


Abstract
Recent observations revealed that human UDP-glucuronosyltransferase (UGT) 2B10 catalyzes N-glucuronidation of amine-containing compounds. Knowledge of the substrate specificity and clinical significance of UGT2B10 is still limited. The purpose of this study was to expand the knowledge of UGT2B10 substrates and to evaluate its significance in drug clearance. Using recombinant UGT2B10, we found that it catalyzes the N-glucuronidation of amitriptyline, imipramine, ketotifen, pizotifen, olanzapine, diphenhydramine, tamoxifen, ketoconazole, and midazolam. These are drugs that were previously reported to be substrates for UGT1A4 or UGT1A3, and that contain in their structure either tertiary aliphatic amines, cyclic amines, or an imidazole group. UGT2B10 was inactive in the glucuronidation of desipramine, nortriptyline, carbamazepine, and afloqualone. This group of drugs contains secondary or primary amines, and these results suggest that UGT2B10 preferably conjugates tertiary amines. This preference is partial because UGT2B10 did not conjugate the tertiary cyclic amine in trifluoperazine. Kinetic analyses revealed that the affinity and clearance of UGT2B10 for amitriptyline, imipramine, and diphenhydramine are significantly higher than the corresponding values of UGT1A4 and UGT1A3, although the Vmax values of UGT1A4 toward these drugs are considerably higher. These findings suggest that UGT2B10 plays a major role in the N-glucuronidation of these drugs at therapeutic concentrations. These results are also supported by inhibition studies with nicotine and hecogenin. In conclusion, this study expands the understanding of the substrate specificity of UGT2B10, highlighting its preference for tertiary amines with higher affinities and clearance values than those of UGT1A4 and UGT1A3.
Introduction
UDP-glucuronosyltransferases (UGTs) catalyze glucuronidation of a variety of xenobiotics and endogenous compounds (Tukey and Strassburg, 2000). The 19 functional human UGT enzymes are classified into three subfamilies, UGT1A, UGT2A, and UGT2B. The individual human UGTs show both overlaps and specificity in their substrate preferences. The major reactions that the UGTs catalyze are O- and N-glucuronidations (Chiu and Huskey, 1998). The N-glucuronidation of primary, secondary, and tertiary amines represents an important pathway in the elimination of many drugs, as indicated by the high amounts of N-glucuronides in human urine (Green and Tephly, 1996; Green et al., 1998; Kaivosaari et al., 2011). It is widely recognized that the N-glucuronidation of tertiary amines is catalyzed by UGT1A4 and UGT1A3 (Green and Tephly, 1998). However, because it was difficult to reconcile the observed biphasic kinetics in the N-glucuronidation of some tertiary amines by human liver microsomes (HLMs) with the low affinity that UGT1A4 and UGT1A3 exhibited toward the tested compounds, the possibility that another UGT might be responsible for the high-affinity reaction has been raised (Breyer-Pfaff et al., 2000; Nakajima et al., 2002).
Jin et al. (1993) cloned the cDNA for human UGT2B10, expressed it in COS-7 cells, and tested the enzyme activity in 42 compounds. These compounds mostly contained a hydroxyl group, but one of them did contain a primary amine. Since UGT2B10 did not catalyze the glucuronidation of any of the compounds in this study, it was considered an orphan UGT for more than 10 years. A breakthrough occurred in 2007 with the findings from two different laboratories that UGT2B10 catalyzes the N-glucuronidation of nicotine and cotinine (Chen et al., 2007; Kaivosaari et al., 2007). It was later reported that UGT2B10 also catalyzes the N-glucuronidation of nitrosamines (Chen et al., 2008) and R-medetomidine (Kaivosaari et al., 2008). These UGT2B10 substrates are also glucuronidated by UGT1A4, but at much lower affinity. The latter studies implied a preference of UGT2B10 for tertiary amines and an overlapping substrate specificity with UGT1A4 and UGT1A3 (Chen et al., 2007, 2008; Kaivosaari et al., 2007, 2008). Regardless of these and later studies (Zhou et al., 2010; Erickson-Ridout et al., 2011), the current understanding of the substrate specificity of UGT2B10 is limited, and its clinical significance remains to be clarified. To this end, we have screened 14 drugs containing different amine structures to find whether they are UGT2B10 substrates. Based on these findings, we determined the affinity and clearance for the glucuronidation of three representative substrates by UGT2B10 and compared them with the corresponding values for reactions catalyzed by UGT1A4 and UGT1A3.
Materials and Methods
Materials.
UDP-glucuronic acid (UDPGA), alamethicin, hecogenin acetate, nortriptyline hydrochloride, and β-glucuronidase from Helix pomatia (type H-2) were purchased from Sigma-Aldrich (St. Louis, MO). Amitriptyline, imipramine hydrochloride, ketotifen fumarate, trifluoperazine dihydrochloride, desipramine hydrochloride, carbamazepine, diphenhydramine hydrochloride, and midazolam were from Wako Pure Chemicals (Osaka, Japan). Olanzapine and ketoconazole were purchased from Toronto Research Chemicals (Toronto, Canada). Afloqualone was provided by Mitsubishi Tanabe Pharma (Osaka, Japan). Microsomes from 15 individual human livers (HG03, HG32, HG43, HG64, HG70, HG95, HH1, HH6, HH13, HH18, HH31, HH35, HH40, HH47, and HH89) and pooled HLMs (n = 50) were obtained from BD Gentest (Woburn, MA). Cellfectin Reagent and Spodoptera frugiperda (Sf9) cells were purchased from Invitrogen (Carlsbad, CA). Mouse anti–tetra-His monoclonal antibody was obtained from QIAGEN (Valencia, CA). All other reagents were of the highest grade commercially available.
Construction of Expression Vectors for His-Tagged UGT1A3, UGT1A4, and UGT2B10.
Expression vectors for human UGT1A3 or UGT1A4 containing a His tag (6 histidines) at the C terminus were constructed using the Bac-to-Bac Baculovirus Expression System (Invitrogen). Briefly, the UGT1A3 and UGT1A4 cDNAs were digested from the previously constructed pTargeT/UGT1A3 or pTargeT/UGT1A4 vectors (Fujiwara et al., 2007; Koga et al., 2011) and used to replace the UGT1A1 cDNA in the pFastBac1/His-UGT1A1 vector (Kurkela et al., 2007); this resulted in the pFastBac/UGT1A3 and pFastBac/UGT1A4 vectors. The nucleotide sequences of the constructed plasmids were confirmed by DNA sequencing analyses (Long-Read Tower DNA sequencer; GE Healthcare, Waukesha, WI). The expression vector for human UGT2B10 containing a His tag (6 histidines) (pFastBac/UGT2B10 vector) was previously constructed (Kurkela et al., 2007). These pFastBac1 vectors were transformed into DH10Bac competent cells, followed by transposition of the inserts into bacmid DNA.
Expression of His-UGT1A3, UGT1A4, and UGT2B10 in Sf9 Cells.
Sf9 cells (Invitrogen) were grown in Sf-900 II serum free medium containing 10% fetal bovine serum at 27°C. The recombinant bacmid DNAs were transfected into the Sf9 cells using the Cellfectin Reagent (Invitrogen), and the recombinant baculoviruses were harvested from the cell culture medium 72 hours after transfection. Cells were routinely harvested 5 days after the infection of the recombinant baculoviruses, washed twice with phosphate-buffered saline, and stored at −80°C until analysis. Cell homogenates were prepared by suspension in Tris-buffered saline [25 mM Tris-HCl buffer (pH 7.4), 138 mM NaCl, and 2.7 mM KCl] followed by three cycles of freezing and thawing to disrupt the cells. The suspensions were then homogenized with 10 strokes in a Teflon-glass homogenizer. The protein concentrations of the homogenates were determined according to Bradford (1976).
Glucuronidation Assays.
The typical incubation mixture (200 µl total volume), containing 50 mM Tris-HCl (pH 7.4), 10 mM MgCl2, 25 µg/ml alamethicin, 0.5 mg/ml HLM, or 1 mg/ml cell homogenates containing recombinant human UGT, and test compound was kept on ice for 10 minutes. The compounds and their concentrations, which were set at approximately their Km value in HLM, are shown in Table 1. The mixture was preincubated at 37°C for 3 minutes, and reaction was initiated by the addition of UDPGA (final concentration of 5 mM). After incubation at 37°C for 120 minutes, the reaction was terminated by the addition of 100 µl of ice-cold acetonitrile. Following removal of the protein by centrifugation at 10,000g for 5 minutes, a 20-µl portion of the sample was subjected to liquid chromatography–tandem mass spectrometry (LC-MS/MS) analysis.
Compound concentrations optimized for each compound and the mass/charge (m/z) values to detect the glucuronide in LC-MS/MS analyses
N-Glucuronides Detection by LC-MS/MS.
The LC equipment comprised an HP1100 system including a binary pump, an automatic sampler, and a column oven (Agilent Technologies, Santa Clara, CA), which was equipped with a ZORBAX SB-C18 column (2.1 × 50 mm, 3.5 µm; Agilent Technologies). The column temperature was set at 25°C and the flow rate was 0.2 ml/min. The mobile phase was 0.1% formic acid (A) and methanol including 0.1% formic acid (B). The conditions for elution were as follows: 20% B (0–2.0 minutes), 20%–90% B (2.01–3 minutes), 90% B (3.01–10 minutes), and 20% B (10.01–18 minutes). The LC was connected to a PE Sciex API2000 tandem mass spectrometer (AB Sciex, Framingham, MA) operated in the positive electrospray ionization mode. The turbo gas was maintained at 550°C. Nitrogen was used as the nebulizing, turbo, and curtain gas at 70, 40, and 50 psi, respectively. Parent and/or fragment ions were filtered in the first quadrupole and dissociated in the collision cell using nitrogen as the collision gas. The different transitions selected for each of the compounds are detailed in Table 1. The analytical data were processed using Analyst software (version 1.5; Applied Biosystems, Foster City, CA). Because authentic standards for the glucuronides were not available, we used the peak area values as the index of activity when comparing the three enzymes.
Quantification of Amitriptyline, Imipramine, and Diphenhydramine N-Glucuronides.
High-performance liquid chromatography (HPLC) analyses were performed to quantify the amitriptyline, imipramine, and diphenhydramine N-glucuronides. A 50-µl portion of the sample described earlier was subjected to HPLC analysis. The equipment consisted of an L-2130 pump (Hitachi, Tokyo, Japan), an L-2200 autosampler (Hitachi), an L-2400 UV detector (Hitachi), a D-2000 HPLC System Manager program (Hitachi), and an L-2300 column oven, which was equipped with a Mightysil RP-18GP column (4.6 × 150 mm, 5 µm; Kanto Chemical, Tokyo, Japan). The flow rate was 1 ml/min and the column temperature was 35°C. The mobile phases and the UV wavelength are shown in Table 2. The eluate containing each glucuronide was collected with reference to the retention time. A part of the eluate (200 µl) was incubated with 1000 U/ml β-glucuronidase at 37°C for 24 hours. The reaction was terminated by the addition of 100 µl of ice-cold acetonitrile. After removal of the protein by centrifugation at 10,000g for 5 minutes, a 50-µl aliquot of the sample was subjected to HPLC analysis. Since the conversion of the glucuronide to the parent compound was partial (<50%), the peak area of the remaining glucuronide in the treated sample was compared with that in a control sample that was not treated with β-glucuronidase. The content of the regenerated parent compound in the β-glucuronidase–treated sample could be determined using a standard curve made by the authentic standard. Since the content of the regenerated parent compound should correspond to that of the hydrolyzed glucuronide, we could calculate the content of the glucuronide in the collected eluate. The collected eluate containing the glucuronide, at a known concentration, was used to prepare a standard curve for LC-MS/MS analyses.
The mobile phases and the UV wavelengths in this study
Kinetic Analyses.
Kinetic analyses of amitriptyline, imipramine, and diphenhydramine N-glucuronidation by recombinant UGTs or HLM were performed. The substrate concentration range in each case was 10–1000 µM. The kinetic parameters were estimated from the fitted curve to a Michaelis-Menten, two-enzyme Michaelis-Menten, substrate inhibition, or Hill equation (Bowalgaha et al., 2005) using a computer program (KaleidaGraph; Synergy Software, Reading, PA) designed for nonlinear regression analysis. The kinetic models that best fit to appropriate curves with the highest R2 values were chosen based on the Levenberg-Marquardt algorithm in the KaleidaGraph program. Eadie-Hofstee plots were drawn to visually reveal the suitability of biphasic kinetics in each analysis. For reactions exhibiting Michaelis-Menten kinetics and substrate inhibition, the intrinsic clearance was calculated as Vmax/Km. For reactions exhibiting sigmoidal kinetics, the maximum clearance was calculated as Vmax “ (n - 1)/[S50 ” n(n - 1)1/n] to estimate the highest clearance value (Houston and Kenworthy, 2000). Data are expressed as the means ± S.D. of three independent determinations.
Inhibition Analyses of N-Glucuronidation in HLM.
The concentrations of substrates were determined according to the Km1 and Km2 values in HLM and were as follows: amitriptyline, 20 and 400 µM; imipramine, 50 and 400 µM; and diphenhydramine, 20 and 500 µM. For selective inhibition of UGT1A4 and UGT2B10, we used hecogenin (Uchaipichat et al., 2006; Zhou et al., 2010) and nicotine (Zhou et al., 2010), respectively. Hecogenin (dissolved in dimethylsulfoxide) and nicotine (dissolved in methanol) were added to the incubation mixtures, and their final concentrations were 10 and 500 µM, respectively. The final solvent concentration was 2% (v/v) for both inhibitors; the control incubation mixture contained the same solvent concentration.
Immunoblot Analyses of Recombinant His-Tagged UGTs.
The Sf9 cell homogenates expressing His-tagged UGTs (2–45 µg) were separated on 10% SDS-polyacrylamide gels and electrotransferred onto polyvinylidene difluoride membranes, Immobilon-P (Millipore, Billerica, MA). The membranes were probed with the monoclonal tetra-His antibody (QIAGEN) and then with an IRDye 680LT goat antimouse IgG (LI-COR Biosciences, Lincoln, NE). The band intensity was quantified using the Odyssey infrared imaging system (LI-COR Biosciences).
Statistical Analysis.
Correlation analysis was determined by the Spearman rank correlation method, and P values lower than 0.05 were considered statistically significant.
Results
UGT2B10 Substrates.
To expand the knowledge of the importance of UGT2B10 in drug glucuronidation, we have tested its activity toward drugs that contain different amine structures. Analysis of previous studies implied a preference of UGT2B10 for tertiary amines and substrate specificity overlaps with UGT1A4 and UGT1A3 (Chen et al., 2007, 2008; Kaivosaari et al., 2007, 2008). Therefore, we first chose to examine amitriptyline and imipramine, tricyclic antidepressants that were previously reported to be substrates of UGT1A4 and UGT1A3 (Breyer-Pfaff et al., 1997; Nakajima et al., 2002). We also selected other tricyclic compounds for the current analyses, namely, ketotifen, pizotifen, olanzapine, trifluoperazine, desipramine, nortriptyline, and carbamazepine, of which the first four compounds contain tertiary amines. Ketotifen, olanzapine, and trifluoperazine were reported to be substrates of UGT1A4 (Breyer-Pfaff et al., 2000; Linnet, 2002), but there is no information about which UGT enzyme(s) could catalyze pizotifen glucuronidation. Desipramine and nortriptyline carry secondary amines, and were previously reported not to be glucuronidated by UGT1A4 (Green and Tephly, 1996). Carbamazepine, a substrate of UGT2B7, contains a primary amine (Staines et al., 2004). Afloqualone, diphenhydramine, tamoxifen, ketoconazole, and midazolam were also examined in the present study for glucuronidation by UGT2B10 because these drugs have amine structures and were reported to be substrates for UGT1A4 (Green and Tephly, 1996; Kaku et al., 2004; Kaji and Kume, 2005; Klieber et al., 2008; Bourcier et al., 2010).
The results revealed that UGT2B10 catalyzes the N-glucuronidation of amitriptyline, imipramine, ketotifen, pizotifen, olanzapine, diphenhydramine, tamoxifen, ketoconazole, and midazolam. The structures of these compounds are shown in Fig. 1, along with their conjugation site (indicated by solid black arrows). The tested compounds that were not glucuronidated by UGT2B10 are also shown in Fig. 1. The conjugation sites in the non-UGT2B10 substrates, and the human UGT(s) that conjugates them, are indicated by open arrows. These results suggest that UGT2B10 has a clear preference for tertiary amines over secondary or primary amines. The negative results with trifluoperazine, however, indicate that not every drug with a tertiary amine is a good substrate for UGT2B10 (Fig. 1).
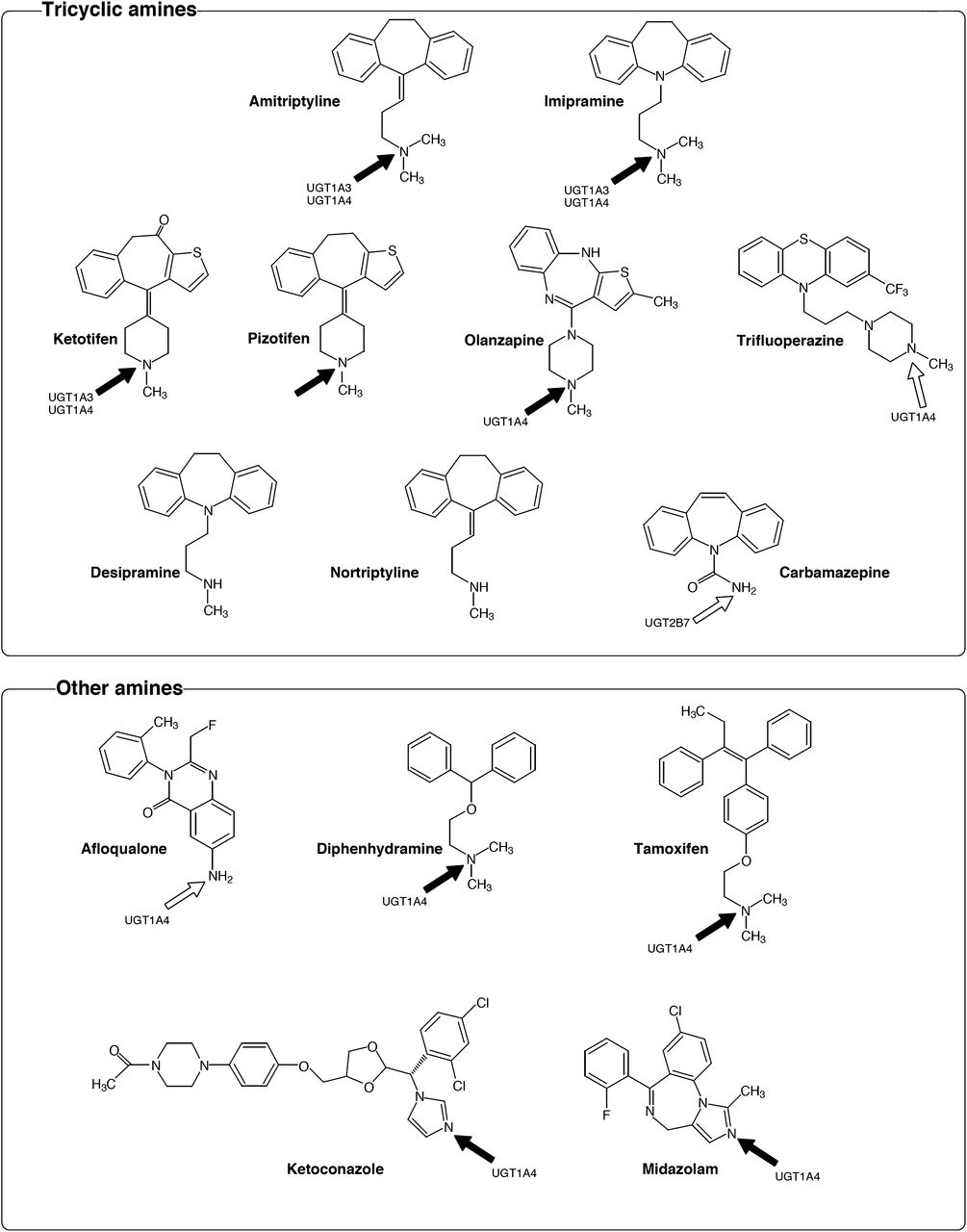
Chemical structures of drugs containing an amine used for the screening of UGT2B10 substrates. Arrows indicate the position of N-glucuronidation. The UGT enzymes, which were previously reported to have this activity, are shown. The compounds with solid arrows were glucuronidated by UGT2B10, whereas the compounds with open arrows were not glucuronidated by UGT2B10.
The nine compounds that were found to be substrates for UGT2B10 were subsequently subjected to activity assays with UGT1A3 and UGT1A4. The substrate concentrations in these experiments are listed in Table 1, and the obtained peak areas are shown in Fig. 2; peak areas are shown because authentic glucuronide standards were not available. In all of these examinations, UGT1A4 exhibited the highest activity, whereas the relative activity of UGT1A3 and UGT2B10 varied between the compounds. It should be noted that comparison of the rate values of the different enzymes without accounting for their relative expression levels might be misleading. Therefore, for the detailed enzyme kinetic studies of the glucuronidation of amitriptyline, imipramine, and diphenhydramine, the relative expression levels of UGT1A3, UGT1A4, and UGT2B10 were determined and used in the calculations.
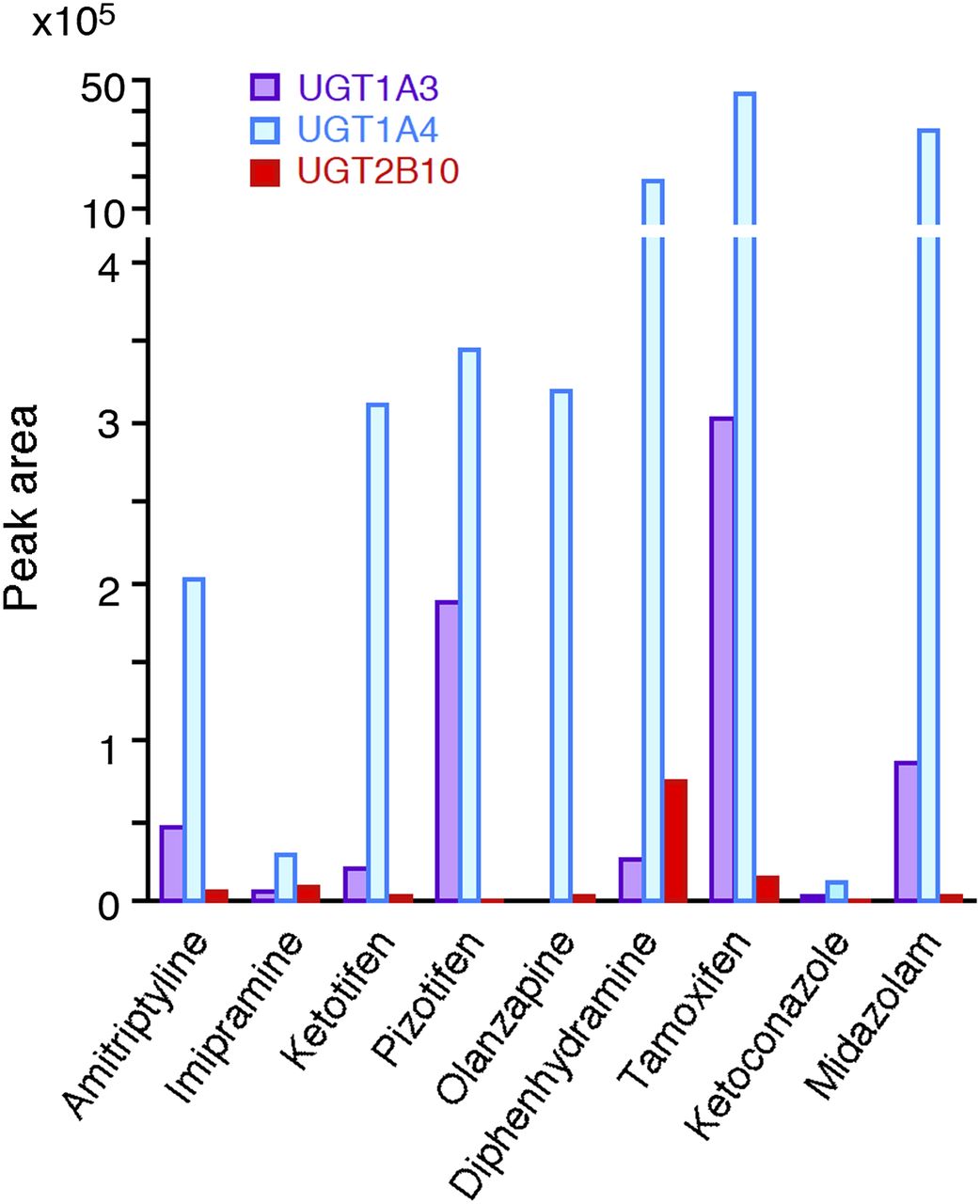
Comparison of the N-glucuronidation of drugs by UGT2B10 with that by UGT1A3 and UGT1A4. Recombinant UGT1A3, UGT1A4, and UGT2B10 (0.5 mg/ml) were incubated with various compounds and UDPGA for 120 minutes. The substrate concentrations are shown in Table 1. Peak area values of the formed N-glucuronide from the LC-MS/MS analysis were used as the indices of activity. Each column represents the mean of duplicate determinations.
Enzyme Kinetics of N-Glucuronidation of Amitriptyline, Imipramine, and Diphenhydramine by recombinant UGT1A3, UGT1A4, and UGT2B10.
We compared the affinities and clearance values of UGT1A3, UGT1A4, and UGT2B10 for the glucuronidation of amitriptyline, imipramine, and diphenhydramine. To normalize the activity with UGT protein expression levels, we performed immunoblot analyses of the recombinant UGTs using a monoclonal antibody toward the C-terminal histidine tag that these recombinant enzymes carry (Fig. 3A). The expression level of UGT2B10, per milligram of protein, was taken as 1.0 and the relative expression levels of UGT1A4 and UGT1A3 were calculated to be 9.4 and 34.3, respectively. Using these relative expression values to normalize activity, the enzyme kinetics of the three UGT enzymes for the glucuronidation of amitriptyline, imipramine, and diphenhydramine were examined (Fig. 3, B–D). For the three tested substrates, UGT1A4 exhibited the highest Vmax values, although relatively high substrate concentrations were required to reach high turnover rates by this enzyme. Amitriptyline and diphenhydramine N-glucuronidation by UGT1A4 followed Michaelis-Menten kinetics, whereas imipramine N-glucuronidation by UGT1A4 fit best to the Hill equation. In the case of UGT1A3, amitriptyline and imipramine N-glucuronidation fit best to the Hill equation, whereas diphenhydramine N-glucuronidation by UGT1A3 was barely detectable, and the activity was too low to calculate any kinetic parameters.
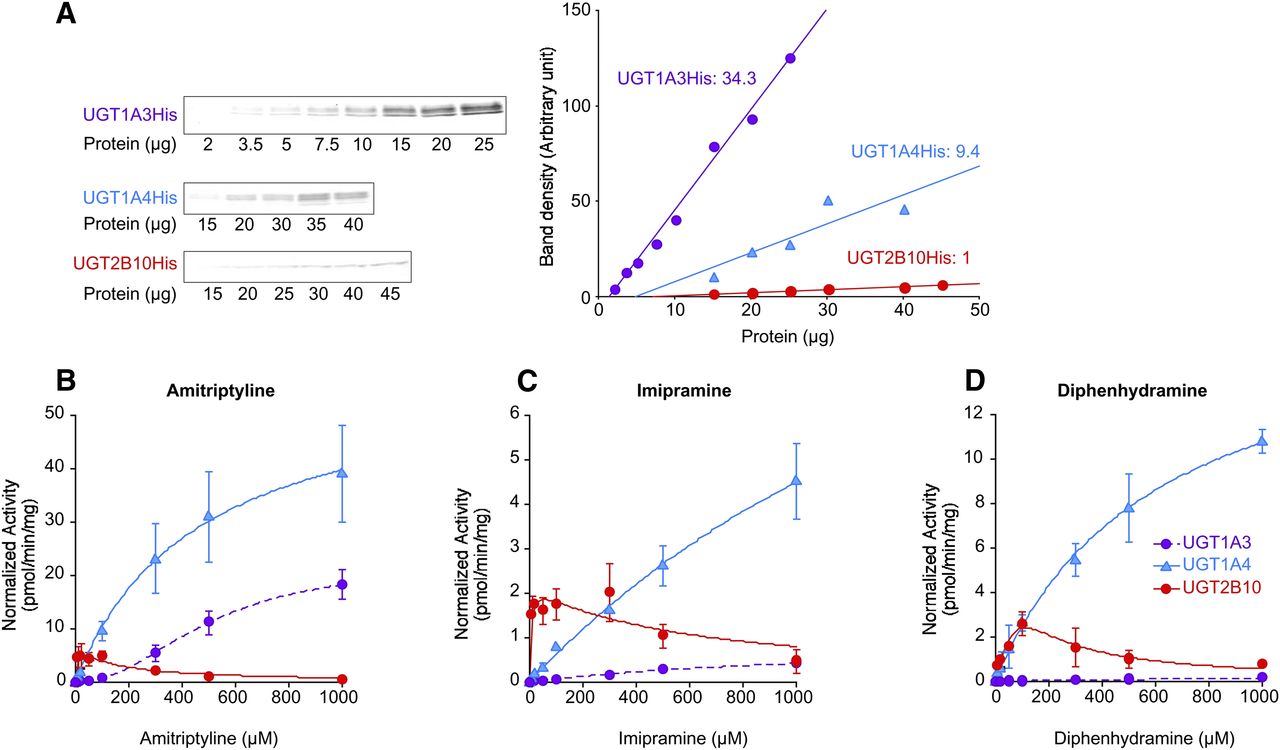
Amitriptyline, imipramine, and diphenhydramine N-glucuronidation by recombinant UGT1A3, UGT1A4, and UGT2B10. (A) Immunoblot analyses of the recombinant UGT1A3, UGT1A4, and UGT2B10 using anti–tetra-His antibody. The relative expression levels of UGT2B10, UGT1A4, and UGT1A3 were calculated as 1, 9.4, and 34.3, respectively, using the calibration curves. Each point represents the mean of duplicate determinations. Kinetics of amitriptyline (B), imipramine (C), and diphenhydramine (D) N-glucuronidation by UGT1A3, UGT1A4, and UGT2B10 using the normalized activities. Data are the mean ± S.D. of three independent determinations.
In contrast to both UGT1A3 and UGT1A4, UGT2B10 exhibited substrate inhibition kinetics for all the three substrates, whereas the diphenhydramine N-glucuronidation was difficult to fit. For this reason, the apparent kinetic parameters for diphenhydramine were estimated only from the results with up to 100 µM, a part of the V versus S curve that fit best to the Michaelis-Menten equation. A clear outcome of the kinetic assays was that, at substrate concentrations below 100 µM, the glucuronidation rates of UGT2B10 were the highest of the three tested enzymes for all of the tested substrates. This result is mainly due to the prominently lower Km values and the higher clearance values of UGT2B10, in comparison with both UGT1A4 and UGT1A3, in the glucuronidation of these three substrates (Table 3).
Kinetic parameters for amitriptyline, imipramine, and diphenhydramine N-glucuronidation by recombinant His-tagged UGT1A3, UGT1A4, and UGT2B10
Kinetics of Amitriptyline, Imipramine, and Diphenhydramine Glucuronidation by HLM.
The results of the kinetic analyses of amitriptyline, imipramine, and diphenhydramine glucuronidation by HLM are shown in Fig. 4. For all three of the tested substrates, the glucuronidation kinetics fit best to the two-enzyme Michaelis-Menten equation. The Eadie-Hofstee plots were biphasic, supporting previous reports for imipramine (Nakajima et al., 2002) and diphenhydramine (Breyer-Pfaff et al., 1997). The Km values for the high-affinity component of these multiple enzyme reactions were close to the Km values that we have determined for UGT2B10, whereas the Km values for the low-affinity component were close to the Km values of UGT1A4 (Table 4).

Kinetics of amitriptyline (A), imipramine (B), and diphenhydramine (C) N-glucuronidation in HLM. An Eadie-Hofstee plot is shown in the inset. Pooled HLMs (0.5 mg/ml) were incubated with 10–1000 μM substrate and 5 mM UDPGA at 37°C for 120 minutes. Data are the mean ± S.D. of three independent determinations.
Inhibition Studies.
To confirm the contribution of UGT2B10 and UGT1A4 to the glucuronidation reactions in HLM at low and high substrate concentrations, respectively, we performed inhibition studies (Fig. 5). Using recombinant UGT1A3, UGT1A4, and UGT2B10, we first confirmed that 10 µM hecogenin selectively inhibited UGT1A4, whereas 500 µM nicotine selectively inhibited UGT2B10 (Fig. 5). Under these assay conditions, we investigated the effects of the two inhibitors on amitriptyline, imipramine, and diphenhydramine glucuronidation by HLM at low and high substrate concentrations. The results demonstrated that hecogenin inhibited the activities by HLM only at high substrate concentrations, whereas nicotine inhibited the activities by HLM only at low substrate concentrations (Fig. 5). These results strongly support the conclusion from the kinetic analyses that UGT2B10 and UGT1A4 are responsible for the high- and low-affinity components in the HLM-catalyzed reactions, respectively.

Effects of hecogenin and nicotine on the amitriptyline (A), imipramine (B), and diphenhydramine (C) N-glucuronidation by recombinant UGTs and HLM. (A) The amitriptyline concentrations for UGT1A3, UGT1A4, UGT2B10, and high- and low-affinity components in HLM were 600, 600, 10, 20, and 400 µM, respectively. (B) The imipramine concentrations for UGT1A3, UGT1A4, UGT2B10, and high- and low-affinity components in HLM were 600, 400, 100, 50, and 400 µM, respectively. (C) The diphenhydramine concentrations for UGT1A4, UGT2B10, and high- and low-affinity components in HLM were 600, 60, 20, and 500 µM, respectively. Data are the mean ± S.D. of three independent determinations.
Correlation Analysis of N-Glucuronidation Using a Panel of 15 HLMs.
To verify the significance of UGT2B10 and UGT1A4 in amitriptyline, imipramine, and diphenhydramine N-glucuronidations by HLM at low and high substrate concentrations, we performed a correlation analysis (Fig. 6). The glucuronidation of the three drugs by 15 individual HLMs was determined at substrate concentrations of 20 and 1000 µM; these concentrations were selected based on the Km or S50 values for the recombinant UGT2B10 and UGT1A4, respectively (Fig. 3, B–D; Table 3). The amitriptyline, imipramine, and diphenhydramine glucuronidation rates at the 20 µM substrate concentration varied from 14 to 51, 36 to 150, and 58 to 210 pmol/min/mg, showing 3.6-, 4.2-, and 3.6-fold variability, respectively. These three glucuronidation rates were significantly (P < 0.001) correlated with each other (r = 0.775, 0.964, and 0.779). The activity rates at the 1000 µM substrate concentration ranged from 110 to 340, 220 to 800, and 580 to 2400 pmol/min/mg, showing 3.1-, 3.6-, and 4.1-fold variability, respectively. These activities were also significantly (P < 0.001) correlated with each other (r = 0.983, 0.946, and 0.911). Interestingly, however, for a given substrate, the correlation between the activities at 20 and 1000 µM substrate concentration was weak, suggesting that the enzymes responsible for the activities were different.

Correlation coefficients between the amitriptyline, imipramine, and diphenhydramine N-glucuronidation in a panel of 15 HLMs. The correlation analyses were performed using the Spearman rank correlation method. The Spearman correlation coefficients (r) are shown. *P < 0.05; **P < 0.01; and ***P < 0.005. Concn, concentration.
Discussion
In this study, we sought to expand knowledge of the substrate specificity and clinical significance of UGT2B10. Starting with aromatic amines, we found that UGT2B10 has the ability to catalyze the N-glucuronidation of amitriptyline, imipramine, ketotifen, pizotifen, olanzapine, diphenhydramine, tamoxifen, ketoconazole, and midazolam. These drugs, except pizotifen, for which there is no report, were previously reported as substrates of other human UGTs, UGT1A4 and/or UGT1A3. Amitriptyline and imipramine were shown by Zhou et al. (2010) also to be substrates of UGT2B10, but their work was unpublished and unknown to us at the start of the current study. A common property among these UGT2B10 substrates is the presence of a tertiary amine. UGT2B10 did not catalyze the N-glucuronidation of carbamazepine, afloqualone, desipramine, and nortriptyline, all of which have a primary or secondary amine. Thus, it appears that, whereas UGT1A4 can metabolize primary and secondary amines (Green and Tephly, 1996; Kaji and Kume, 2005) in addition to tertiary amines, UGT2B10 exhibits a clear preference for tertiary amines. However, the substrate preference of UGT2B10 should presently be regarded as a hypothesis that could be accepted or rejected following studies with yet-untested compounds.
Ketoconazole and midazolam are N-glucuronidated at a tertiary amine within an imidazole ring. The ability of UGT2B10 to conjugate a tertiary amine within an imidazole ring was previously shown for medetomidine (Kaivosaari et al., 2008). It is interesting that UGT2B10 can efficiently form a glucuronide at the tertiary amine within the imidazole ring of R-medetomidine, whereas this enzyme can form two glucuronides in the imidazole ring of S-medetomidine. These reactions include both glucuronidation on a tertiary amine and glucuronidation on a secondary amine. However, these reactions occur at relatively low rates (Kaivosaari et al., 2008). In addition, it was unexpected that UGT2B10 does not glucuronidate trifluoperazine, which has a tertiary cyclic amine and is a typical substrate of UGT1A4. Taken together, our findings may suggest that the length of side chains, and the configuration and spatial orientation of the compound in the active site of the enzyme are also important determinants of UGT2B10 glucuronidation in addition to the chemistry of the amine.
As for carbamazepine and afloqualone, toward which recombinant UGT2B10 showed no activity, we could detect their glucuronides when HLM was used as an enzyme source, revealing the validity of our assays. However, in the cases of desipramine and nortriptyline, we could not detect N-glucuronides even with HLM as the enzyme source. For these compounds, there is currently no report of detected glucuronide in either human plasma or urine. Therefore, it is feasible that desipramine and nortriptyline do not undergo glucuronidation in the human body, or the formed glucuronides are highly unstable.
For the amitriptyline, imipramine, and diphenhydramine N-glucuronidation, the Km values of UGT2B10 were close to those of the high-affinity component in HLM, whereas the Km values of UGT1A4 were close to those of the low-affinity component (Tables 2 and 3). Zhou et al. (2010) reported similar results for amitriptyline and imipramine N-glucuronidation by UGT2B10 and UGT1A4 (Table 4). However, the Vmax and clearance values of UGT2B10 and UGT1A4 cannot be compared because the values determined by Zhou et al. (2010) were calculated as per milligram of protein, without normalization according to relative expression levels (which could not be done in the study by Zhou et al., 2010). The ability to perform such normalization is an advantage of our study system that uses His-tagged recombinant UGTs. Using glucuronidation rate normalization, we found that the clearances of amitriptyline, imipramine, and diphenhydramine by N-glucuronidation of UGT2B10 were higher than those by UGT1A4 by 12-, 57-, and 2.9-fold, respectively.
Kinetic parameters for amitriptyline, imipramine, and diphenhydramine N-glucuronidation in HLM
The reported steady-state plasma concentrations of amitriptyline, imipramine, and diphenhydramine in patients receiving therapeutic doses were approximately 0.2, 0.6, and 0.2 µM, respectively (Bailey and Jatlow, 1976a,b; Blyden et al., 1986). Based on these plasma concentrations and the determined Km values, it appears that UGT2B10 is the major enzyme responsible for their glucuronidation in vivo. We have also tested whether UGT1A1, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B4, UGT2B7, UGT2B15, and UGT2B17 have the ability to catalyze the glucuronidation of these drugs, but found no evidence for such an activity.
UGT2B10 exhibited a prominently higher affinity, for the amitriptyline, imipramine, and diphenhydramine N-glucuronidation, than either UGT1A4 or UGT1A3. This observation is in line with previous studies demonstrating that UGT2B10 has a higher affinity than UGT1A4 for all of the substrates that both enzymes glucuronidate, namely, nicotine (Kaivosaari et al., 2007), clomipramine (Zhou et al., 2010), trimipramine (Zhou et al., 2010), medetomidine (Kaivosaari et al., 2008), and tobacco-specific nitrosamines (Chen et al., 2008). It is expected that this phenomenon will be observed in the other UGT2B10 substrates that were identified in this study, such as ketotifen, pizotifen, olanzapine, tamoxifen, ketoconazole, midazolam, and in UGT2B10 substrates that will be identified in the future.
Previously, we determined the expression levels of all of the UGT genes that are expressed in the human liver (Izukawa et al., 2009) and found that the mRNA of UGT2B10 (19.6% ± 13.4% of all UGTs) was more abundant than the mRNA of UGT1A4 (5.5% ± 3.2%) and UGT1A3 (1.4% ± 1.4%). Although we have to consider the possibility of a poor correlation between mRNA levels and protein expression in UGTs (Izukawa et al., 2009; Oda et al., 2012), the quantitative mRNA data appear to support the significance of UGT2B10 in N-glucuronidation in human livers. In a panel of 25 human liver samples, a huge interindividual variability (506-fold) was observed in the levels of UGT2B10 mRNA (Izukawa et al., 2009). In contrast, the present study showed that the interindividual variability in the amitriptyline, imipramine, and diphenhydramine N-glucuronidation was, at most, 4-fold. This discrepancy might partly be due to the amplifying characteristic of the real-time polymerase chain reaction method. In addition, the possibility of post-transcriptional regulation cannot be ruled out at this stage.
In conclusion, we found that UGT2B10 has the ability to metabolize compounds with tertiary aliphatic amines, cyclic amines, and tertiary amines within an imidazole ring. For amitriptyline, imipramine, and diphenhydramine N-glucuronidation, UGT2B10 exhibited a higher affinity and a higher clearance than UGT1A4. The current results significantly expand the knowledge of UGT2B10 and its substrate specificity. In addition, they demonstrate that UGT2B10 should be ranked highly as an enzyme catalyzing the N-glucuronidation of clinically important drugs.
Authorship Contributions
Participated in research design: Kato, Izukawa, Fukami, Yokoi, Nakajima.
Conducted experiments: Kato, Izukawa, Oda.
Performed data analysis: Kato, Izukawa, Nakajima.
Wrote or contributed to the writing of the manuscript: Kato, Izukawa, Finel, Yokoi, Nakajima.
Footnotes
- Received February 13, 2013.
- Accepted April 23, 2013.
Abbreviations
- HLM
- human liver microsome
- HPLC
- high-performance liquid chromatography
- LC
- liquid chromatography
- MS/MS
- tandem mass spectrometry
- UDPGA
- UDP-glucuronic acid
- UGT
- UDP-glucuronosyltransferase
- Copyright © 2013 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}