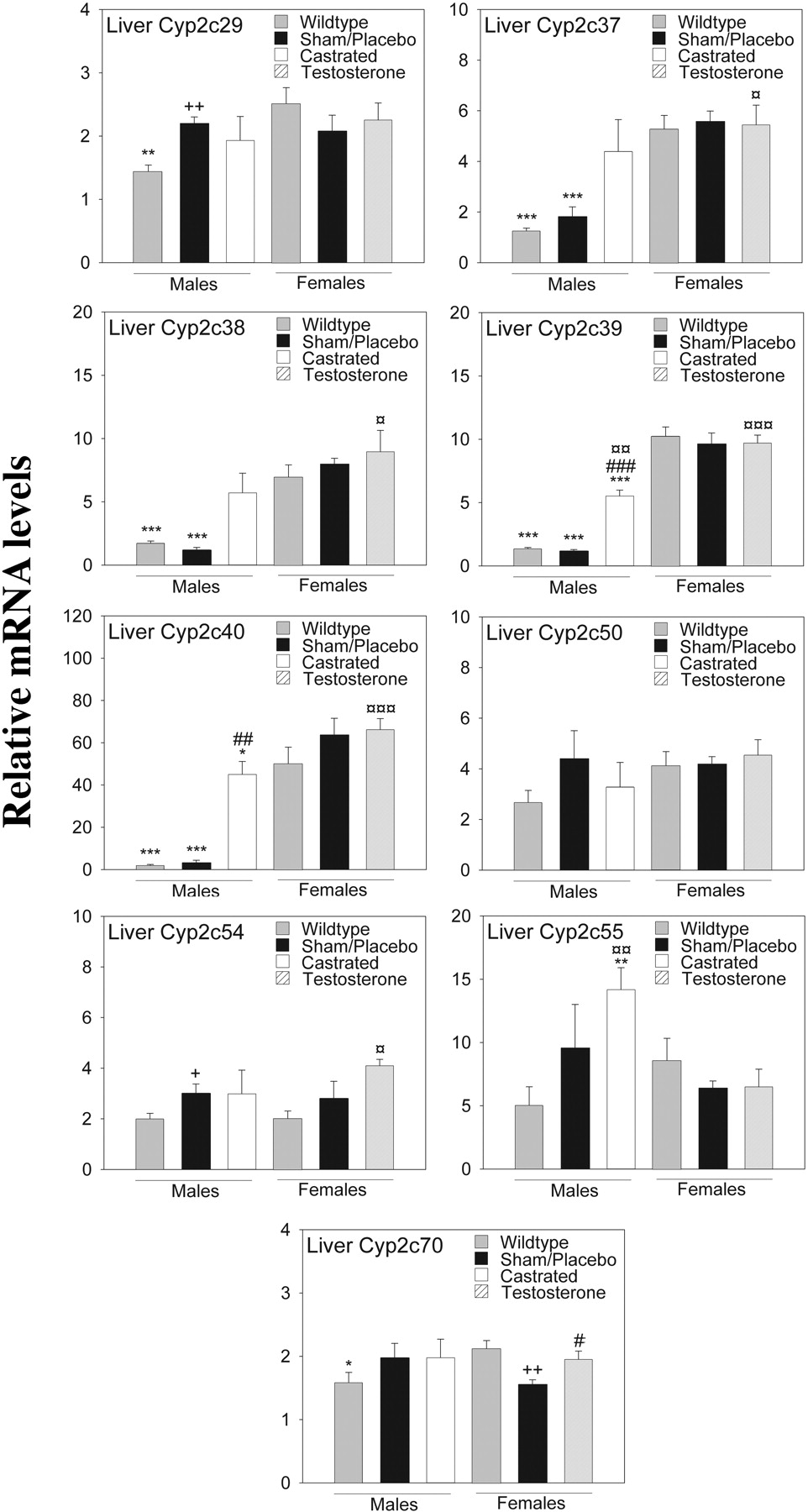
Of the nine endogenous mouse Cyp2c genes investigated, four showed a clear female-specific pattern of expression, i.e., Cyp2c37, Cyp2c38, Cyp2c39, and Cyp2c40 both in wild-type and transgenic mice. The transgenic expression of the CYP2C18 and CYP2C19 genes did not influence hepatic mRNA levels of the endogenous mouse Cyp2c genes (Fig. 2). A much smaller gender difference in expression characterized Cyp2c29 and Cyp2c70 when comparing wild-type males and wild-type females (p ≤ 0.01 for Cyp2c29 and p ≤ 0.05 for Cyp2c70). When comparing sham males with placebo females, no such gender differences were observed (Fig. 2). This gender-dependent expression for all six genes was the opposite of CYP2C18 and CYP2C19, the expression of which was higher in males than in females.
Next, we investigated the effect of castration on the expression of five mouse genes showing male-specific expression in liver (Cyp2d9, Cyp7b1, Gstπ, Mup1/2/6/8, and Mup3) (Supplemental Table 1). Seven female-specific liver genes identified in other studies were also examined (Cyp2a4, Cyp2b9, Cyp3a16, Cyp17a1, Cutl2, Trim 24, and Tox) (Supplemental Table 1) (Wiwi et al., 2004). For all five male-specific genes, castration decreased gene expression in male liver down to female expression levels. A significantly increased expression in castrated males (compared with sham-operated males) was observed for the female-specific genes Cyp2b9 and Tox (p ≤ 0.01 for both). A trend of increased expression was also seen for the other female-specific genes, except for Cyp3a16. It should be noted that all these female genes, with the exception of Cyp3a16, are up-regulated in male liver in the absence of STAT5b (Holloway et al., 2006; Laz et al., 2007).
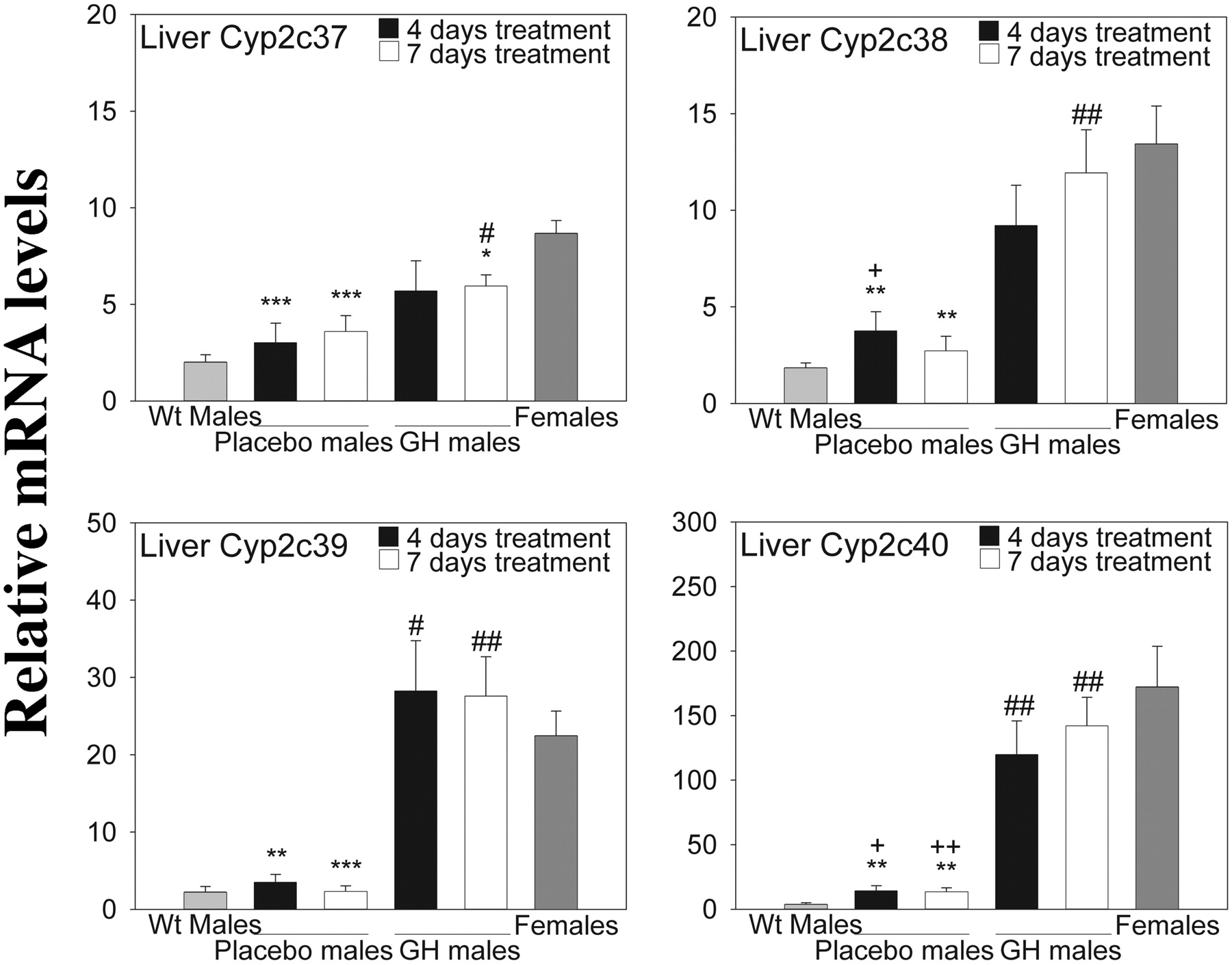
Effect of GH on mRNA Expression in Liver, Kidney, Brain, and Small Intestine. The CYP2C18 and CYP2C19 genes in both liver and kidney showed sexually dimorphic expression as shown in Fig. 3, and continuous GH administration to males caused a gradual, time-dependent drop in the expression levels of both genes in both tissues. The GH effect was more dramatic in liver and was much more pronounced for CYP2C19. As expected, no pronounced effect of GH was seen in brain or small intestine (data not shown). As a comparison, we investigated the effect of GH treatment on the hepatic expression of the endogenous gender-specific murine P450s that were affected by castration (Cyp2c37, Cyp2c38, Cyp2c39, and Cyp2c40) (Fig. 4). Cyp2c39 showed the most pronounced response to 4 days of continuous GH administration to males, with mRNA expression levels being slightly higher than in untreated females. Cyp2c37, Cyp2c38, and Cyp2c40 transcripts almost reached female expression levels after 7 days of GH treatment.
As a positive control for the effect of continuous GH treatment, we assayed mRNA levels of two male-specific genes (Cyp2d9 and Mup1/2/6/8) and two female-specific genes (Cyp2a4 and Cyp2b9) (Supplemental Table 2). The most pronounced effect was on the male-specific genes, where a decrease in mRNA levels was observed after 4 days of continuous GH treatment, and an even greater decrease was seen after 7 days. The effect of continuous GH administration on the female-specific genes, Cyp2a4 and Cyp2b9, was not as pronounced, and the GH-treated male mice never attained full female RNA levels.
Discussion
The sexually dimorphic expression of several hepatic drug-metabolizing P450 enzymes has been shown and studied extensively in mice and rats. The present study shows that in a transgenic mouse model, the human CYP2C19 and CYP2C18 genes are regulated in a sexually dimorphic manner and are under similar regulation by GH and castration as sexually dimorphic endogenous Cyp2c genes. This observation suggests regulatory elements of these genes might also be subject to GH and hormone-dependent regulation in humans.

Relative CYP2C18 and CYP2C19 mRNA expression in transgenic mice approximately 11 weeks old. Reverse-transcribed total RNA and gene-specific primers were used as described under Materials and Methods. The geometric mean of two reference transcripts (GAPDH and HPRT) was used to normalize expression levels. Data are presented relative to the lowest individual expression level for each gene transcript and organ (mean ± S.E.). Each group contains four or five mice (sham-treated males, n = 4; all the other groups, n = 5). Analyses were performed in triplicate. Experimental groups were compared using Student's t test. The location of the statistical symbol for each respective comparison is denoted by boldface type: * and ***, p ≤ 0.05 and p ≤ 0.001, respectively, for sham-operated males versus placebo-treated females or castrated males versus testosterone treated females. #, ##, and ###, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for sham-operated males versus castrated males or placebo-treated females versus testosterone-treated females.  and
and  , p ≤ 0.01 and p ≤ 0.001, respectively, for sham-operated male versus testosterone-treated females or placebo-treated females versus castrated males.
, p ≤ 0.01 and p ≤ 0.001, respectively, for sham-operated male versus testosterone-treated females or placebo-treated females versus castrated males.
Growth hormone, acting via its gender-specific pituitary profiles, regulates the expression of many gender-specific and gender-predominant genes, including P450 genes. GH is secreted in pulses but with different levels and frequencies in males and females in several species, including humans (van den Berg et al., 1996; Jaffe et al., 1998). Treatment with exogenous GH has been shown to alter CYP2C11 and CYP2C12 expression in rats (Waxman et al., 1991; Kawai et al., 2001), whereas mouse Cyp2c29 was shown to be gender-independent and GH archetype-independent (Jarukamjorn et al., 2006). In humans, CYP2C19 activity has been shown to be stimulated in healthy elderly men by subcutaneous injections of GH once daily (Jürgens et al., 2002). The present study shows that GH is crucial in the sexually dimorphic expression of CYP2C18 and CYP2C19 in the CYP2C18/CYP2C19 humanized mouse model. Thus, the human CYP2C18 and CYP2C19 genes, when inserted in this transgenic mouse model, contain all the DNA sequences required to respond to the endogenous mouse hormonal environment, including the regulatory elements required for responsiveness to exogenously administered GH.
STAT5b has been identified as an essential determinant of GH-mediated sexual expression of P450 enzymes (Clodfelter et al., 2006; Holloway et al., 2006). However, the precise intracellular regulatory mechanisms activated by STAT5b, leading to the regulation of hepatic P450 expression, are only partially understood and await more extensive investigation. Although a STAT5b consensus sequence was identified in the promoter region of CYP2C19, no protein binding could be detected in electrophoretic mobility shift assay studies despite the fact that a positive control oligonucleotide for STAT5b showed binding that was supershifted with STAT5b antibodies (data not shown). The GH and androgen responsive elements in the CYP2C18 and CYP2C19 genes will be the focus of future studies.
Castration caused CYP2C18 and CYP2C19 mRNA levels in males to be reduced to female levels, a response that was also seen for several endogenous male-specific mouse liver genes (Supplemental Table 1). This is believed to reflect the role of testicular androgens in maintaining the characteristic gender-dependent patterns of pituitary GH secretion in adult male rats (Jansson et al., 1985, Jansson et al., 1985). Continuous GH treatment of intact male CYP2C18/CYP2C19 transgenic mice suppressed liver CYP2C19, as well as the endogenous male-specific mouse genes Cyp2d9 and Mup1/2/6/8 to near-female levels. Only partial (∼50%) suppression of liver CYP2C18 expression was achieved, which could be explained if CYP2C18 mRNA has a long intrinsic half-life or, alternatively, could indicate that other hormonal factors, in addition to continuous GH, may be required to achieve complete feminization. CYP2C18 and CYP2C19 were largely unresponsive to GH treatment in the kidney, consistent with earlier studies showing that male-specific P450 gene expression in kidney is primarily regulated by androgen by a mechanism that is independent of GH (Henderson and Wolf, 1991; Imaoka et al., 1992; Sundseth and Waxman, 1992).
Testosterone treatment in female transgenic mice did not have as drastic an effect on the mRNA levels as did castration. In naive males, testosterone is not secreted at constant levels but rather in an episodic manner (Mock et al., 1978), which was not the case in our study. The lack of pulsatility may be the reason for the limited response in testosterone-treated females. Another possible explanation is the continued presence of estrogen in the testosterone-treated females, which may antagonize the effect of androgen treatment. A further possibility is the time period under which the study was conducted. The testosterone capsules used were reported to have a 60-day release period, and the necropsies were conducted after 55 to 56 days. It may be possible that the testosterone levels were lower at the end of the study or that the transgenic females metabolize testosterone faster, causing testosterone levels to fall below the required male physiological levels.
The GH-dependent changes in hepatic P450 expression described here may significantly alter the pharmacokinetics and pharmacodynamics of drugs metabolized by CYP2C19. The interactions between GH and P450-metabolized drugs may lead to changes in drug efficacy and, as a consequence, to adverse effects. The P450-humanized mouse model presented here and the GH-responsive CYP3A4/CYP3A7 transgenic model described earlier (Cheung et al., 2006) may contribute to the understanding of GH regulation of human P450 genes. Use of these models in medical testing may reduce differences in drug metabolism caused by interspecies variation in P450-mediated metabolism. However, other physiological parameters and hormone profiles also vary between mice and humans, and there are metabolic differences as a result of the involvement also of the endogenous mouse enzymes. Although rodents have more drastic differences in their GH plasma levels than humans, humans show significant differences in pituitary GH-release patterns between genders. These species differences, as well as other physiological parameters, can make extrapolation from animal models to humans difficult and thus must be taken into consideration when evaluating results obtained from these types of models.

Relative murine hepatic CYP2C mRNA expression in transgenic (sham-operated, placebo, and testosterone-treated) and wild-type mice approximately 11 weeks old. Reverse-transcribed total RNA from liver and gene-specific primers was used as described under Materials and Methods. The geometric mean of two reference transcripts (GAPDH and β-actin) was used to normalize expression levels. Data are presented relative to the lowest individual expression level for each gene transcript (mean ± S.E.). Each group contains three to five mice (sham-operated males, n = 3; castrated males, n = 4; all the other groups, n = 5). Analyses were performed in triplicate. Experimental groups were compared using Student's t test. The location of the statistical symbol for each respective comparison is denoted by boldface type: *, **, and ***, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for wild-type males versus wild-type females or sham-operated males versus placebo-treated females or castrated males versus testosterone-treated females. #, ##, and ###, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for sham-operated males versus castrated males or placebo-treated females versus testosterone-treated females.  ,
,  , and
, and  , p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for sham-operated males versus testosterone-treated females or placebo-treated females versus castrated males. + and ++, p ≤ 0.05 and p ≤ 0.01, respectively, for wild-type males versus sham-operated males or wild-type females versus placebo-treated females.
, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for sham-operated males versus testosterone-treated females or placebo-treated females versus castrated males. + and ++, p ≤ 0.05 and p ≤ 0.01, respectively, for wild-type males versus sham-operated males or wild-type females versus placebo-treated females.
In conclusion, the human CYP2C18/CYP2C19 transgenic mouse line described here provides a novel tool for studying the regulation and function of human CYP2C18 and CYP2C19 genes. Sexually dimorphic expression of CYP2C18 and CYP2C19 and murine Cyp2c37/38/39 and Cyp2c40 genes was observed in the mice, which may lead to significant gender differences in drug metabolism, disposition, pharmacokinetics, and pharmacodynamics. Although the underlying mechanism for regulation of these sexually dimorphic genes could not be fully determined, this animal model may help in studying the regulation of human CYP2C18 and CYP2C19 in a whole-animal system.

Relative CYP2C18 and CYP2C19 mRNA expression in transgenic mice, approximately 11 weeks old. Reverse-transcribed total RNA and gene-specific primers were used as described under Materials and Methods. The geometric mean of two reference transcripts (GAPDH and HPRT) was used to normalize expression levels. Data are presented relative to the lowest individual expression level for each gene transcript and organ (mean ± S.E.). Each group contains four to eight mice (placebo-treated males 4- and 7-day exposures, n = 4; GH-treated males 4-day exposure, n = 5; GH-treated males 7-day exposure, n = 6; and transgenic females, n = 7). Analyses were performed in duplicate. Experimental groups were compared using Student's t test. The location of the statistical symbol for each respective comparison is denoted by boldface type: * and ***, p ≤ 0.05 and p ≤ 0.001, respectively, for placebo-treated males (4 days) versus transgenic female, placebo-treated males (7 days) versus transgenic female, GH-treated males (4 days) versus transgenic females or GH-treated males (7 days) versus transgenic females. #, ##, and ###, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for placebo-treated males (4 days) versus GH-treated males (4 days) or placebo-treated males (7 days) versus GH-treated males (7 days).

Relative hepatic expression of gender-specific murine CYP2C transcripts in transgenic mice, approximately 11 weeks old. Reverse-transcribed total RNA from liver and gene-specific primers was used as described under Materials and Methods. The geometric mean of two reference transcripts (GAPDH and HPRT) was used to normalize expression levels. Data are presented relative to the lowest individual expression level for each gene transcript (mean ± S.E.). Each group contains four to eight mice [placebo-treated males (4 and 7 days) n = 4, GH-treated males (4 days) n = 5, GH-treated male (7 days) n = 6, transgenic females n = 7, and wild-type males n = 8]. Analyses were performed in duplicate. Experimental groups were compared using Student's t test. The location of the statistical symbol for each respective comparison is denoted by boldface type: *, **, and ***, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively, for placebo-treated males (4 days) versus transgenic female, placebo-treated males (7 days) versus transgenic female, GH-treated males (4 days) versus transgenic females or GH-treated males (7 days) versus transgenic females. # and ##, p ≤ 0.05 and p ≤ 0.01, respectively, for placebo-treated males (4 days) versus GH-treated males (4 days) or placebo-treated males (7 days) versus GH-treated males (7 days). + and ++, p ≤ 0.05 and p ≤ 0.01, respectively, for wild-type male versus placebo male (4 days) or wild-type male versus placebo male (7 days).
Acknowledgments
We thank Anne Carlberg for assistance in the mouse castration study. We also thank the personnel at the Karolinska Institute animal facilities for care of the mice during the in vivo parts of our studies.
Footnotes
-
This work was supported in part by the National Institutes of Health National Institute of Diabetes and Digestive and Kidney Diseases [Grant DK33765]; and The Swedish Research Council.
-
S.L. and R.M.B. contributed equally to this work.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.109.026963.
-
ABBREVIATIONS: P450, cytochrome P450; GH, growth hormone; STAT, signal transducer and activator of transcription; PCR, polymerase chain reaction; gDNA, genomic DNA; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HPRT, hypoxanthine guanine phosphoribosyltransferase; CLint, intrinsic clearance.
-
↵
 The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material. -
↵1 Current affiliation: Department of Biopharmaceutical Sciences, School of Pharmacy, University of California San Francisco, San Francisco, California.
- Accepted March 26, 2009.
- Received January 29, 2009.
- The American Society for Pharmacology and Experimental Therapeutics






{kind=link}
{kind=link}
{kind=link}
{kind=link}