


Abstract
The efflux transporter responsible for the canalicular elimination of bile salts from the hepatocytes is the bile salt export pump (BSEP, ABCB11). Absence or inhibition of this transporter leads to bile salt retention in the hepatocyte and in turn can lead to cholestatic liver disease. We expressed the BSEP/Bsep protein from three species (human, rat, and mouse) in a baculovirus-infected Sf9 system. Vesicles prepared from these cells were used to evaluate bile salt transport of four conjugated bile salts. Because the Sf9 system contains less membrane cholesterol than the liver canalicular membrane, the effect of added cholesterol on the kinetics of BSEP/Bsep-mediated bile salt transport was also investigated. Cholesterol treatment increased the Vmax values in all the species, with the most pronounced effect observed in the rat transporter. In contrast, Km values, with the exception of glycochenodeoxycholate, remained largely unchanged. The species-specific bile salt transport inhibition potential of three compounds known to cause clinical cholestasis was investigated in vesicles containing BSEP/Bsep. Troglitazone and glibenclamide inhibited the BSEP/Bsep-mediated transport of different bile salts with similar affinities, whereas the potential of cyclosporine A to inhibit bile salt transport showed species- and bile salt-specific variations. In conclusion, the cholesterol-loaded Sf9 vesicles overexpressing BSEP/Bsep seem to be a useful system for the identification of potential cholestatic compounds and can also be used for the investigation of species specificity. We observed greater differences in IC50 values for inhibitors than in Km values for substrates between species.
Lipophilic drug molecules and metabolites are eliminated through the bile from the human body. The bile contains a highly concentrated solution of bile salts, biliary lipids (phospholipids and cholesterol), and electrolytes. The concentration of these components can be 500 to 1000 times greater in the bile than in hepatocytes. The active transport process mediating canalicular solute transport (which generates bile flow) relies on the activity of efflux transporters in the canalicular membrane, i.e., bile salt export pump (BSEP, ABCB11), multidrug resistance-associated protein (MRP) 2 (ABCC2), multidrug resistance protein (MDR) 3 (ABCB3), and ATP-binding cassette protein subfamily G (ABCG) 5/8 (Arrese and Trauner, 2003; Pauli-Magnus and Meier, 2006). BSEP/Bsep is a member of the ATP-binding cassette (ABC) transporter protein family. It was first cloned in 1998 from rat and identified as the “sister of P-glycoprotein (P-gp),” a name previously proposed for its close similarity in amino acid sequence to P-gp (Gerloff et al., 1998). It is a highly conserved protein; the amino acid similarity between the human and the skate protein is 68% (Cai et al., 2001). The rat and mouse orthologs of the human BSEP are also close relatives, sharing 82 and 80% similarity, respectively (Green et al., 2000; Lecureur et al., 2000). BSEP/Bsep is specialized for transporting monovalent bile salts, such as taurine and glycine conjugates (Stieger et al., 2000; Noe et al., 2001; Byrne et al., 2002), through the canalicular membrane against a concentration gradient in an ATP-dependent manner. The transport of bile salts is saturable and has similar Km values in the low micromolar range for humans, rats, and mice (Stieger et al., 2000; Byrne et al., 2002; Noé et al., 2002; Hayashi et al., 2005).
An important difference among these species is their sensitivity to impairment of BSEP/Bsep transport function (Wang et al., 2001). Mutations of BSEP in humans generally lead to progressive intrahepatic cholestasis that is fatal unless liver transplantation is performed. Absence of BSEP function is termed progressive familial intrahepatic cholestasis type 2 and is manifested clinically by bile salt retention in the liver and plasma and by decreased bile salt concentrations in the bile (Strautnieks et al., 1998; Trauner et al., 1998). The consequences are not as severe in Bsep null mice, the animal model of the disease, probably because of the formation of tetrahydroxy bile salts, which are less toxic than trihydroxy bile salts and which can be excreted into the bile via a non-Bsep pathway (Wang et al., 2001; Perwaiz et al., 2003). Some drugs by inhibiting BSEP produce similar bile salt retention that ultimately leads to severe cholestasis and liver damage. Such drug-induced cholestasis may be considered as an example of ABC transporter-mediated drug/endogenous substrate interactions. Animal models do not always show the severity of the disease seen in humans (Kostrubsky et al., 2003).
The Sf9 system is widely used for the expression of plasma membrane proteins and is an accepted tool for studying ABC transporters. BSEP/Bsep from various species has also been expressed using the baculovirus-infected insect cell system (Stieger et al., 2000; Byrne et al., 2002). The transport characteristics of these proteins were similar to those expressed in the canalicular membrane of the given species (Byrne et al., 2002; Noé et al., 2002). ABC transporters, like ABCB1 (P-gp/MDR1), ABCC2 (MRP2), and ABCG2 (human breast cancer resistance protein), are apically expressed in several physiologically and pharmacologically important barriers, including hepatocytes. These transporters are known to localize in cholesterol-rich microdomains and are sensitive to the cholesterol content of their membrane environment (Bacso et al., 2004; Tietz et al., 2005; Pál et al., 2007; Storch et al., 2007). Along this line, we have previously shown that enrichment of the insect cell membranes with cholesterol can improve the activity of ABCG2/human breast cancer resistance protein (Pál et al., 2007).
Therefore, we hypothesized that cholesterol modulates BSEP/Bsep activity. We report here the effect of added cholesterol on the transport properties of BSEP/Bsep expressed in the Sf9 system using BSEP/Bsep from mouse, rat, and human. We have also examined the effects of three compounds noted earlier to cause clinical cholestasis, namely, cyclosporine A (CsA), troglitazone, and glibenclamide (Stieger et al., 2000; Funk et al., 2001a; Byrne et al., 2002).
Materials and Methods
Materials. [3H]Taurocholate [specific activity (SA) 5.0 Ci/mmol] was purchased from PerkinElmer Life and Analytical Sciences (Waltham, MA). 14C-Glycocholate (SA, 55.0 mCi/mmol) was purchased from Amersham Biosciences (Little Chalfont, Buckinghamshire, UK). 22,23-[3H]Taurochenodeoxycholate (SA, 10.0 Ci/mmol) and [3H]glycochenodeoxycholate (SA, 14.0 Ci/mmol) were synthesized as described by reductive tritiation of the Δ22,23 precursor (Sorscher et al., 1992) and provided by Dr. Alan F. Hofmann (University of California, San Diego, CA). The recombinant baculoviruses encoding wild-type BSEP, mouse Bsep, and rat Bsep and a polyclonal antibody against the C terminus of BSEP, K24 (Noé et al., 2002), were provided by Bruno Stieger (University Hospital, Zürich, Switzerland). Cholesterol complex of RAMEB (cholesterol@RAMEB) was a product of Cyclolab (Budapest, Hungary). Unlabeled taurocholate (TC), glycocholate (GC), taurochenodeoxycholate (TCDC), glycochenodeoxycholate (GCDC), and all the other chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO), unless stated otherwise in the text.
Membrane Preparation and Cholesterol Loading. Insect membrane vesicles were produced using Sf9 insect cells infected with the recombinant baculoviruses listed above. Sf9 cells were cultured and infected with a recombinant baculovirus stock as described earlier (Pál et al., 2007). To prepare cholesterol-loaded membranes, cells were collected and treated for 30 min at 37°C with 10 times the volume of Hanks' balanced salt solution (Invitrogen, Carlsbad, CA) containing 1 mM cholesterol@RAMEB unless indicated otherwise. After 30-min treatment, cells were washed and resuspended in Hanks' balanced salt solution. Purified membrane vesicles from cholesterol-treated and untreated baculovirus-infected Sf9 cells were prepared according to the method described by Pál et al. (2007). Membrane protein content was determined using the bicinchoninic acid method (Pierce Biotechnology, Rockford, IL). Membrane cholesterol content was optimized and determined similarly as described earlier (Pál et al., 2007). Cholesterol-treated membranes are indicated as high activity membranes (HAMs) hereafter.
SDS-Polyacrylamide Gel Electrophoresis, Western Blotting, and Coomassie Blue Staining. The detailed protocol for Western blotting is described elsewhere (Pál et al., 2007). In brief, 6 μg of total protein per sample was loaded per lane. The primary antibody, described under Materials, was used in a 1:500 dilution in blocking buffer, whereas the secondary antibody, sheep anti-rabbit IgG-horseradish peroxidase (HRP), an HRP-conjugated species-specific whole antibody (Sigma-Aldrich), was used in a 1:5000 dilution in 0.05% Tween 20 in phosphate-buffered saline.
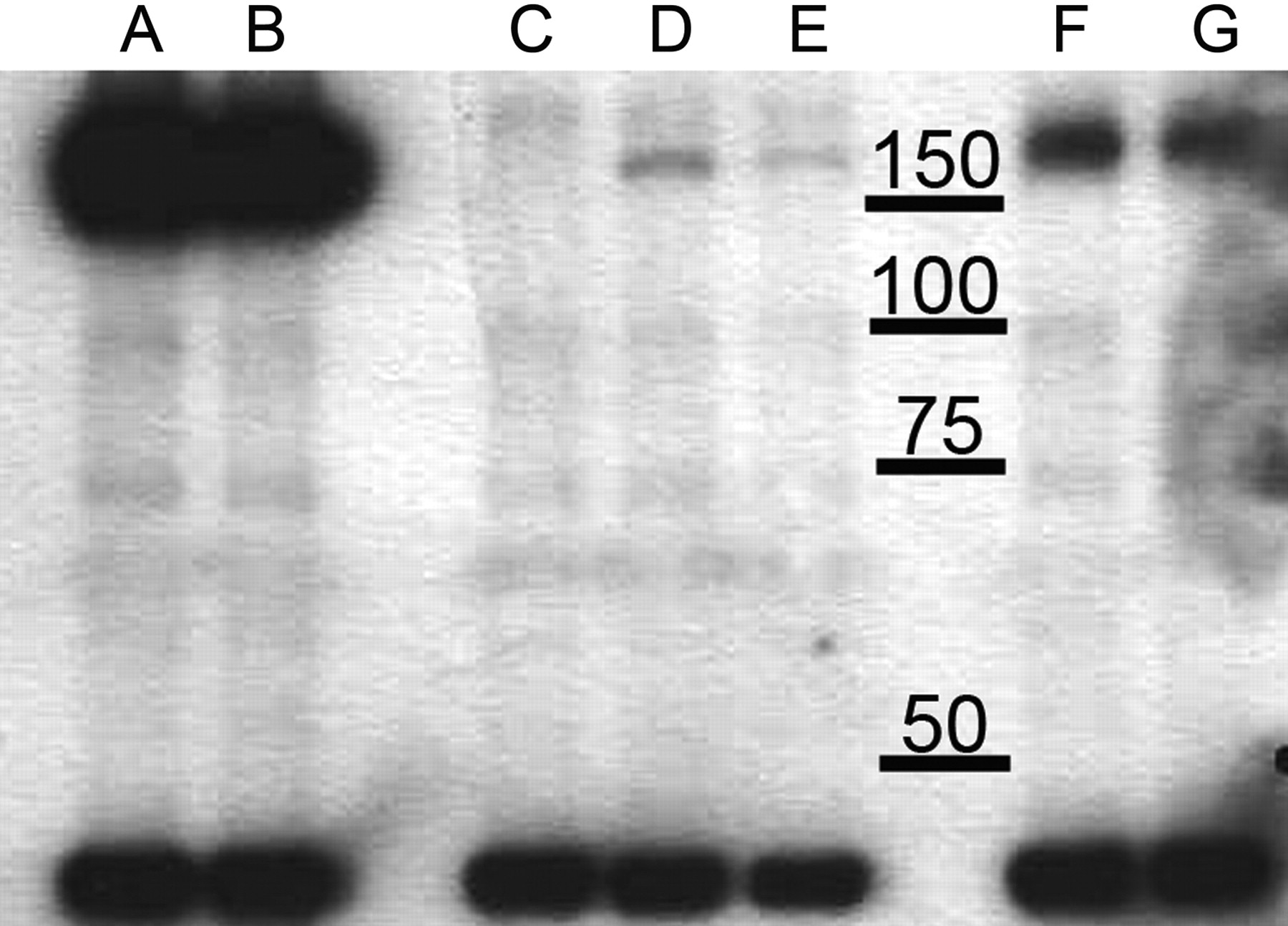
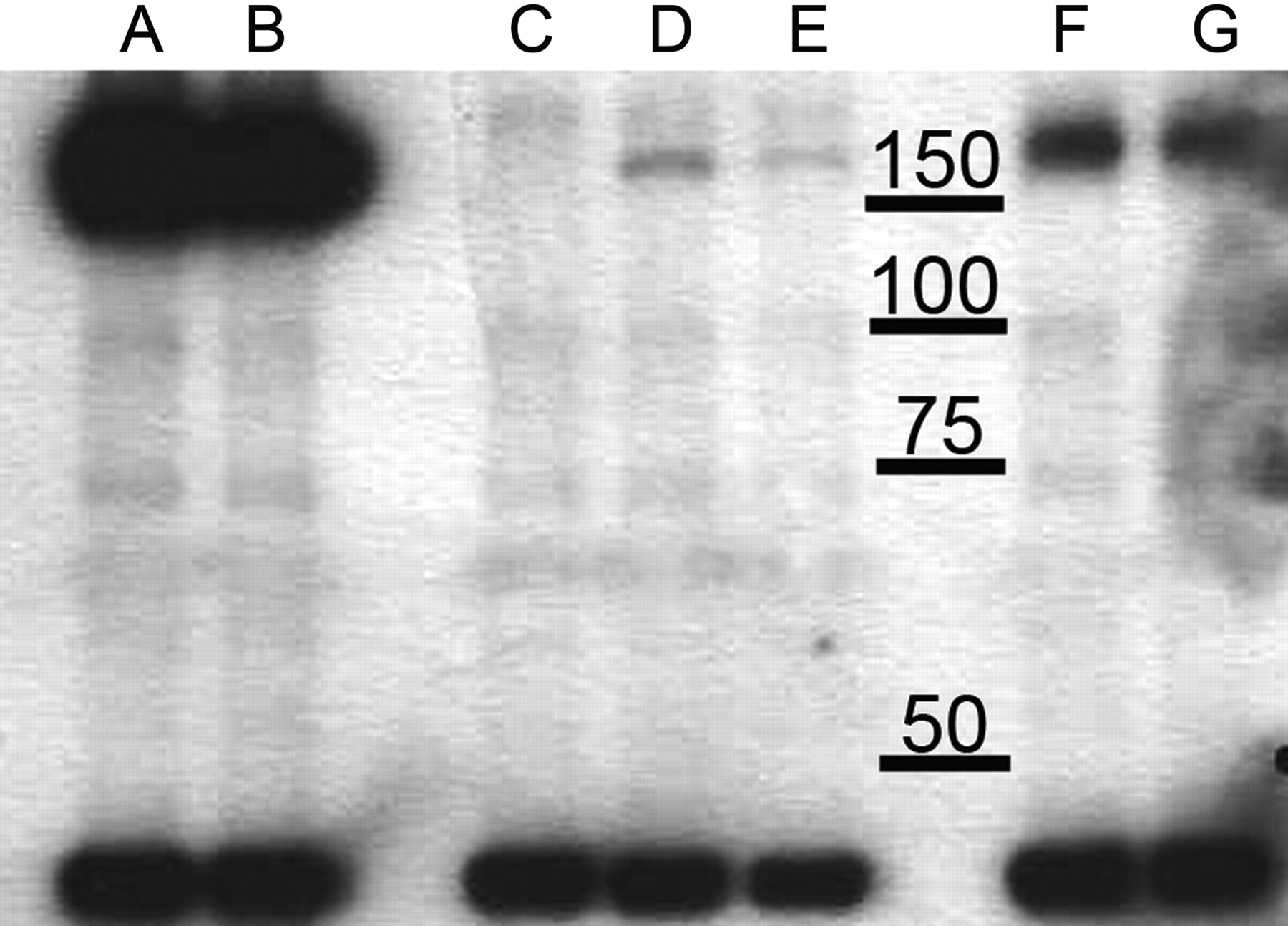
Expression of BSEP/Bsep in Sf9 vesicles treated or untreated with cholesterol. Human BSEP, mouse and rat Bsep, and control β-galactosidase proteins were expressed in Sf-9 cells, and cells indicated as HAM were loaded with cholesterol. Samples collected from the membrane fractions of BSEP-Sf9 (A), BSEP-HAM-Sf9 (B), β-galactosidase containing control vesicles (C), rat Bsep-Sf9 (D), rat Bsep-HAM-Sf9 (E), mouse Bsep-Sf9 (F), and mouse Bsep-HAM-Sf9 (G) were subjected to Western blotting. The band around 150 kDa indicates BSEP/Bsep; the band below 50 kDa indicates α-actin. For detailed description of the method, see under Materials and Methods.
An antibody against α-actin (dilution 1:500) was used as a control for lane-loading discrepancy. The secondary antibody was sheep anti-rabbit IgG-HRP (dilution 1:10,000).
For semiquantitative determination of BSEP/Bsep in the samples, 15 μg (total) of protein was separated on 7.5% polyacrylamide gel and stained with Coomassie Blue. Bovine gamma globulin, a 140-kDa protein (Fermentas UAB, Vilnius, Lithuania), was used as internal standard for the calibration curve (1.25–5 μg/well). For visualization and quantification, ImageJ (National Institutes of Health, Bethesda, MD) software was used.
Vesicular Transport Assay. Inside-out membrane vesicles were incubated in the presence or absence of 4 mM ATP. Incubations were carried out at 37°C for 5 min in 1 M KNO3, 0.1 M Mg(NO3)2, 10 mM Hepes-Tris, pH 7.4, using 50 μg of total protein/well. The bile salts were present in the mixture in 2 μM total concentration (including hot and cold bile salts) unless stated otherwise. The reaction was stopped by the addition of ice-cold wash buffer (100 mM Tris-HCl, 1 M KNO3) with consecutive rapid filtration through Millipore Corporation (Billerica, MA) B-glass fiber filters of a 96-well filter plate. After washing five times with 200 μl of ice-cold wash buffer, the filters were dried, and the retained radioactivity was measured in scintillation mixture (Packard UltimaGold; PerkinElmer Life and Analytical Sciences) using a Wallac MicroBeta TriLux liquid scintillation analyzer (PerkinElmer Life and Analytical Sciences) with ∼50% counting efficiency. ATP-dependent transport was calculated by subtracting the values obtained in the absence of ATP from those in the presence of ATP. Experimental data were evaluated by nonlinear regression analysis using GraphPad Software Inc. (San Diego, CA) Prism version 5.00 as described in detail in a previous report from this laboratory (Pál et al., 2007). Statistical analysis of the effect of cholesterol on bile salt transport by BSEP/Bsep was done using the Wilcoxon test.
Results
Cholesterol Treatment of Sf9 Cells Expressing BSEP/Bsep Does Not Change the Expression of the Protein. Sf9 cells infected with BSEP/Bsep containing baculovirus were treated with cholesterol@RAMEB complex. The optimized treatment increased the cholesterol content approximately 3- to 4-fold compared with untreated membranes, yielding a cholesterol content of 0.16 μmol/mg protein. Vesicles were analyzed for human, mouse, and rat BSEP/Bsep expression using an antibody against the C terminus of BSEP that recognizes all three proteins. Figure 1 shows a representative Western blot picture of the cholesterol-treated and nontreated membranes. Cholesterol treatment had no effect on the size or on the quantity of BSEP/Bsep expressed. Control Sf9 vesicles do not contain Bsep protein. Because the primary antibody recognizes an epitope that is different in the three proteins, we performed Coomassie staining and quantitated BSEP/Bsep using internal calibration standards. The expression of the mouse and the human protein was similar (2.6 μg/15 μg total protein and 2.2 μg/15 μg total protein, respectively), and the rat protein expression was approximately half of that (1.3 μg/15 μg total protein). Cholesterol treatment had no effect on the amount of transporter in the preparations.

Effect of HAM treatment on the BSEP/Bsep-mediated bile salt transport in different species. A dilution series of the bile salts was analyzed in the presence of equivalent amounts of total protein in the control and HAM-treated membranes. Control Sf9 vesicles are indicated by closed circles, whereas HAM-treated vesicles are indicated by closed squares. Transport values are corrected for micrograms of BSEP/Bsep in the preparation. A, human BSEP; B, mouse Bsep; C, rat Bsep. Data are presented as mean ± S.D. of two independent determinations.
Cholesterol Stimulates BSEP/Bsep Activity but Does Not ChangeKm Values. The species-specific transport of four different bile salts was investigated: TC, TCDC, GC, and GCDC. All the bile salts showed saturable transport, following Michaelis-Menten kinetics. Cholesterol-loading increased the Vmax values in all cases (Fig. 2), and the increase in transport was statistically significant in almost all cases. In addition, ATP-dependent transport values were converted to picomole of substrate per microgram of BSEP/Bsep per minute for more proper comparison among species. No such trend was observed with Km values (Fig. 2; Table 1). The bile salt transport of β-galactosidase containing control vesicles was negligible, irrespective of cholesterol treatment (data not shown). Linearization of TC transport by human, mouse, and rat proteins is shown in Fig. 3. No significant change was observed in the Km values as indicated by the parallel curves. The species- and substrate-specific maximal difference in Km values varied around 3-fold in both cholesterol-treated and untreated membranes (Table 1). As shown in Table 1, the species-specific ranking of Km values for bile salts was similar for untreated membranes. GCDC displayed the highest affinity, and GC had the lowest affinity. Only in the case of the rat transporter did cholesterol treatment modulate this order, where treatment increased the uniformity of the Km values (Table 1). The ranking of intrinsic clearance values slightly varies between species (TCDC–GCDC > TC–GC, TC > GCDC–TCDC > GC, and TCDC–GCDC > TC–GC for human, mouse, and rat, respectively); cholesterol loading caused a small change only for rat Bsep (TC > GC–TC–GCDC) (Table 1). Intrinsic clearance values of control and cholesterol-treated vesicles were correlated (Fig. 4). The slopes of the curves were 1.26 ± 0.34, 1.48 ± 0.12, and 2.83 ± 1.34 for human BSEP, mouse Bsep, and rat Bsep, respectively. This indicates that on cholesterol loading the most pronounced effect was in the rat protein, whereas the activity of human BSEP was least affected by the treatment.
Comparison of Km and intrinsic clearance values calculated for BSEP/Bsep
Cholestatic Compounds Inhibit the Transport of Bile Salts to Various Extents.Table 2 summarizes the IC50 values and maximal inhibitory effects obtained from vesicular transport assays. The most pronounced difference within one species was observed in the case of CsA (Fig. 5). The species-specific inhibitory potential of CsA is different for the various bile salts. For human BSEP, there is a 20-fold difference in the IC50 values between GC and TC inhibitions (0.93 and 18.9 μM, respectively). The species-specific inhibitory profiles of troglitazone and glibenclamide were more uniform.
Effect of known cholestatic compounds on BSEP/Bsep-mediated bile salt transport
Species-Specific Bile Salt Transport Is Affected Differently by Cholestatic Compounds. Even though the inhibition profiles of troglitazone (Fig. 6) and glibenclamide (data not shown) are similar within one species, there are species-specific differences in the calculated IC50 values (Table 2). There is approximately an order of magnitude difference in the affinity of glibenclamide for the human and the mouse transporter. IC50 values ranged from 7.6 to 18.8 μM for the human transporter and 95.5 to 153 μM for the mouse transporter (Table 2). The corresponding ranges for troglitazone are 9.5 to 27.6 μM for the human protein and 40.4 to 63.4 μM for the mouse protein. Such a trend could not be observed for CsA. For this compound, the largest interspecies difference in IC50 values was obtained for TCDC, and the smallest was obtained for GC (with ranges of 0.09–7.17 μM and 0.93–2.28 μM, respectively).

Eadie-Hofstee plots for BSEP/Bsep-mediated TC transport. A, human BSEP; B, mouse Bsep; C, rat Bsep. Solid lines represent the nontreated vesicles, whereas dotted lines show HAM vesicles. Data are presented as mean ± S.D. of two independent determinations.
Cholesterol Loading of Sf9 Vesicles Does Not Influence the Kinetic Parameters of BSEP Inhibitors. The effect of nine known BSEP inhibitors (Wang et al., 2003) on TC transport was analyzed with high and low cholesterol-containing Sf9 vesicles expressing human BSEP. All the compounds were able to inhibit BSEP-mediated TC transport both in the control and cholesterol-loaded vesicles. The calculated IC50 data (Table 3) displayed near-identical values for the vesicles with different cholesterol contents. The only exception was CsA, in which a nearly 10-fold difference (2.0 μM in control versus 19.8 μM in HAM vesicles) was observed.
Comparison of the effect of various compounds on TC transport by BSEP in high and low cholesterol membrane environment
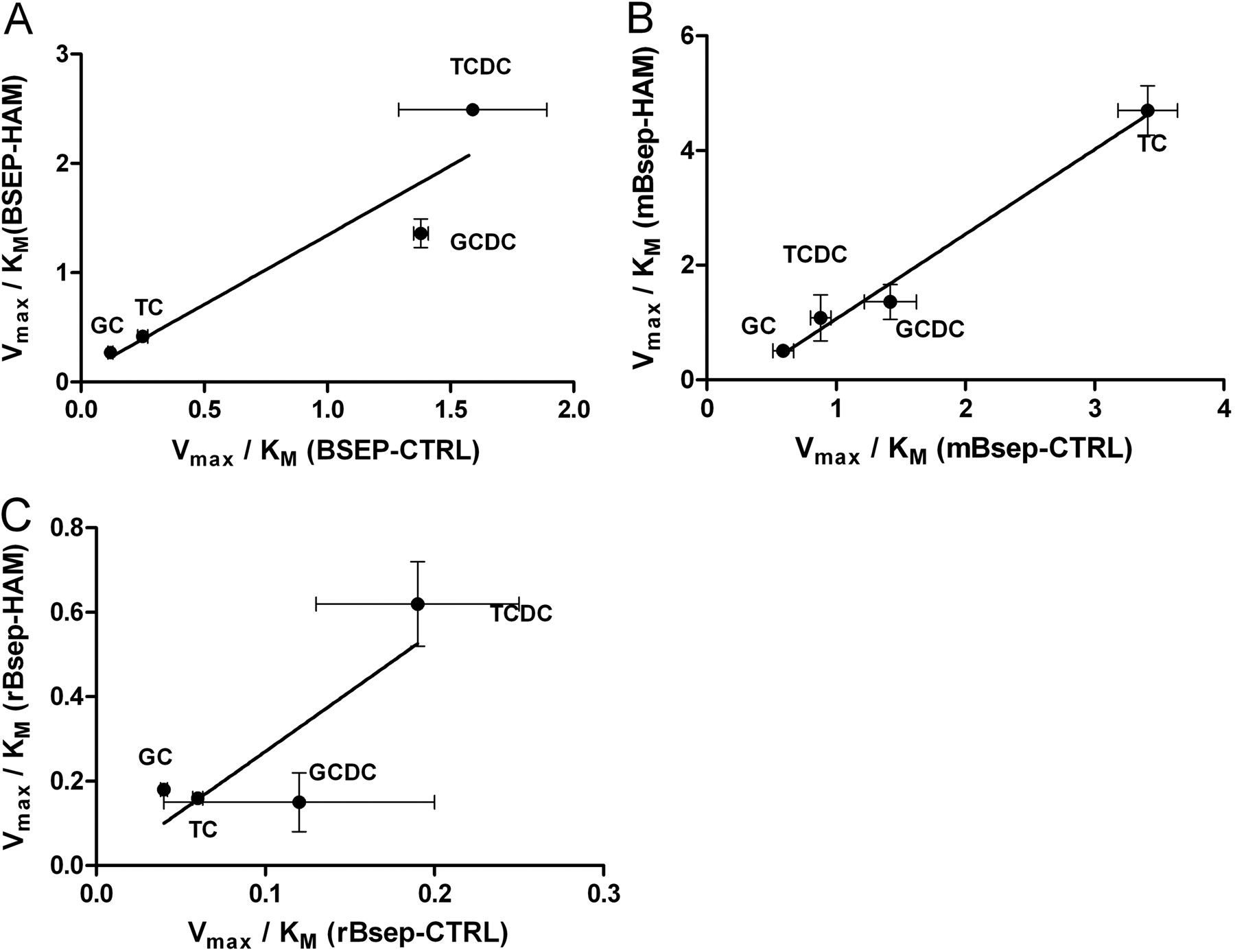
Effect of HAM treatment on the intrinsic clearance of bile salts. Intrinsic clearance values from control and cholesterol-loaded BSEP/Bsep-containing vesicles are compared. Intrinsic clearance values were calculated as stated in the text. The slopes are 1.26 ± 0.34 for human BSEP (A), 1.48 ± 0.12 for mouse Bsep (B), and 2.83 ± 1.34 for rat Bsep (C).
Discussion
Chemical cholestasis is a form of liver toxicity inflicted by drugs. Inhibition of BSEP/Bsep-mediated bile salt transport is now recognized as one of the most common mechanisms for drug-induced cholestasis. Other factors, such as altered metabolism, may reduce the severity of consequences of Bsep function insufficiency. Certain animal species, such as the Bsep-deficient mice (abcb11–/– mice) are still able to secrete bile salts at a rate one third that of the wild-type, presumably by altering bile salt metabolism and thus generating bile salts that can be eliminated via other transporters. Despite the presence of compensatory mechanisms, drug-induced inhibition of Bsep is still considered a major contributor to the development of cholestatic injury. Therefore, measurement of BSEP/Bsep inhibition by candidate drugs is of high importance in the early phases of drug discovery and development.
For studying Bsep-mediated interactions, besides the intricate system of sandwich-cultured hepatocytes, the vesicular transport assay is used most commonly. The first vesicular transport studies used canalicular membrane vesicles (CMVs). The CMV is a popular model (Meier et al., 1984a; Funk et al., 2001a,b; Bode et al., 2002), because this system contains all the relevant canalicular transporters at an expression level close to physiological. In addition, Mrp2 null rat strains (Jansen, 1985; Hosokawa et al., 1992; Paulusma et al., 1996; Ito et al., 1997; Xiong et al., 2000) and Mrp2 (Chu et al., 2006) and Bsep (Wang et al., 2001) knockout mice exist, making transporter specificity studies in rodent CMVs straightforward. However, species specificity studies using CMVs from humans are difficult because of limited access to human samples and the complexity of dealing with polymorphisms. To circumvent this obstacle, Bsep containing insect or mammalian membrane vesicles purified from infected or transfected cells can be used. Insect cells are ideal for large-scale production of membranes; most studies on BSEP/Bsep use this system (Gerloff et al., 1998; Stieger et al., 2000; Byrne et al., 2002; Noé et al., 2002). Inherent limitations, such as improper protein glycosylation and low membrane cholesterol content (Pál et al., 2007), must be taken into consideration. Although underglycosylation does not seem to affect the function of ABC transporters (Bakos et al., 2000; Ozvegy et al., 2002; Pál et al., 2007), membrane cholesterol has been reported to modulate the activity of several ABC transporters localized preferably in cholesterol-rich microdomains of apical membranes (Bacso et al., 2004; Tietz et al., 2005; Storch et al., 2007). Thus, cholesterol is likely to modulate the activity of BSEP/Bsep. Indeed, our data show that cholesterol loading increases the maximal velocity of transport for BSEP/Bsep-mediated conjugated bile salt transport, resulting in increased clearance values for three of four substrates tested. GCDC is the only exception, in which increases in Vmax were compensated by the increases in Km, leading to largely unchanged intrinsic clearance values (Figs. 2 and 3; Table 1). Cholesterol loading had the most pronounced effect on rat Bsep, increasing 3- to 5-fold the intrinsic clearance values for TC, GC, and TCDC (Fig. 4; Table 1).

Effect of CsA on the BSEP/Bsep-mediated transport of bile salts. Comparison of the effect of CsA on the transport of different bile salts using human BSEP (A), mouse Bsep (B), and rat Bsep (C) expressing membrane vesicles. Symbols representing the different bile salts are the following: closed rectangles, TC; closed triangles, GC; closed circles, TCDC; closed diamonds, GCDC. All the bile salts were present in 2 μM concentration. Data are presented as mean ± S.D. of two independent determinations.
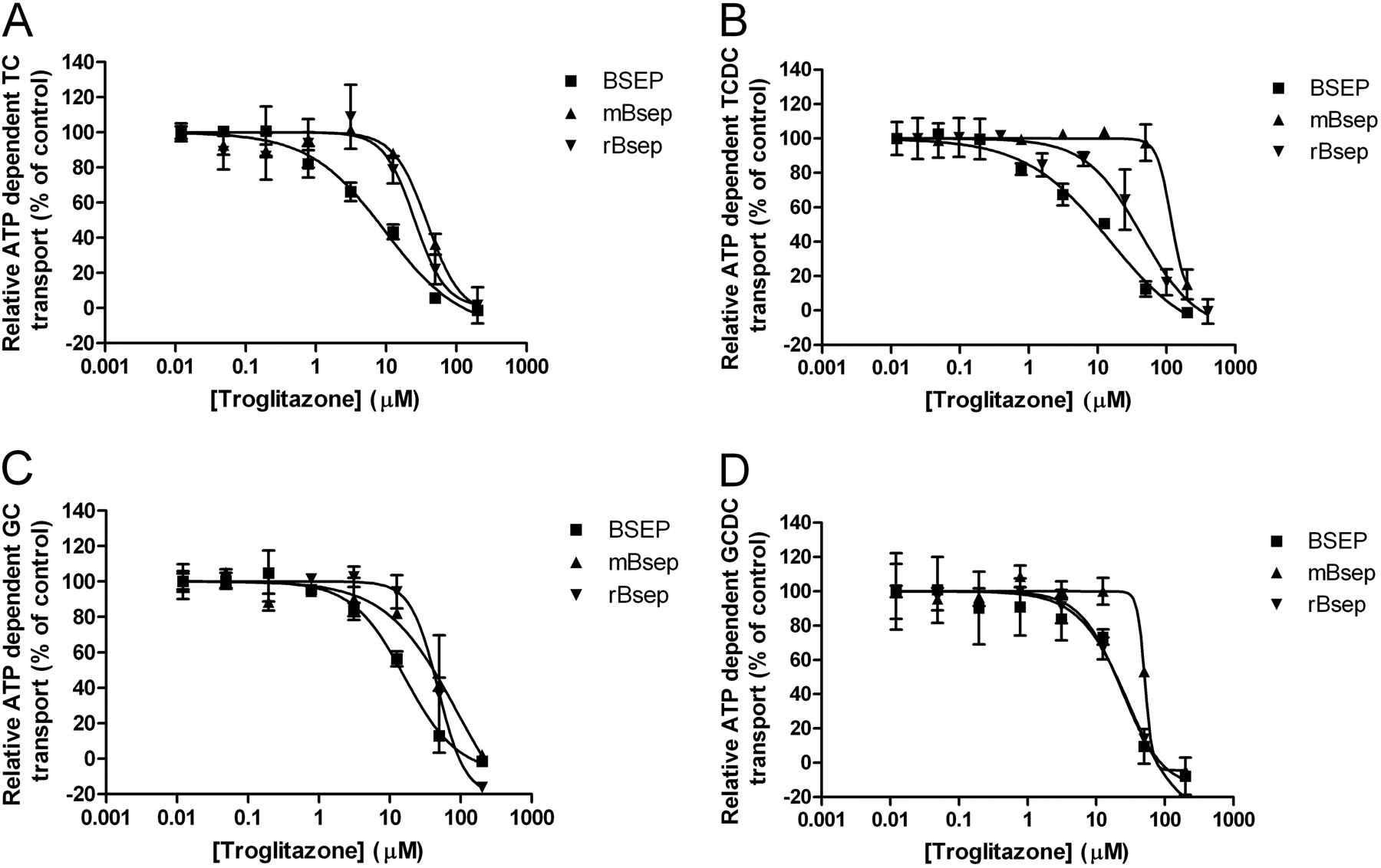
Effect of troglitazone on the BSEP/Bsep-mediated transport of bile salts. Comparison of the effect of troglitazone in the different species on the transport of 2 μM TC (A), TCDC (B), GC (C), and GCDC (D). Data are presented as mean ± S.D. of two independent determinations.
Previous studies using Sf9 membranes overexpressing BSEP/Bsep (Stieger et al., 2000; Noe et al., 2001; Byrne et al., 2002; Noé et al., 2002) reasonably agree in Km values with our results. To illustrate differences typical for this experimental setup, we selected TC as substrate because Km data (4.2, 7.9, and 15.0 μM) were available from three different studies (Byrne et al., 2002; Noé et al., 2002; this study, respectively). In addition, our result is in agreement with the ranking of Km values (TCDC < TC < GC) published earlier using the same baculovirus vectors (Noé et al., 2002). For the cholesterol-loaded membranes (HAM), the Km values correlate reasonably well with data obtained using the similarly cholesterol-rich human embryonic kidney 293 membranes overexpressing BSEP/Bsep (Hayashi et al., 2005). Maximum 2- to 3-fold difference was observed for both the human and the rat transporter.
Because of the use of different expression systems, Vmax values vary from study to study. The maximal velocity depends on the expression level of the transporter that may differ between studies. Because of differences in Vmax in the published data, the intrinsic clearance values also cannot be directly compared.
We tested three inhibitors with four substrates on all three BSEP/Bsep transporters in this novel high-throughput screening system (Figs. 2, 3, 4, 5; Tables 1 and 2). Our data show different patterns for CsA and troglitazone, as well as glibenclamide. Not only did species-specific IC50 values for CsA differ greatly for all the substrates, but also they differed widely between species for the same substrate (Table 2). For troglitazone and glibenclamide there was little deviation within IC50 values for the different substrates within a species. In contrast, there were significant discrepancies between species for the same substrate (Table 2). Glibenclamide and troglitazone displayed the lowest inhibitory potency toward the mouse protein. However, it is interesting to note that for glibenclamide, the IC50 values of the rat transporter are closer to the relevant data on the human transporter. For troglitazone, the IC50 values for the rat protein are closer to the data on the mouse protein (Table 2).
Our results correlate reasonably well with published data. In the case of the human BSEP-mediated TC transport inhibition, the difference between our data and published results is approximately 2-fold for both glibenclamide and CsA (Byrne et al., 2002). For rat Bsep-mediated TC transport inhibition, 3- to 6-fold differences are observed compared with our results (Funk et al., 2001b; Stieger et al., 2002). However, the rankings are the same (CsA < troglitazone–glibenclamide) in all the studies.
To ensure that the obtained IC50 values are valid, we tested a small set of compounds (nine altogether) for their abilities to inhibit BSEP-mediated TC transport in both control and cholesterol-loaded Sf9 vesicles containing human BSEP. The set included the three compounds in focus as well. All the compounds inhibited TC transport with near-identical IC50 values for the respective compounds in the membrane vesicles with different cholesterol content (Table 3). The only outlier was CsA, in which a difference close to 10-fold could be observed. The fact that only this compound was affected by the increased cholesterol content of the system further highlights the special behavior of CsA toward BSEP. Not only was this the compound showing the greatest interspecies and bile salt variability, but also—for yet unknown reasons—this was the one apparently sensitive for the cholesterol content of the membrane.
The cholesterol content of our cholesterol-loaded membrane is 0.16 μmol of cholesterol/mg membrane protein, which is close to data published in the literature for human (0.36 μmol of cholesterol/mg membrane protein) and rat (0.45 μmol of cholesterol/mg membrane protein) canalicular membranes (Ahmed et al., 1995 and Meier et al., 1984b, respectively). The mechanism of cholesterol modulation of BSEP/Bsep activity is unclear. It does not seem to be a high turnover substrate because it does not elevate basal ATPase of mouse Bsep (Kis et al., 2008). A similar pattern (increased Vmax without an effect of similar magnitude on Km) has been observed for ABCB1 (Rothnie et al., 2001) and ABCG2 (Pál et al., 2007). It does not seem to be a consequence of increased expression or more efficient membrane insertion (Fig. 1) or folding. The same effect is observed regardless of treatment protocols (treatment of cells versus treatment of membrane vesicles) (Pál et al., 2007).
Caveolin-1 has been shown to colocalize with ABCB1 (Troost et al., 2004) and coprecipitate with ABCG2 (Storch et al., 2007). In addition, as a result of cholesterol depletion both ABCB1 and caveolin-1 from detergent-resistant membrane fractions decreased in a manner reversible by cholesterol repletion (Troost et al., 2004). It is tempting to speculate that cholesterol promotes ABCB1–caveolin-1 complex formation and activity. However, the effect of caveolin-1 on ABCB1 activity is rather controversial (reviewed in Orlowski et al., 2006). It is also unlikely that cholesterol exerts its effects through the modulation of partitioning into the lipid membranes because changes of membrane cholesterol per se did not cause membrane accumulation or extrusion of substrates (Troost et al., 2004). It is also unlikely that highly hydrophilic substrates would be transported from a binding site embedded in the lipid bilayer (Pál et al., 2007). In sum, it is likely that cholesterol is either an allosteric modulator of ABC transporters or simply acts via changing membrane fluidity.
In summary, we have shown that cholesterol treatment has a marked effect on BSEP/Bsep activity, namely, it increases Vmax values, without producing the same shift in Km values. The significance of our findings is 2-fold. On the one hand, we show that although cholesterol makes BSEP/Bsep work more efficiently, cholesterol loading for the most important substrates does not change the affinity for the transporter, making all data generated on cholesterol-poor native Sf9 membranes still valid. Cholesterol loading did not affect the inhibitor potencies with the exception of CsA. On the other hand, the species specificity of inhibitors (Table 2) is much greater than what would be expected from the moderate interspecies variation in the substrate kinetics (Table 1).
Acknowledgments
We thankÁgnes Potári and Mária Fejes and Dr. Judit Janossy for review and preparation of the manuscript. We also thank Dr. Alan F. Hofmann (University of California, San Diego) for assistance in manuscript presentation.
Footnotes
-
This work was supported in part by Asbóth [Grant XTTPSRT1]; Kozma [Grants TUDAS_06-HTShuMXR and NKFP-1A/041/04]; European Community [Grants BIOSIM-LSHB-CT-2004-005137 and LIINTOP-LSHB-CT-2006-037499]; and Memtrans [Grant LSBH-CT-2006-518246].
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.024778.
-
ABBREVIATIONS: BSEP/Bsep, bile salt export pump, ABCB11; MRP, multidrug resistance-associated protein, ABCC; MDR, multidrug resistance protein, ABCB; ABCG, ATP-binding cassette protein subfamily G; ABC, ATP-binding cassette; P-gp, P-glycoprotein; CsA, cyclosporine A; SA, specific activity; TC, taurocholate; GC, glycocholate; TCDC, taurochenodeoxycholate; GCDC, glycochenodeoxycholate; HAM, high activity membrane; HRP, horseradish peroxidase; CMV, canalicular membrane vesicle.
- Accepted June 10, 2009.
- Received September 22, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}