


Abstract
The hepatic uptake of glucuronic acid and sulfate conjugates of 6-hydroxy-5,7-dimethyl-2-methylamino-4-(3-pyridylmethyl) benzothiazole (E3040), a dual inhibitor of 5-lipoxygenase and thromboxane A2 synthetase, was investigated in rats. The biliary excretion clearance values for the glucuronide and the sulfate, obtained after i.v. administration of E3040, were similar and corresponded to approximately 30% of the hepatic blood flow rate. The influx clearance values of E3040 conjugates in the presence of 3% bovine serum albumin, measured by a multiple indicator dilution method in the perfused liver, were 1.20 ml/min/g liver for the glucuronide and 0.74 ml/min/g liver for the sulfate, which were twice and equal to the normal hepatic plasma flow rate, respectively, which suggests the presence of an efficient transport system(s). The uptake of E3040 conjugates into the isolated hepatocytes is mediated by Na+-independent active transport system(s), which is inhibited by dibromosulfophthalein and bile acids. The uptake for the sulfate had high-affinity and high-capacity transport activity (Km = 25 μM; V max = 7.8 nmol/min/106 cells) compared with that for the glucuronide (Km = 59 μM; V max = 2.2 nmol/min/106 cells). The uptakes of E3040 conjugates (glucuronide, sulfate) exhibited a mutual competitive inhibition. It is suggested that both conjugates share a multispecific organic anion transporter located on the sinusoidal membrane.
Conjugative metabolism, such as glucuronidation and sulfation, is an important pathway for the inactivation or detoxification of xenobiotics. On the other hand, conjugative metabolites of certain drugs with pharmacologically active (such as the 6-glucuronide of morphine; Osborne et al., 1988) or toxic (such as the glucuronides of anti-inflammatory drugs;Spahn-Langguth and Benet, 1992) properties have been reported. In such cases, the disposition of metabolites, as well as the parent drug, should be considered to identify any pharmacological and/or toxic effect. However, it is difficult to predict the disposition of formed metabolites based on the kinetics of preformed metabolites, because of the uneven distribution of enzymes along the sinusoid, as well as the lower membrane permeability of the metabolites compared with the parent ligands. Pang (1985) and Pang et al. (1992) quantitatively evaluated the kinetics of formed and preformed metabolites after administration of parent ligands and preformed metabolites, respectively. We have also investigated the disposition of conjugative metabolites (Miyauchi et al., 1988; Sato et al., 1986; Shimamura et al., 1993).
In previous studies, we reported the disposition of glucuronide and sulfate of E3040, a novel dual inhibitor of 5-lipoxygenase and thromboxane A2 synthetase, after administration of E3040 byin vivo and by the liver perfusion experiments in rats (Takenaka et al., 1995a,b). We showed that E3040 conjugates were excreted efficiently into the bile. With single-pass steady-state liver perfusion, the hepatic clearance of E3040 was found to be limited by hepatic blood flow, and the formed conjugates were concentratively excreted into bile. Furthermore, studies with bile canalicular membrane vesicles suggested that the biliary excretion of glucuronide across the bile canalicular membrane was mediated by the primary active transport system, whereas that for the sulfate was mediated by another transport system (Takenaka et al., 1995b). This result suggested that the carrier-mediated transport systems across the bile canalicular membrane were one of the reasons for the efficient biliary excretion of E3040 conjugates.
Because we found extrahepatic formation of E3040 conjugates in in vivo experiments, an additional factor for the efficient biliary excretion of E3040 conjugates may be the hepatic uptake mechanism(s) located on the sinusoidal membrane. It is possible that a specific transport mechanism is present for the hepatic uptake of E3040 conjugates, which are organic anions. Cumulative evidence suggests that the nonbile acid organic anions are taken up into the livervia the Na+-independent transport system, which is defined as a multispecific organic anion transporter (Meier, 1988). Substrates for this transport system include bilirubin, bromosulfophthalein, indocyanine green (Laperche et al., 1981; Paumgartner and Reichen, 1976; Scharschmidt et al., 1975; Schwenk et al., 1976; Wolkoff et al., 1987;Yamazaki et al., 1992), β-lactam antibiotics such as benzylpenicillin (Tsuji et al., 1986), the hydroxymethyl glutaryl coenzyme A reductase inhibitor pravastatin (Yamazaki et al., 1993) and DBSP (Blom et al., 1981), although the driving force of this transport system still remains to be clarified (Potter et al., 1987; Wolkoff et al., 1987). The cDNA which encodes the Na+-independent transport system was previously cloned with an expression cloning technique in Xenopus laevis oocytes (Jacquemin et al., 1994).
Many reports have thus been published on the hepatic uptake of nonbile acid organic anions; however, little attention has been given to the hepatic uptake of conjugative xenobiotic metabolites. The uptake and efflux of glucuronide and sulfate of acetaminophen across the plasma membrane of hepatocytes are mediated by carrier-mediated transport system(s) (Iida et al., 1989; Studenberg and Brouwer, 1993). The sulfate conjugates of harmol and 4-methylumbelliferone are transported into hepatocytes via Na+-independent transport systems (Hassen et al., 1996; Sundheimer and Brendel, 1983). In addition, the uptake of glucuronides (harmol glucuronide and glycyrrhizin) are also mediated by active transport systems (Ishida et al., 1993; Sundheimer and Brendel, 1983).
In the present study, we quantitatively evaluated the disposition of E3040 glucuronide and sulfate in rats in in vivo andin vitro experiments. Furthermore, the transport mechanism was also examined with use of the isolated hepatocytes.
Materials and Methods
Materials.
Unlabeled and 14C-labeled E3040 was synthesized in our laboratories (Tsukuba, Japan) (Hibi et al., 1994). The radiochemical purity of [14C]E3040, determined by HPLC, was 98.7%, and the specific activity was 50.9 μCi/μmol. 125I-Labeled BSA was purchased from New England Nuclear Corp. (Boston, MA). DBSP was obtained from Societé d’Etudes et de Recherches Biologiques (Paris, France). Cholate, taurocholate, PCMBS, DIDS, rotenone and BSA (fraction V) were purchased from Sigma Chemical Co. (St Louis, MO). FCCP was purchased from Aldrich Chemical Co. (Milwaukee, WI). The chemical structure of E3040 and its conjugates (glucuronide and sulfate) is shown in figure1.

Plasma concentration-time profiles of E3040 conjugates after i.v. injection to male SD rats. After the i.v. injection of [14C]E3040 glucuronide (2 μCi/0.04 μmol, left panel) or [14C]E3040 sulfate (2 μCi/0.04 μmol, right panel) to male SD rats, blood was collected at specific times to determine the concentration of conjugates by TLC. Each point and vertical bar represents the mean ± S.E. of three different experiments. •, glucuronide; ▪, sulfate.
The glucuronide of [14C]E3040 was prepared by incubating [14C]E3040 in rat liver microsomes. The reaction mixture contained 0.1 M Tris-HCl at pH 7.4, 5 mM MgCl2, 1 mMd-saccharic acid-1,4-lactone, 0.01% (v/v) Triton X-100, 0.2 mg/ml rat liver microsomes and 50 μCi (1 μmol) [14C]E3040 in a final volume of 0.9 ml. The assay was started by adding 0.1 ml of 30 mM UDP-glucuronic acid and incubated for 60 min at 37°C. For the sulfate of [14C]E3040, the reaction mixture contained 0.1 M potassium phosphate buffer (pH 7.4), 3 mM 2-mercaptoethanol, 5 mM MgCl2, 0.2 mg/ml rat liver cytosol and 50 μCi (1 μmol) [14C]E3040 in a final volume of 0.9 ml. The assay was started by adding 0.1 ml of 2 mM 3′-phosphoadenosine-5′-phosphosulfate and incubated for 60 min at 37°C. Both assays were stopped by adding 5 ml CHCl3. After centrifugation the aqueous layer which contained the E3040 conjugates was lyophilized. E3040 conjugates were purified from the lyophilized sample by HPLC. E3040 conjugates labeled with14C were checked for purity by HPLC and confirmed to be more than 99% pure. HPLC analysis was performed on a YMC AM-312 column (C18, 5 μm, 150 mm × 6 mm internal diameter). The mobile phase consisted of MeOH/water/trifluoroacetic acid (100:900:1) (solvent A) and MeOH/water/trifluoroacetic acid (700:300:1) (solvent B). A linear gradient was run from 0 to 30 min to increase the amount of solvent B from 10 to 60%, followed by a 5-min elution with 60% solvent B, and a reverse gradient was run from 35 to 40 min to reduce the solvent B content to 10% again. The chromatographic analysis was performed at a flow rate of 1 ml/min. [14C]E3040 conjugates were identified by means of TLC and HPLC with previously prepared unlabeled E3040 conjugates as standards. The structure of the unlabeled E3040 conjugates was determined by nuclear magnetic resonance and mass spectrometry. Other chemicals used were commercially available and of reagent grade.
Male SD rats (250–330 g) from Japan Laboratory Animals Inc. (Tokyo, Japan) were used.
In vivo study.
The SD rats (n = 3) were lightly anesthetized with diethyl ether, and the femoral artery and vein were cannulated with polyethylene tubing (PE-50) for blood sampling and ligand administration, respectively. The common bile duct was also cannulated with PE-10 to collect bile specimens. Rats were housed in metabolic cages during the in vivo study to obtain arterial blood, bile and spontaneous urine specimens. The rats were allowed to recover from anesthesia before the initiation of the experiment. [14C]E3040 glucuronide or sulfate (2 μCi/0.04 μmol) in saline was injected intravenously through the femoral vein cannula. Blood, bile and urine specimens were obtained at specified times. At each time point, we collected 100 μl of arterial blood.
Analysis of specimens obtained from the in vivo study and the pharmacokinetic analysis.
Plasma, bile and urine specimens were analyzed by TLC as described previously (Takenaka et al., 1995a). For individual rats, the AUC∞ for E3040 conjugates was calculated as the zero-order moment of the time profiles for plasma concentration. Each CLtot of the E3040 conjugates was calculated by dividing the dose by the AUC∞ value. CLbile and CLrenalvalues for E3040 conjugates were calculated by dividing the cumulative amount of E3040 conjugates excreted into the bile and the urine (Xbile and Xurine, respectively) by the corresponding AUC∞ value. CLu,renal was calculated by dividing the CLrenal by the corresponding plasma unbound fraction of each conjugate (the value of glucuronide and sulfate was 0.379 and 0.0613, respectively; Takenaka et al., 1995a).
Estimation of LUR from in vivo experiments.
The LUR, i.e., the fraction of [14C]E3040 conjugates extracted by the liver during a single passage, was measuredin vivo (Liu et al., 1992; Pardridge et al., 1985). The SD rats were lightly anesthetized with diethyl ether, and the hepatic artery was ligated. A 200-μl volume of [3H]inulin (2 μCi), as an extracellular reference, and [14C]E3040 glucuronide or sulfate (0.2 μCi/0.004 μmol) in rat plasma or 3% BSA in Krebs-Ringer bicarbonate buffer (pH 7.4) were injected as a bolus into the portal vein. After 18 sec, the liver was excised, a portion of the liver weighed and the radioactivity counted in a liquid scintillation spectrophotometer (LSC-3500, Aloka Co., Tokyo, Japan). The LUR of E3040 conjugates was obtained by the following equation:
Liver perfusion study (MID method).
All isolated liver procedures were performed as reported previously (Miyauchi et al., 1987). The perfusate consisted of 3% BSA in Krebs-Ringer bicarbonate buffer (pH 7.4), and the flow rate was 30 ml/min. After a stabilization period of approximately 15 min, a 180-μl volume of125I-labeled BSA (0.1 μCi), as an extracellular reference, and [14C]E3040 glucuronide (0.5 μCi/0.01 μmol) or sulfate (0.7 μCi/0.013 μmol) were injected simultaneously as a bolus into the portal vein. Subsequently, the hepatic venous outflow was collected at 1-sec intervals during a 20-sec period with a turntable; then the effluent was collected at 10 sec up to 60 sec. The outflow specimens were analyzed by TLC by the same method as used for the in vivo study. The effluent dilution curves were analyzed based on the distributed model (Bass and Keiding, 1988) by a method reported previously (Miyauchi et al., 1987; Sato et al., 1988; Tsao et al., 1986) to obtain the K inf, the K effand the K seq for total E3040 conjugates. We calculated the PSinf by multiplying theK inf by V ext.V ext is the volume accessible to the extracellular reference during its passage through the liver, which can be estimated by multiplying the flow rate by the mean transit time of125I-labeled BSA (Miyauchi et al., 1987). The ratio of the AUC∞ of E3040 conjugates to that of125I-labeled BSA was defined as the hepatic availability (F), and the extraction ratio was calculated as 1 −F.
Cell preparation.
Hepatocytes were isolated from SD rats (250–330 g) by the procedure of Baur et al. (1975). After isolation, the hepatocytes were suspended (2 × 106cells/ml) at 0°C in albumin-free Krebs-Henseleit buffer supplemented with 12.5 mM HEPES (pH 7.4). Cell viability was routinely checked by the trypan blue (0.4%, wt/vol) exclusion test, and only hepatocytes exhibiting more than 90% viability were used.
Uptake study.
The cell suspension (2 × 106cells/ml) was preincubated in the medium (albumin-free Krebs-Henseleit buffer supplemented with 12.5 mM HEPES, pH 7.4) for 3 min at 37°C. The uptake of [14C]E3040 glucuronide or sulfate was initiated by adding the ligand to the preincubated cell suspension. The final concentration of [14C]E3040 conjugates was 2.5 μM, i.e., the isotope concentration for the conjugates was 0.12 μCi/ml. At designated times, the reaction was terminated by separating the cells from the medium by centrifugal filtration (Yamazaki et al., 1992), and the radioactivity in both cells and medium was determined in a liquid scintillation spectrophotometer (LSC-3500, Aloka Co., Tokyo, Japan). For inhibition studies, the cell suspension was preincubated with either sulfhydryl reagent (100 μM PCMBS), metabolic inhibitors (30 μM rotenone or 2 μM FCCP) or an anion-exchange inhibitor (100 μM DIDS) for 3 min at 37°C before adding [14C]E3040 conjugates. To determine the inhibitory effect of organic anions, either DBSP, unlabeled E3040 conjugate, taurocholate or cholate was added to the cell suspension simultaneously with [14C]E3040 conjugate. The uptake of [14C]E3040 conjugates was corrected for the adherent fluid volume (2.0 μl; Yamazaki et al., 1992). Intracellular space (4.3 μl) was considered when the C/M ratio was calculated. The initial uptake velocity (V 0) was calculated by linear regression on data points taken at 15 and 60 sec.
Determination of kinetic parameters.
The kinetic parameters for E3040 conjugate uptake were estimated by use of the following equation:
To determine the inhibition constant (Ki
) of DBSP and E3040 conjugates on the uptake of [14C]E3040 conjugates, saturable uptake of [14C]E3040 glucuronide and sulfate was examined in the presence and absence of inhibitors. The data were fitted simultaneously to equations 2 and 4 to calculate the kinetic parameters, assuming the competitive inhibition.
Estimation of influx clearance from the in vivoexperiment.
Based on the LUR values of E3040 conjugates obtainedin vivo, the PSu,inf was calculated by use of either the distributed model (Bass, 1980; Bass and Keiding, 1988) or dispersion model (Roberts and Rowland, 1986a; Roberts et al., 1988). The distributed model is:
Results
In vivo study.
Plasma concentration-time profiles of E3040 conjugates after i.v. injection of [14C]E3040 glucuronide or sulfate administered to rats are shown in figure1. Neither glucuronide formed from the sulfate nor sulfate formed from the glucuronide was observed in plasma. The CLtot values of the glucuronide and sulfate were 56.3 ml/min/kg and 19.7 ml/min/kg, respectively (table 1). The cumulative biliary and urinary excretion rates of E3040 conjugates are shown in figure 2. The cumulative amount of glucuronide excreted into the bile up to 3 hr after the i.v. injection of [14C]E3040 glucuronide was approximately 95%. After dosing of [14C]E3040 sulfate, the biliary and urinary excretion rates of the sulfate up to 22 hr were approximately 60 and 21%, respectively (table 1), and an unknown metabolite (M1) was detected in the bile as 7% of the dose (fig. 2). As shown in table 1, the CLbile and CLrenal values of the glucuronide and sulfate were 53.5 and 11.6 ml/min/kg and 2.8 and 3.9 ml/min/kg, respectively. This indicated that the major clearance of E3040 conjugates was by biliary excretion. The CLu,renalcorrected by the plasma unbound fraction was 7.17 ml/min/kg for the glucuronide and 63.8 ml/min/kg for the sulfate (table 1). The CLu,renal of the sulfate was much greater than the glomerular filtration rate in rats (approximately 5 ml/min/kg), which suggested that a secretion mechanism contributes to the urinary excretion of the sulfate.
Disposition of E3040 glucuronide or sulfate in SD rats

Cumulative biliary and urinary excretion of E3040 conjugates in male SD rats. [14C]E3040 glucuronide (2 μCi/0.04 μmol, left panel) or [14C]E3040 sulfate (2 μCi/0.04 μmol, right panel) were injected intravenously as a bolus to male SD rats. Top and bottom panels show the time course of the cumulative biliary and urinary excretion of E3040 conjugates, respectively. Each point and vertical bar represents the mean ± S.E. of three different experiments. •, glucuronide; ▪, sulfate; ⧫, M1.
LUR study.
The LUR calculated by equation 1 is shown in table2. The LUR value of the glucuronide was approximately 0.65 in rat plasma and 3% BSA injected solution, and that of the sulfate was 0.61 in plasma and 0.53 in BSA solution.
The LUR of E3040 conjugates
MID study.
Figure 3 shows the outflow dilution curves of [14C]E3040 glucuronide or sulfate and125I-labeled BSA, used as the extracellular reference. The extraction ratios were 0.31 and 0.13 for the glucuronide and sulfate, respectively (table 3). The dilution curve was analyzed to determine the PSinf, K eff andK seq for total E3040 conjugates and the PSu,inf of E3040 conjugates (table 3). The PSinf value of the glucuronide was 1.20 and that of the sulfate was 0.744. The PSu,inf value of the sulfate was approximately 2.5 times higher than that of the glucuronide (table 3).

Normalized hepatic venous outflow dilution curves of [14C]E3040 conjugates. Rat liver was perfused with erythrocyte-free perfusate for 15 min before a bolus injection of125I-labeled BSA (0.1 μCi) and [14C]E3040 glucuronide (0.5 μCi/0.01 μmol) or 125I-labeled BSA (0.1 μCi) and [14C]E3040 sulfate (0.7 μCi/0.013 μmol) into the portal vein. The total effluent from the hepatic vein was collected at 1-sec intervals over a 20-sec period and then at 10 sec up to 60 sec (left panel, glucuronide; right panel, sulfate). Values on the y-axis are outflow concentrations that have been normalized for the injected dose. Each point and vertical bar represents the mean ± S.E. of five different experiments. •, glucuronide; ▪, sulfate; ○, □, BSA.
MID analysis
The uptake of E3040 conjugates by isolated rat hepatocytes.
The time course of [14C]E3040 glucuronide and sulfate (2.5 μM) uptake by isolated rat hepatocytes is shown in figure4. The C/M ratio at 15 min was 40 for the glucuronide and 200 for the sulfate. Previously, we determined the unbound fraction of E3040 conjugates in the liver by steady-state perfusion of [14C]E3040 (Takenaka et al., 1995b). Because the unbound fraction values in the hepatic cytosol were 0.36 and 0.062 for E3040 glucuronide and sulfate, respectively, the unbound C/M ratio values at 15 min were calculated to be 14.4 and 12.4 for the glucuronide and sulfate, respectively, which indicated the presence of highly concentrative uptake of E3040 conjugates.

Time course for the uptake of [14C]E3040 conjugates (2.5 μM) by isolated rat hepatocytes. • and ▪, the C/M ratio for the glucuronide (left panel) and the sulfate (right panel), respectively. Each point and vertical bar represents the mean ± S.E. from three determinations.
A concentration dependence was observed in the E3040 conjugates uptake, and the Eadie-Hofstee plots of the uptake data indicated the presence of two components (fig. 5). The kinetic parameters are shown in table 4. Comparison of the kinetic parameters for E3040 conjugates revealed that the uptake of the sulfate was composed of a high affinity (1/Km ) and high capacity (V max) transport characteristics compared with the glucuronide (table 4).

Concentration dependence for the uptake of E3040 glucuronide (left panel) and sulfate (right panel) by isolated rat hepatocytes. The isolated hepatocytes were incubated in the medium containing 2.5 μM of [14C]E3040 conjugates with or without unlabeled conjugates. Each point and vertical bar represents the mean ± S.E. from three different preparations. Upper panels, the relationship between the initial uptake velocity (V 0) and the concentration of E3040 conjugates in the incubation medium (S). ———, the least-squares fit of the data to equation 2; … ., the estimated nonspecific diffusion calculated with the value of the nonspecific uptake clearance (P dif); – – –, the theoretical curve of saturable uptake. The same uptake data are shown as Eadie-Hofstee plots in the lower panels.
Kinetic parameters for uptake of E3040 glucuronide or sulfate by isolated rat hepatocytes
The uptake of both conjugates exhibited a marked temperature dependence (fig. 6); the V 0 of the glucuronide decreased to 31% (at 27°C) and 13% (at 0°C) of the control (37°C), and that of the sulfate was reduced to 40% at 27°C and 11% at 0°C. The Q 10 value, which is the ratio of V 0 at 37°C to that at 27°C, was estimated to be 3.2 and 2.5 for the glucuronide and sulfate, respectively. The uptake of E3040 conjugates were also decreased significantly by the sulfhydryl-modifying reagent (PCMBS) and metabolic inhibitors (rotenone and FCCP) (fig. 6). DIDS, an anion-exchange inhibitor, reduced the uptake of E3040 conjugates (fig. 6). Na+ replacement by choline+ had no marked effect on the uptake of E3040 conjugates (fig. 6).

Effect of temperature, sulfhydryl-modifying reagent, ion substitution, metabolic inhibitors and DIDS on the initial uptake of [14C]E3040 glucuronide (left panel, 2.5 μM) or sulfate (right panel, 2.5 μM) by isolated rat hepatocytes. Each bar represents the mean ± S.E. of four determinations in two separate preparations. ** P < .001, * P < .01 by Student’st test.
We also studied the effects of another organic anion, DBSP, and bile acids, taurocholate and cholate, on the uptake of E3040 conjugates. TheV 0 of E3040 conjugates was inhibited by all these compounds in a concentration-dependent manner (fig.7). The uptake of the glucuronide was reduced to 10 to 20% of the control by the presence of these inhibitors (100 μM), whereas the inhibitors had less effect on sulfate uptake (fig. 7). Glucuronide and sulfate also exhibited a mutual inhibition of uptake (fig. 7).

Effect of DBSP, cholate, taurocholate and E3040 conjugates on the initial uptake of [14C]E3040 glucuronide (left panel, 2.5 μM) or sulfate (right panel, 2.5 μM) by isolated rat hepatocytes. Each bar represents the mean ± S.E. of four determinations in two separate preparations.
The inhibition by DBSP and E3040 conjugates on E3040 glucuronide and sulfate uptake was further characterized. The Eadie-Hofstee plots in the presence or absence of these inhibitors are shown in figure8. DBSP (5 μM) and E3040 sulfate (15 μM) were used as inhibitors for E3040 glucuronide uptake, whereas DBSP (5 μM) and E3040 glucuronide (35 μM) were used for E3040 sulfate uptake. Because the nonsaturable component of the uptake of E3040 conjugates may be accounted for by the passive diffusion which was not affected the inhibitor (fig. 8), the kinetic parameters for the uptake of E3040 conjugates in the presence and absence of inhibitors were simultaneously fitted to equations 2 and 4, assuming the competitive inhibition for the saturable component. The calculated curves obtained from least-squares fit of data to the equation coincided with experimental results (fig. 8). It was thus demonstrated that DBSP competitively inhibited E3040 conjugates uptake, and that E3040 glucuronide and sulfate competitively inhibit each other’s uptake. TheKi values of DBSP were 8.4 μM and 5.4 μM for E3040 glucuronide and sulfate uptake, respectively (table5). The Ki value of E3040 sulfate for E3040 glucuronide uptake, and Ki value of E3040 glucuronide for E3040 sulfate uptake were 6.3 μM and 66 μM, respectively (table 5).
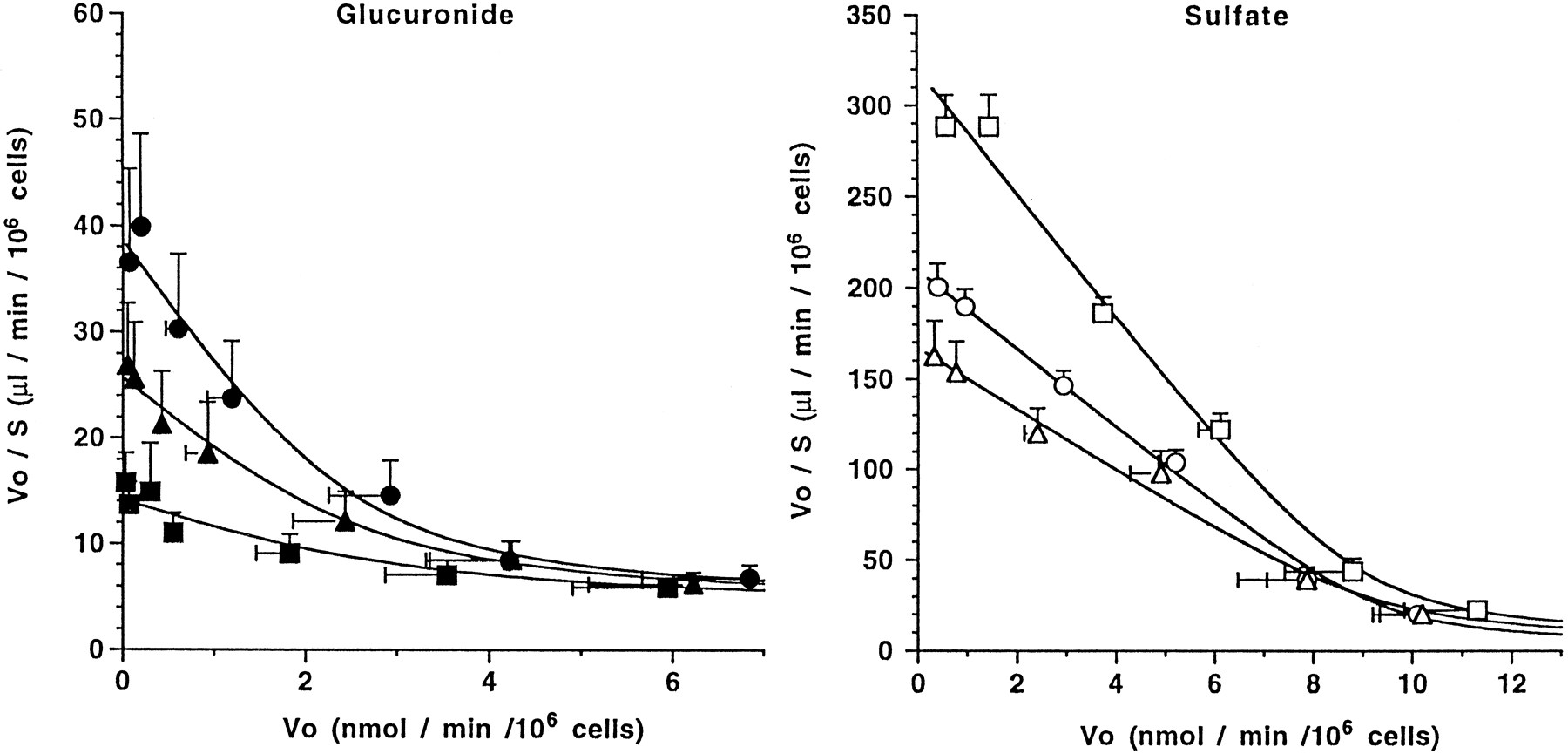
Eadie-Hofstee plots for the uptake of [14C]E3040 glucuronide (left panel) or sulfate (right panel). To characterize the inhibition effect of DBSP on the uptake of [14C]E3040 glucuronide and sulfate, the saturation of [14C]E3040 conjugates was examined in the presence and absence of 5 μM DBSP. In the same manner, the mutual inhibition between E3040 glucuronide and sulfate was examined. Each point and vertical bar represents the mean ± S.E. of four determinations from two independent experiments, both of which were performed in duplicate. ———, the least-squares fit of the data which was obtained by simultaneously fitting the data to equations 2 and 4; left panel, saturation of the uptake of E3040 glucuronide: •, absence of inhibitors; ▴, presence of DBSP (5 μM); ▪, presence of E3040 sulfate (15 μM). Right panel, saturation of the uptake of E3040 sulfate: □, absence of inhibitors; ▵, presence of DBSP (5 μM); ○, presence of E3040 glucuronide (35 μM).
K i values of DBSP and the conjugates for the uptake of E3040 glucuronide and sulfate
Comparison of permeability (PSu,inf) in three different experiments.
The hepatic influx clearance of E3040 conjugates (PSu,inf) obtained from isolated hepatocytes, liver perfusion (MID) and liver in vivo (LUR) are shown in table6. The PSu,inf values of the glucuronide were 2.64, 2.85, 4.19 and 4.92 ml/min/g liver for distributed (DB) or dispersion (DP) model analysis of LUR, MID and isolated hepatocytes, respectively, and these values were similar. The PSu,infvalues of the sulfate were 15.6 (DB), 16.7 (DP) and 11.5 (MID) ml/min/g liver. The PSu,inf value obtained from isolated hepatocytes, however, was 38.4 ml/min/g liver, which was approximately three times higher than those from LUR and MID.
Comparison of PSu,inf of E3040 conjugates determined by different experiments
Discussion
In the present study, we investigated the disposition of E3040 glucuronide and sulfate in rats. The hepatic uptake mechanism of E3040 conjugates was also examined. The CLbile of E3040 conjugates was 54 ml/min/kg for the glucuronide and 12 ml/min/kg for the sulfate; these values accounted for approximately 95% and 62% of the CLtot for the glucuronide and sulfate, respectively (table 1). This result indicates that the main clearance pathway of E3040 conjugates is biliary excretion. Absence of glucuronide or sulfate in plasma, urine and bile after i.v. injection of the sulfate and glucuronide, respectively (figs. 1 and 2 and table 1), suggests that the deconjugation of E3040 may not take place, which was in marked contrast to the conjugative metabolites of 4-methylumbelliferone and harmol (Kauffman et al., 1991; Miyauchi et al., 1989; Sundheimer and Brendel, 1983). Because we found that a demethylated metabolite of E3040 sulfate was present in bile after oral administration of [14C]E3040 to rats and beagle dogs (unpublished data), it is possible that the unknown metabolite, detectable in the bile after i.v. injection of the sulfate, might be the demethylated metabolite of the sulfate (fig. 2). The CLbile value, calculated by considering the partition to red blood cells, for the glucuronide (90 ml/min/kg) was almost the same as the hepatic blood flow rate in rats (60–80 ml/min/kg), and that for the sulfate (19 ml/min/kg) was approximately 30% of the hepatic blood flow rate despite the plasma protein binding of the sulfate being 94%. This indicates that E3040 conjugates are transported efficiently into the liver followed by excretion into the bile. The finding that the CLbile of the preformed and formed conjugates was almost the same after i.v. injection of E3040 conjugates and E3040, respectively, suggests that the specific transport mechanism of E3040 conjugates is also involved in the hepatic uptake process across the sinusoidal membrane (Takenaka et al., 1995a).
To further quantify the hepatic uptake of E3040 conjugates, MID studies were performed. The PSu,inf of E3040 conjugates was estimated with perfused liver after a bolus injection (MID) of E3040 conjugates. As shown in table 3, the hepatic extraction ratios of the glucuronide and sulfate were 0.31 and 0.13, respectively. Because the hepatic extraction of E3040 was previously determined as 0.98 in perfused liver under the same conditions (Takenaka et al., 1995a), the membrane permeability of E3040 conjugates are markedly reduced compared with E3040. The PSinf values of the glucuronide and sulfate, calculated by analysis of the dilution curves (fig. 3) were twice and equal to the hepatic plasma flow rate in rats (0.7 ml/min/g liver), respectively (table 3). Considering the unbound fraction of E3040 conjugates, PSu,inf values of the glucuronide and sulfate were calculated to be 4.19 and 11.5 ml/min/g liver, respectively (table 3). Similar PSu,inf values were obtained in LUR experiments (table 6). It is thus suggested that the hepatic uptake of E3040 conjugates is mediated by specific uptake mechanism(s), such as carrier-mediated active transport system(s), because E3040 conjugates are efficiently taken up into the liver in spite of their hydrophilic nature. We must be cautious in the interpretation of LUR experiments, however, because LUR at 18 sec is a function of not only the PSinf but also the elimination of ligands from the tissue. To accurately determine PSinf from LUR experiments, therefore, the time profiles for LUR values after injection should be analyzed to determine LUR at time zero by extrapolating the profiles. To evaluate the specific uptake mechanism(s) of E3040 conjugates, the uptake by isolated rat hepatocytes was investigated. The uptake characteristics of E3040 glucuronide and sulfate, which are highly concentrative (fig. 4), saturable (fig. 5), temperature dependent (fig. 6) and sensitive to metabolic inhibitors (fig. 6) demonstrated that E3040 conjugate uptake is mediated by energy-dependent uphill transport. The uptake of E3040 conjugates consists of a saturable component and nonspecific diffusion (fig. 5). The uptake affinity (1/Km ) and uptake capacity (V max) of the sulfate was 2.4 times and 3.5 times higher than those of the glucuronide and, consequently, the saturable uptake clearance (V max/Km ) of the sulfate was 8.5 times greater than that of the glucuronide (table 4), which indicates that the intrinsic hepatic uptake ability of the sulfate is higher than that of the glucuronide. The contribution of the carrier-mediated uptake to the total uptake (V max/Km )/(V max/Km + P dif) in a linear range (i.e., at a low concentration of conjugates) is estimated as 89% for the glucuronide and 98% for the sulfate (table 4). Thus, E3040 conjugates are considered to be taken up by the liver predominantly by carrier-mediated mechanism(s).
The E3040 conjugate uptake is Na+-independent, as found for nonbile acid organic anion uptake, and inhibited by the nonbile acid organic anion DBSP and bile acids (taurocholate and cholate) in a concentration-dependent manner (fig. 7). E3040 glucuronide and sulfate also mutually inhibit each other’s uptake (fig. 7). The inhibition of DBSP and E3040 sulfate on E3040 glucuronide uptake and of DBSP and E3040 glucuronide on E3040 sulfate uptake were competitive. TheKi value of DBSP for the uptake of E3040 conjugates (8.4 and 5.4 μM) were similar to theKm value of DBSP itself (Blom et al., 1981), which suggests that E3040 glucuronide, sulfate and DBSP might share the transport carriers (table 5). Furthermore, theKi value of the glucuronide for the sulfate uptake (66 μM) was also similar to the Km itself (73 μM), whereas the Ki of the sulfate for the glucuronide uptake was 6.3 μM, which is smaller than theKm itself (28 μM) (table 5). It is possible that both conjugates may share a common transport carrier. At present, however, we cannot explain the difference between theKi and Km of the sulfate in detail, and we cannot exclude the possibility of noncompetitive inhibition for glucuronide uptake. Taken together, these results suggest that E3040 conjugates may be taken up by hepatocytesvia an organic anion transport system, mediated by the “Na+-independent multispecific organic anion transporter” (Meier, 1988), although the driving force of the uptake mediated by Na+-independent multispecific organic anion transporter has not yet been clearly identified. Furthermore, this system recognizes not only nonbile acid organic anions but also bile acids (taurocholate and cholate) as the substrate (Jacquemin et al., 1994). This result supports our present findings that the uptake of E3040 conjugates is inhibited by bile acids (fig. 7).
Finally, the absolute values of PSu,inf determined from MID studies and those from isolated hepatocytes should be compared. As listed in table 6, the PSu,inf values for the glucuronide estimated from both studies were similar, whereas the PSu,inf value for the sulfate from in vitroexperiments was 2.5 to 3 times higher than that from the perfused liver (table 6); albumin-mediated transport, which was originally proposed byWeisiger et al. (1981), was observed for the sulfate. These results are consistent with our previous results; we determined the PSu,inf values by use of the isolated hepatocytes for several ligands whose degree of protein binding and membrane permeability are significantly different each other, along with the effect of albumin on the uptake of respective ligands in MID experiments (Ichikawa et al., 1992). Among the ligands examined, the albumin-mediated transport was most extensive for warfarin with the lowest fu (0.02) and highest PSu,inf(higher than 140 ml/min/g liver) (Ichikawa et al., 1992). Such phenomena were observed for diazepam, taurocholate, tolbutamide and salycylate whose unbound fraction and PSu,inf were 0.1 to 0.25 and 6 to 140 ml/min/g liver, respectively (Ichikawaet al., 1992). In contrast, the PSu,inf was comparable between the two experimental systems for cefodizime with the highest fu (0.56) and lowest PSu,inf (0.26 ml/min/g liver) (Ichikawa et al., 1992). The discrepancy of PSu,inf for E3040 sulfate between the MID and isolated hepatocyte studies, but not that for E3040 glucuronide, is consistent with the previous results; much higher membrane permeability (38.4 for the sulfate vs. 4.92 for the glucuronide), along with much lower unbound fraction of the sulfate than of the glucuronide (0.06 for the sulfate vs. 0.29 for the glucuronide) could be the cause of the discrepancy.
Because the presence of albumin receptor was denied previously (Weisiger et al., 1984; Weisiger, 1985), one of the possible hypotheses to account for these results is the assumption that the rate-limiting process for the hepatic uptake of ligands with high membrane permeability is the diffusion process through the UWL (Bass and Pond, 1987; Ichikawa et al., 1992; Miyauchi et al., 1993; Weisiger et al., 1989). However, because it is reported that erythrocytes are being squeezed through the sinusoid to erupt UWL (Barry and Diamond, 1984; Holland et al., 1982) and that the UWL effect is more profound under in vitroexperimental conditions (Barry and Diamond, 1984; Bass and Pond, 1987;Blouin et al., 1977), the albumin-mediated transport cannot absolutely be accounted for by this hypothesis. Further, elaborate studies are required to reveal the mechanism for this phenomenon.
In conclusion, E3040 glucuronide and sulfate are efficiently excreted into the bile, and the active transport system for hepatic uptake plays an important role in the efficient biliary excretion of E3040 conjugates. It is suggested that the hepatic uptake of E3040 conjugates is mediated via an Na+-independent multispecific organic anion transport system.
Footnotes
-
Send reprint requests to: Yuichi Sugiyama, Ph.D., Faculty of Pharmaceutical Sciences, The University of Tokyo, Hongo Bunkyo-ku, Tokyo 113, Japan.
- Abbreviations:
- E3040
- 6-hydroxy-5,7-dimethyl-2-methylamino-4-(3-pyridylmethyl) benzothiazole
- [14C]E3040
- [2-14C]6-hydroxy-5,7-dimethyl-2-methylamino-4-(3-pyridylmethyl) benzothiazole dihydrochloride
- SD rats
- Sprague-Dawley rats
- DBSP
- dibromosulfophthalein
- HEPES
- N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid
- MID method
- multiple indicator dilution method
- HPLC
- high-performance liquid chromatography
- BSA
- bovine serum albumin
- TLC
- thin-layer chromatography
- AUC∞
- the area under the plasma concentration-time profiles from zero to infinity
- CLbile
- the biliary excretion clearance
- CLrenal
- the urinary excretion clearance
- CLu,renal
- the unbound urinary excretion clearance
- Xbile
- the amount excreted into the bile
- Xurine
- the amount excreted into the urine
- CLtot
- the total body clearance
- PSinf
- the influx clearance
- PSu,inf
- the unbound influx clearance
- Kinf
- the influx rate constant
- Keff
- the efflux rate constant
- Kseq
- the sequestration rate constant
- Km
- Michaelis constant
- Vmax
- maximal uptake rate
- Pdif
- the nonspecific uptake clearance
- LUR
- the first-pass liver uptake ratio
- UWL
- the unstirred water layer
- PCMBS
- p-chloromercuriphenylsulfonic acid
- DIDS
- 4,4′-diisothiocyanatostilbene-2,2′-disulfonic acid
- FCCP
- carbonyl cyanide-p-(trifluoromethoxy)-phenylhydrazone
- C/M
- cell-to-medium concentration
- Received May 24, 1996.
- Accepted October 30, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}