


Abstract
Ezetimibe lowers plasma cholesterol levels by inhibiting the uptake of cholesterol in the intestine. Because of the extensive enterohepatic circulation of ezetimibe, relatively low doses are required to be effective. In blood and bile the majority of ezetimibe is present as a glucuronide conjugate, which is formed in the enterocyte. Presently, it is not clear which mechanisms are responsible for this efficient enterohepatic circulation. Abcc2, Abcc3, and Abcg2 are ATP-binding cassette (ABC) transporters that are expressed in both liver and intestine and are capable of transporting glucuronidated compounds. The aim of this study was to investigate the contribution of these transporters in the enterohepatic cycling of ezetimibe glucuronide (Ez-gluc). Transport studies were performed in plasma membrane vesicles from ABCC2-, ABCC3-, and ABCG2-expressing Sf21 insect cells. Furthermore, intestinal explants from wild-type and Abcc3(–/–) mice were used to study vectorial transport in a Ussing chamber setup. Finally, biliary excretion of Ez-gluc was measured in vivo after duodenal delivery of ezetimibe in wild-type, Abcc3(–/–), Abcc2(–/–), Abcg2(–/–), and Abcg2(–/–)/Abcc2(–/–) mice. ABCC3-, ABCC2-, and ABCG2-mediated transport was dose dependently inhibited by Ez-gluc. In the Ussing chamber Ez-gluc recovered from the basolateral side was significantly reduced in duodenal (2.2%), in jejunal (23%), and in ileal (23%) tissue of Abcc3(–/–) mice compared with that in tissues of wild-type mice. Biliary excretion of Ez-gluc was significantly reduced in Abcc3(–/–) (34%), Abcc2(–/–) (56%), and Abcg2(–/–)/Abcc2(–/–) (2.5%) compared with that in wild-type mice. These data demonstrate that the enterohepatic circulation of Ez-gluc strongly depends on the joint function of Abcc3, Abcc2, and Abcg2.
Hypercholesterolemia can be treated by inhibition of cholesterol synthesis with statins and by inhibition of intestinal uptake of cholesterol. The latter can be achieved with ezetimibe (van Heek et al., 1997, 2000), which potently inhibits Niemann-Pick C1-like 1 protein (Altmann et al., 2004; Davis et al., 2004). Ezetimibe itself is also taken up by enterocytes and extensively glucuronidated (van Heek et al., 2000). The major part of ezetimibe is conjugated to the benzylic glucuronide by UDP-glucuronosyltransferases 1A1, 1A3, and 2B15. A minor amount of ezetimibe is conjugated to the phenolic glucuronide by UDP-glucuronosyltransferase 2B7 (Ghosal et al., 2004). Clearance of ezetimibe glucuronide (Ez-gluc) from blood is not a straightforward process as the concentration-time profiles exhibit multiple peaks (Ezzet et al., 2001; Patrick et al., 2002). The multiple peaks are thought to be caused by enterohepatic circulation: after glucuronidation in the enterocyte, ezetimibe travels via blood to the liver and is extruded into bile (van Heek et al., 2000). The excretion of Ez-gluc from the enterocyte into blood is likely to be transporter-mediated. ABCC3 is a potential candidate transporter for this step because it accepts glucuronidated compounds, such as acetaminophen glucuronide (Manautou et al., 2005) and is expressed in the basolateral membrane of enterocytes (Scheffer et al., 2002). The biliary excretion of Ez-gluc is also likely to be transporter-mediated. Oswald et al. (2006a) demonstrated that transport of the model substrate estradiol-17β-glucuronide via ABCC2 is inhibited by Ez-gluc. This contention was further supported by the finding that the AUC for plasma Ez-gluc in Abcc2-deficient rats is significantly increased compared with that for wild-type rats (Oswald et al., 2006b). To test the relative role of Abcc2 as well as that of another canalicular transporter, Abcg2, we studied biliary elimination in Abcc2(–/–), Abcg2(–/–), and double-knockout mice.
Materials and Methods
Materials. Ezetimibe was obtained from Merck Sharp and Dohme (Hertfordshire, UK). Acetonitrile (HPLC grade) was bought from Mallinckrodt Baker (Deventer, The Netherlands). [3H]Estradiol-17β-glucuronide (E217βG) and [3H]methotrexate (MTX) were purchased from GE Healthcare (Roosendaal, The Netherlands). Ez-gluc was synthesized as follows: pooled mouse liver microsomes (2.5 mg/ml) were incubated during 2 h with ezetimibe (10 mM), MgCl2 (10 mM), and UDP-glucuronide (20 mM) in a Tris-HCl buffer (0.1 M, pH 7.4). Thereafter, Ez-gluc was purified by use of reverse-phase HPLC (see below). All other chemicals and reagents were bought from Sigma-Aldrich (Zwijndrecht, The Netherlands).
Animals. Mice were housed and handled according to institutional guidelines complying with Dutch legislation. All animal experiments were approved by the Institutional Animal Care and Use Committee of the Academic Medical Center.
Wild-type and Abcc3(–/–) (Zelcer et al., 2006) mice (against a 99% FVB/N background) were bred at the Animal Institute of the Academic Medical Center. Abcc2(–/–) (Vlaming et al., 2006), Abcg2(–/–) (Jonker et al., 2002), and Abcg2(–/–)/Abcc2(–/–) (Vlaming et al., 2009) mice (all against a 99% FVB/N background) were bred at The Netherlands Cancer Institute (Amsterdam, The Netherlands). The animals used were between 9 and 17 weeks of age. Animals were kept in a temperature-controlled environment with a 12-h light/dark cycle. They received a standard diet (AM-II; Hope Farms, Woerden, The Netherlands) ad libitum.
Methods.Preparation of membrane vesicles. The cDNA of the ABCC2 gene (Paulusma et al., 1997) [a kind gift from Professor P. Borst (The Netherlands Cancer Institute)] was cloned into the pFastBac1 (Invitrogen Breda, The Netherlands) donor plasmid. This was followed by transformation of the donor plasmid into DH10Bac (Invitrogen) cells, which allowed for transposition of the ABCC2 gene into bacmid DNA. ABCG2 and ABCC3 recombinant baculoviruses were a kind gift from Professor P. Borst (Zelcer et al., 2001; Breedveld et al., 2004). Sf21 cells grown at 27°C were infected with ABCC2, ABCC3,or ABCG2 recombinant baculovirus. Cells were harvested 2 days (ABCC3) or 3 days (ABCC2 and ABCG2) postinfection and resuspended in ice-cold hypotonic buffer (250 mM mannitol-20 mM HEPES/Tris (pH 7.4)-2 mM EDTA) supplemented with protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 1 μg/ml aprotinin, 1 μg/ml leupeptin, and 1 μg/ml pepstatin) and incubated for 30 to 60 min on ice. Cells were homogenized using a tight-fitting Dounce homogenizer. The cell homogenate was layered on top of a discontinuous sucrose gradient (i.e., 56, 38, and 19% sucrose) and centrifuged for 3.2 h at 141,000g [SW28 rotor and a Beckman Optima L-90K centrifuge (Beckman Coulter, Mijdrecht, The Netherlands)]. The 19 to 38% interface was collected, washed, and resuspended in 250 mM sucrose-20 mM HEPES/Tris (pH 7.4). Membrane vesicles were obtained after revesiculation of the preparation by passing it 30 times through a 27-gauge needle. Vesicles were aliquoted and stored at –80°C until use. Protein content was determined using the bicinchoninic acid method.
Western blotting and protein analysis. Membrane vesicles were fractionated by 6% SDS-polyacrylamide gel electrophoresis and blotted on nitrocellulose membranes (Whatman Schleicher & Schuell, Dassel, Germany), which were blocked in phosphate-buffered saline-5% milk powder-0.05% Tween 20. The following antibodies were used: anti-ABCG2, BXP-21; anti-ABCC2, M2III6; and anti-ABCC3, M3III21 [all antibodies were kind gifts from Dr. G. L. Scheffer (Free University Hospital, Amsterdam, The Netherlands)]. Immune complexes were visualized with horseradish peroxidase-conjugated immunoglobulins (Bio-Rad, Veenendaal, The Netherlands) and detected using chemiluminescence (GE Healthcare).
Vesicular transport assays. The vesicular transport studies were performed using the rapid filtration technique as described previously (Heijn et al., 1992). In brief, 30 μl of membrane vesicles (10–30 μg of protein) were added to 30 μl of prewarmed (37°C) reaction mixture containing [3H]estradiol-17β-glucuronide or [3H]methotrexate (0.05 μCi in 250 mM sucrose, 20 mM HEPES/Tris (pH 7.4)-20 mM MgCl2-30 mM creatine phosphate-1.2 mg of creatine kinase/ml with or without ATP (8 mM). Uptake was terminated by the addition of 20 volumes of ice-cold 250 mM sucrose and 20 mM HEPES/Tris (pH 7.4). The suspension was immediately filtered through 0.2-μm nitrocellulose filters under vacuum. The filters were washed three times and counted in a liquid scintillation counter.
Calculation of half-maximal inhibition. Transport of [3H]estradiol-17β-glucuronide or [3H]methotrexate was plotted against Ez-gluc concentration. A linear trend matching known data points, using the least-squares method, was acquired. From this formula, the concentration of Ez-gluc, at which half-maximal inhibition was achieved, was calculated.
Ussing chamber experiments. Duodenal, jejunal, and ileal tissues from wild-type and Abcc3(–/–) animals were applied to a holder that was subsequently mounted in a Ussing chamber. The apical and basolateral compartments contained 1.5 ml of buffer, which consisted of Krebs bicarbonate with 2 mM glutamine and 10 mM HEPES (pH 7.4) and were continuously gassed with carbogene. Temperature was set at 37 ± 1°C. After a 5-min equilibration the experiment was started by the addition of 2 μmol of ezetimibe (in 2% dimethyl sulfoxide) at the apical side.

Western blot of membrane vesicles of Sf21 insect cells uninfected and infected with ABCC2, ABCC3, and ABCG2 cDNA containing baculovirus, respectively. Lane 1, molecular ladder; lanes 2, 5, and 8, (uninfected) Sf21 cells; lanes 4 and 7, empty; lane 3, Sf21-ABCG2; lane 6, Sf21-ABCC2; and lane 9, Sf21-ABCC3. The blot was cut in three parts (cut in the middle of lanes 4 and 7) and incubated with antibodies against ABCG2 (lanes 1–3), ABCC2 (lanes 5 and 6), and ABCC3 (lanes 8 and 9), respectively. Shown is the whole blot rearranged.
In vivo experiment: duodenal administration of ezetimibe and subsequent measurement of biliary Ez-gluc. Male mice were anesthetized with a combination of Hypnorm (11.8 mg/kg fluanisone and 0.37 mg/kg fentanyl citrate; VetaPharma, Leeds, UK) and Valium (5.9 mg/kg diazepam; Centrafarm, Etten-Leur, The Netherlands). Body temperature was maintained between 35 and 37°C by keeping the mice on thermostatted heating pads. Thereafter, the vena jugularis was cannulated, and tauroursodeoxycholate was infused at a constant rate of 1 μmol/min/100 g of mouse, followed by cannulation of the gallbladder. After ligation of the stomach just before the duodenum, 1 μmol of ezetimibe was injected into the duodenum. Subsequently, bile was collected every 10 min for 2 h. Furthermore, at 10 min portal blood (using a 1-ml syringe and a 30-gauge needle) was drawn, and at 120 min peripheral blood (heart puncture) was drawn. In addition, livers were collected.
Quantification of Ez-gluc. Biliary Ez-gluc was quantified by reverse-phase HPLC. In brief, 400 times diluted bile was applied to an OmniSpher HPLC column (3 μm, 20 cm; Varian, Middelburg, The Netherlands). The starting eluent consisted of 10% acetonitrile-90% 20 mM ammonium formate (pH 3.5), followed by a linear gradient to 70% acetonitrile in the same buffer. Ez-gluc eluted from the column at a retention time of 11 min and ezetimibe at 16 min. Ezetimibe and its glucuronide were detected at 245 nm and quantitated by using a calibration curve of ezetimibe, with the assumption that ezetimibe and its glucuronide have the same extinction coefficient.
Statistical Analyses. Statistical differences (p < 0.05) were determined by one-way analysis of variance, followed by a multiple comparison test or unpaired Student's t test. All data are expressed as means ± S.D.
Results
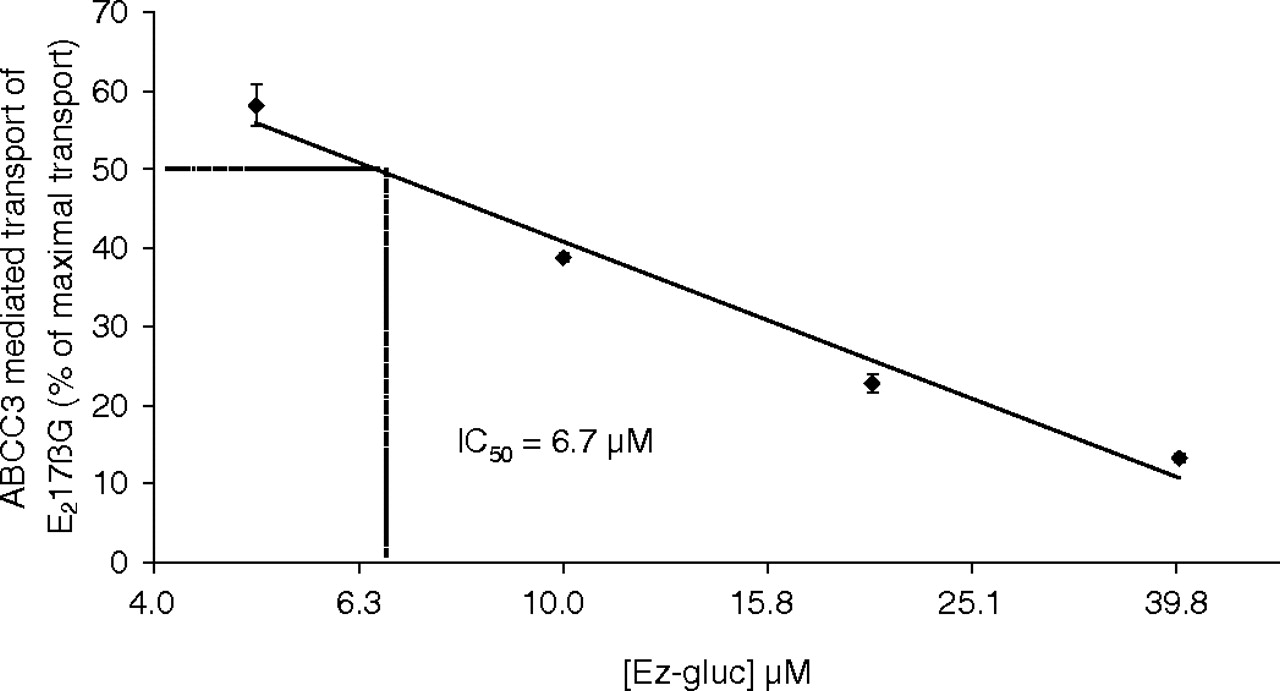
To measure the role of ABCC3 in transport of Ez-gluc, an inhibition experiment was performed with ABCC3-containing membrane vesicles (Fig. 1) and the ABCC3/Abcc3 model substrate E217βG. Ez-gluc inhibits this transport in a dose-dependent way (Fig. 2). Half-maximal inhibition (IC50) of ABCC3-mediated transport of E217βG was achieved at an Ez-gluc concentration of 6.7 μM. This finding could indicate that ABCC3/Abcc3 is able to mediate the transport of Ez-gluc. We tested this hypothesis by examining transport in a Ussing chamber setup with intestinal explants from Abcc3(–/–) and wild-type mice. Apical administration of ezetimibe to duodenal, jejunal, and ileal tissue explants led to a time-dependent increase of Ez-gluc on the basolateral side. Basolateral excretion of Ez-gluc was significantly reduced in duodenal (to 2.2%), in jejunal (to 23%), and in ileal (to 23%) tissue of Abcc3(–/–) mice compared with that in tissues of wild-type mice (Fig. 3).
To investigate the role of Abcc3 in the enterohepatic circulation we injected ezetimibe in the duodenum in vivo and measured the amount of Ez-gluc recovered in bile. Between 10 and 20 min after administration of ezetimibe in the duodenum, a peak of Ez-gluc in bile was seen in both Abcc3(–/–) and wild-type mice (Fig. 4A). However, the total excretion of Ez-gluc in bile after 2 h was significantly reduced in Abcc3(–/–) mice (to 34%) compared with that in wild-type mice (Fig. 4). We further examined whether the transport from enterocyte to blood by Abcc3 was rate-limiting by drawing portal blood at 10 min after duodenal injection. The portal blood concentration was found to be reduced in Abcc3(–/–) mice (to 7.1% of that in wild-type mice) (Fig. 4B). This result indicates a rate-limiting role of Abcc3 in the transport of Ez-gluc from the cytosol of enterocytes into blood.

Ez-gluc inhibition of E217βG transport into Sf21 membrane vesicles containing ABCC3 protein. The values are expressed as a percentage of maximal ATP-dependent transport. Shown is the average ± S.D. of an experiment with triplicate incubations. Note the logarithmic x-axis. A linear trend matching known data points, using the least-squares method, was acquired. From this formula the IC50 value was calculated.
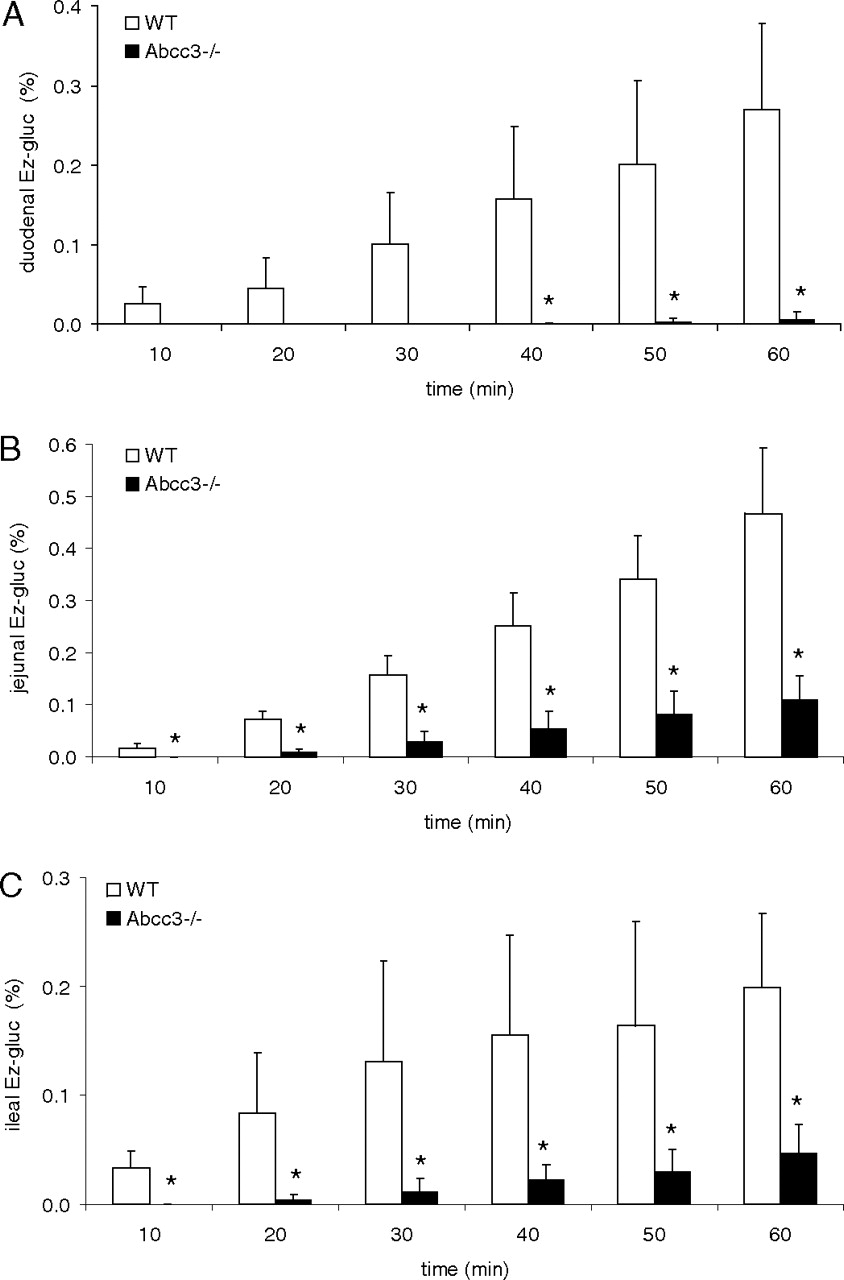
Appearance of glucuronidated ezetimibe on the basolateral side of intestinal explants from Abcc3(–/–) and wild-type (WT) mice: A, duodenum; B, jejunum; C, ileum. Ezetimibe (2 μmol) was applied to the apical compartment of Ussing chambers. At the indicated time points samples were taken from the basolateral side and analyzed by HPLC. Shown is the average ± S.D. of at least three experiments with triplicate incubations. *, p < 0.05, comparing the appearance of Ez-gluc using tissue from Abcc3(–/–) versus wild-type mice for the indicated time points.

Distribution of Ez-gluc after duodenal administration of 1.0 μmol of ezetimibe in Abcc3(–/–) and wild-type (wt) mice. A, appearance of Ez-gluc at the indicated time points in bile. B, total appearance of Ez-gluc in bile and also hepatic and peripheral blood levels of Ez-gluc 2 h after administration of ezetimibe. The portal blood concentration of Ez-gluc was determined in a sample taken at 10 min after administration. Shown is the average ± S.D. of at least three mice. note the logarithmic y-axis. #, p < 0.05, comparing the appearance of Ez-gluc in bile of Abcc3(–/–) versus wild-type mice for the indicated time points. *, p < 0.05, comparing levels of Ez-gluc in bile, liver, and portal blood from Abcc3(–/–) versus wild-type mice.
Both ABCC2 and ABCG2 are potential candidates to mediate biliary transport of Ez-gluc. Therefore, inhibition experiments were performed with ABCC2-containing membrane vesicles (Fig. 1) using the ABCC2 model substrate E217βG (Fig. 5A) and ABCG2-containing membrane vesicles (Fig. 1) using the ABCG2 model substrate MTX (Fig. 5B). Transport of E217βG mediated by ABCC2 and transport of MTX mediated by ABCG2 was dose dependently inhibited by Ez-gluc, with IC50 values of 34.3 and 51.8 μM, respectively. To investigate the relative roles of murine Abcc2 and Abcg2 in biliary excretion of Ez-gluc, ezetimibe was injected into the duodenum of wild-type, Abcc2(–/–), Abcg2(–/–), and Abcg2(–/–)/Abcc2(–/–) mice. Subsequently, bile was collected during 2 h, and Ez-gluc was quantified (Fig. 6). The total excretion of Ez-gluc in bile after 2 h was similar in Abcg2(–/–) and wild-type mice, diminished in Abcc2(–/–) mice (to 56%), but severely reduced in Abcg2(–/–)/Abcc2(–/–) mice (to 2.5%) in comparison with that in wild-type mice (Fig. 6A). Concomitantly, we observed 270-fold higher plasma levels and 70-fold higher hepatic levels of Ez-gluc in Abcg2(–/–)/Abcc2(–/–) mice compared with those in wild-type mice at 2 h after duodenal administration (Fig. 6B). This finding underscores the pivotal role for the joint function of Abcc2 and Abcg2 in the biliary excretion of Ez-gluc.

Ez-gluc inhibition of (A) E217βG transport into Sf21 membrane vesicles containing ABCC2 protein and (B) methotrexate transport into Sf21 membrane vesicles containing ABCG2 protein. The values are expressed as a percentage of maximal ATP-dependent transport. Shown is the average ± S.D. of an experiment with triplicate incubations. Please note the logarithmic x-axis. A linear trend matching known data points, using the least-squares method, was acquired. From this formula the IC50 value was calculated.
Discussion
ABCC3 transport in liver is considered to have a dual role. On the one hand, it diverts certain glucuronides, formed in the liver, toward urinary excretion. As an example, in the mouse, acetaminophen glucuronide is transported via Abcc3 and is excreted exclusively into urine. On the other hand, under cholestatic conditions Abcc3 constitutes an escape route for compounds that are normally transported into bile (e.g., bilirubin glucuronide) (Donner and Keppler, 2001; Soroka et al., 2001). The latter function is underscored by higher expression of ABCC3 during cholestasis in humans and rodents and in Abcc2-deficient TR– rats and Abcc2(–/–) mice (Johnson et al., 2006; Vlaming et al., 2006). However, its role in the intestine has been less well explored. It was proposed to have a role in the enterohepatic circulation of bile salts in the rat as the transporter for bile salts from the enterocytes into blood (Hirohashi et al., 2000). However, Abcc3-deficient mice showed no problem in vectorial transport of bile salts from the intestine to blood (Belinsky et al., 2005; Zelcer et al., 2006). In contrast, Abcc3 could play a direct role in basolateral transport of drugs and their conjugates and, as a consequence, an indirect role in the enterohepatic circulation of these drugs. ABCC3/Abcc3 is able to transport glucuronate conjugates of compounds such as bilirubin, acetaminophen, and taurohyodeoxycholate (Belinsky et al., 2005; Manautou et al., 2005; Zelcer et al., 2006). In this respect ezetimibe is an attractive model drug for Abcc3 function because it is conjugated in the intestine with glucuronic acid, and it undergoes extensive enterohepatic circulation. In this report we show that ABCC3-mediated transport of the glucuronidated model compound estradiol-17β is inhibited by Ez-gluc, indicating that the latter is likely to be a substrate for ABCC3. In Ussing chamber experiments, we showed that Abcc3 deficiency leads to strongly impaired transport of Ez-gluc from enterocyte to the basolateral side. Subsequent in vivo experiments demonstrated that transport of Ez-gluc across the intestinal epithelium strongly depends on Abcc3 function; i.e., little Ez-gluc could be found in the portal blood of Abcc3(–/–) mice.

Distribution of Ez-gluc after duodenal administration of 1.0 μmol ezetimibe in Abcg2(–/–), Abcc2(–/–), Abcg2(–/–)/Abcc2(–/–), and wild-type (wt) mice. A, appearance of Ez-gluc at the indicated time points in bile. B, total appearance in bile and also hepatic and peripheral blood levels of Ez-gluc 2 h after administration of ezetimibe. Shown is the average ± S.D. of at least three mice. #, p < 0.05, comparing appearance of Ez-gluc in the bile of Abcc2(–/–) versus wild-type mice for the indicated time points. †, p < 0.05, comparing appearance of Ez-gluc in bile of Abcg2/Abcc2(–/–) versus wild-type mice for the indicated time points. •, p < 0.05, comparing levels of Ez-gluc in bile and liver Abcc2(–/–) versus wild-type mice. *, p < 0.05, comparing levels of Ez-gluc in bile, liver, and peripheral blood from Abcg2/Abcc2(–/–) versus wild-type mice.
Oswald et al. (2006a) presented indirect evidence that Ez-gluc is a substrate of ABCC2/Abcc2. First, they showed that transport of a model compound for ABCC2 was inhibited by Ez-gluc (Oswald et al., 2006a). Our study confirms this finding because we also showed that ABCC2-mediated transport of the model compound E217βG was inhibited by Ez-gluc. Second, Oswald et al. (2006b) showed severely increased AUC from time 0 to 24 h (AUC0–24 h) after oral dosing of ezetimibe to Abcc2-deficient TR– and wild-type rats (Oswald et al., 2006b). However, a complicating factor in the interpretation of this experiment is the fact that Abcc2-deficient TR– rats show an approximately 6-fold up-regulation of Abcc3 protein in the liver (Johnson et al., 2006). Because Abcc3 is capable of mediating the transport of Ez-gluc (this report), induction of its expression could also be responsible, at least in part, for the reported increase in AUC0–24 h of Ez-gluc in TR– rats. In our experiments using Abcc2(–/–) mice, we observed a 29-fold higher hepatic accumulation of Ez-gluc compared with that in wild-type mice, even though these animals show a 2-fold induction of hepatic Abcc3 protein (Vlaming et al., 2006). This observation shows that, at least in mice, the hepatic expression of Abcc3 protein is not sufficient to markedly divert Ez-gluc from the liver to the blood, underscoring the pivotal role of hepatic Abcc2 protein in clearance of Ez-gluc.
Because ABCG2 is also able to mediate transport of glucuronidated drugs such as estradiol-17β-glucuronide and 4-methylumbelliferone glucuronide (Suzuki et al., 2003), we investigated whether Ez-gluc could also be a substrate for ABCG2. Indeed, transport of a model substrate for ABCG2 could be inhibited dose dependently by Ez-gluc. As discussed above, our data strongly suggest that Abcc2 is the primary determinant for Ez-gluc transport into bile. However, in the absence of Abcc2, Abcg2 may constitute a backup system for transport of this glucuronide. We therefore tested whether mice deficient in both Abcc2 and Abcg2 would have diminished transport of Ez-gluc into bile. Whereas in Abcc2(–/–) mice biliary excretion of Ez-gluc into bile was only marginally (1.8-fold) reduced, a 40-fold reduction of excretion of Ez-gluc in bile was seen in the Abcc2(–/–)/Abcg2(–/–) double-knockout animals. Because the hepatic content of Ez-gluc in Abcc2(–/–) is 29-fold higher than that in wild-type mice, it can be concluded that Abcg2 is also capable of mediating transport of Ez-gluc, albeit with a much lower affinity but with an equally high capacity compared with Abcc2.
In summary, upon glucuronidation in the intestine, the cholesterol-lowering drug ezetimibe undergoes extensive enterohepatic circulation. Based on the results we present here Abcc3, Abcc2, and Abcg2 play a crucial role in this process.
Footnotes
-
This work was supported in part by the Dutch Cancer Society [Grant NKI 2003-2940].
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.026146.
-
ABBREVIATIONS: Ez-gluc, ezetimibe glucuronide; ABC, ATP-binding cassette; AUC, area under the curve; HPLC, high-performance liquid chromatography; E217βG, estradiol-17β-glucuronide; MTX, methotrexate.
- Accepted May 14, 2009.
- Received December 11, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}