


Abstract
YM872 {[2,3-dioxo-7-(1H-imidazol-1-yl)-6-nitro-1,2,3,4-tetrahydro-1-quinoxalinyl]-acetic acid monohydrate}, a selective, potent and highly water-soluble competitive α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptor antagonist, was investigated for its neuroprotective effect against focal cerebral ischemia in halothane-anesthetized cats. Cats were subjected to permanent occlusion of the left middle cerebral artery for 6 h, then sacrificed and examined histologically. The electroencephalogram and cerebral blood flow were monitored. Intravenous infusion of YM872 starting 10 min after the onset of ischemia at a rate of 2 mg/kg/h for 6 h markedly reduced the volume of ischemic damage by 61% (from 2604 ± 202 mm3 of the cerebral hemisphere in saline-treated cats to 1025 ± 277 mm3 in YM872-treated cats; P < .01), as assessed in 12 stereotaxically determined coronal sections. No significant differences were observed between YM872- and saline-treated cats concerning physiological variables including brain temperature. No precipitation of YM872 in the kidney was seen in any YM872-treated animal. The present data further support the notion that the AMPA receptor plays an important role in the progression of focal ischemic damage in a gyrencephalic model. This evidence for the neuroprotective efficacy of YM872 suggests its therapeutic potential in the treatment of acute stroke in humans.
The most significant cause of ischemia-induced neuronal damage is the excessive increase of glutamate in the synaptic cleft. Glutamate acts on two distinct subtypes of postsynaptic ionotropic receptors, namely the NMDA and the AMPA receptor subtypes (Watkins et al., 1990). Both these subtypes might be involved in neuronal damage during cerebral ischemic episodes, and in fact considerable evidence demonstrates the neuroprotective effects of NMDA (Park et al., 1988a, b; Bullock et al., 1990; Gill et al., 1991) and AMPA (Gill, 1994; Graham et al., 1996;Lees, 1996; Shimizu-Sasamata et al., 1996) receptor antagonists in experimental focal cerebral ischemia models in rodents and cats. The relative efficacy of these two receptor blockades in focal cerebral ischemia remains to be established, however.
The usefulness of NMDA antagonists as therapeutic agents may be limited by their association with adverse effects, including psychotomimetic effects (Koek et al., 1988), cognitive impairment (Morriset al., 1986) and neurotoxicity (Olney et al., 1989, 1991). Although AMPA antagonists do not share these unfavorable properties of NMDA antagonists (Burchuladze and Rose, 1992; Paradaet al., 1992; Izumisawa et al., 1995; Grahamet al., 1996), the investigation in clinical trials has been limited because first-generation compounds such as NBQX had the major drawback of being poorly soluble in water (Xue et al., 1994).
We recently discovered YM872 (fig. 1), a novel competitive AMPA receptor antagonist, which is a highly water-soluble agent keeping the selectivity and potency for AMPA receptors (Kohara et al., 1996). YM872 has a potent inhibitory effect on [3H]AMPA binding with aKi value of 0.096 μM. In contrast, YM872 has very low affinity for other ionotropic glutamate receptors, with the use of [3H]CGS19755 (NMDA receptor glutamate binding site), [3H]glycine (NMDA receptor glycine binding site) and [3H]kainate (high-affinity kainate binding site) as ligands (Ki = 100 μM, >100 μM, and 2.2 μM, respectively). The solubility of YM872 at pH 7 in Britton-Robinson Buffer is 83 mg/ml, compared with 0.12 mg/ml for NBQX and 0.10 mg/ml for YM90K, which are competitive AMPA antagonists. The in vivo glutamate antagonism of YM872 was assessed through its anticonvulsant effects on audiogenic seizure in DBA/2 mice exposed to ultrasonic sound, in which excessive glutamate transmission is thought to be involved (Meldrum et al., 1983; Chapman et al., 1991; Baron et al., 1992). The minimum effective dose of YM872 after i.p. administration was 3 mg/kg.

Chemical structure of YM872 ([2,3-dioxo-7-(1H-imidazol-1-yl)-6-nitro-1,2,3,4-tetrahydro-1-quinoxalinyl]-acetic acid monohydrate).
The ultimate clinical target (humans) for anti-ischemic drugs is a gyrencephalic species. Data solely from lissencephalic species would be insufficient to allow progress to clinical trials (Bullock et al., 1994). The cat has advantages with respect to its cardiovascular stability under anesthesia and its possession of a gyrencephalic brain. In the present study, we examined the neuroprotective properties of YM872 against neuronal damage induced by focal cerebral ischemia in anesthetized cats in which key physiological variables including brain temperature, EEG and CBF measured by LDF were controlled and monitored throughout the pre- and postischemic period.
Materials and Methods
Animal preparation.
Male ICo: Eur (Tif) cats (IFFA-CREDO, L’arbresle, France) weighing 2.8 to 3.3 kg were used. The animals were maintained on ordinary laboratory chow and tap water ad libitum under a constant 12-h light-dark cycle.
Each cat was anesthetized with ether, tracheotomized and connected to a ventilator delivering a mixture of nitrous oxide (70%) and oxygen (30%) (vol/vol) containing 1 to 1.5% halothane in a closed circuit. Femoral arteries and veins were cannulated bilaterally to monitor arterial blood pressure and obtain arterial blood samples for determination of respiratory status and to administer drugs and obtain blood samples for determination of plasma drug level, respectively. Anesthesia was maintained until the end of investigation. Throughout the experimental period MABP was maintained constant at about 75 mm Hg. The arterial hematocrit was assessed at intervals throughout the experiment to determine the extent of any hemodilution. After immobilization with gallamine triethiodide (1 mg/kg i.v.), normocapnia (PaCO2 approximately 33 mm Hg) was maintained by adjusting the stroke volume of the respirator throughout the course of the investigation. Metabolic acidosis was corrected by administration of sodium hydrogen carbonate (8.4% solution). The animals were maintained at normothermic temperature with a heating pad and lamp, and rectal temperature was monitored. A thermoprobe (29G, Physitemp Instrument Inc., Clifton, NJ) also was inserted into the left parietal cortex to monitor brain temperature.
EEG and LDF.
To record the EEG, screw electrodes were implanted into the skull on the surface of the bilateral middle ectosylvian gyri, where blood is supplied mainly by the MCA. CBF was also measured by LDF (model FLO-N1, Neuroscience Inc., Tokyo, Japan). A small craniectomy (diameter, 3 mm) was made over the left ectosylvian gyrus adjacent to the left screw electrodes. The LDF probe was positioned on the intact dura, and the base-line level of CBF was determined for at least 15 min before occlusion of the MCA (Patelet al., 1995, 1996). Thereafter, CBF was assessed continuously for the duration of the study (6 h) without repositioning of the LDF probe. CBF changes were expressed as a percentage of preocclusion base-line value.
Focal cerebral ischemia model.
The left MCA was occludedvia a modified transorbital approach (O’Brien and Waltz, 1973). The head of the cat was placed in a stereotaxic frame. By use of microsurgical techniques, the left orbit was exenterated, and the orbital roof and optic foramen were removed with a dental drill to expose the dura mater overlying the MCA. Under the surgical microscope (Operation Microscope 700S, Konan Medical Inc., Tokyo, Japan) the dura was incised and the MCA was exposed. After dissection of arachnoid from the MCA, the MCA trunk was occluded with a miniature clip. To standardize the severity of ischemia, only those cats were used in which the mean ipsilateral EEG amplitude decreased by 70% or more of preocclusion level immediately after the onset of ischemia without recovery during the first 10 min of ischemia. Under this condition, our preliminary data indicated that an occlusion time of 6 h provides maximum volume of ischemic damage.
Administration of YM872.
The subsequent selection process for assigning an animal to the saline-versus-YM872 treatment groups was performed in a random manner. YM872 (MW 331.3, free base; Yamanouchi Pharmaceutical Co., Ltd., Tokyo, Japan) was dissolved in isotonic saline alkalized with a few drops of 1 N sodium hydroxide (NaOH) solution to adjust pH to 7.4. YM872 was infused intravenously at a rate of 0.5 mg/3 ml/kg/h, 1 mg/3 ml/kg/h or 2 mg/3 ml/kg/h starting 10 min after the onset of ischemia and continuing for 6 h (n = 8 in each group). Control animals (n = 10) were given an intravenous infusion of saline (3 ml/kg/h) prepared at pH 7.4. In all YM872-treated animals, samples of venous blood were withdrawn at 2-h intervals and centrifuged, and plasma levels of the drug were determined by HPLC. Further, in six (0.5 mg/3 ml/kg/h and 1 mg/3 ml/kg/h) and four (2 mg/3 ml/kg/h) animals, the concentration of YM872 in CSF in the cisterna magna immediately before sacrifice was also determined by HPLC.
Histopathological analysis.
Six hours after occlusion of the MCA, pentobarbital sodium (20 mg/kg) was given intravenously and the cat was sacrificed without reperfusion by transcardiac perfusion fixation with physiological saline followed by 10% formalin neutral buffer solution at pH 7.4. After perfusion the cat was decapitated and the brain removed. The brain was cut into two blocks in the coronal plane through the mammillary body. The anterior part of the brain was cut into four consecutive 4-mm-thick slices in the coronal plane. A 5-mm-thick slice was made by cutting the posterior part of the brain in the coronal plane. Each slice was embedded in paraffin, and sections 7 to 8 μm thick were cut at 200-μm intervals. The sections were stained with hematoxylin and eosin, and 12 stereotaxically predetermined coronal planes were selected according to the coordinates of Snider and Niemer (1961). Neuronal damage was defined based on the following morphological characteristics: microvacuolation, shrinkage of the neuropil and presence of dark neurons and eosinophilic neurons. Neuronal damage in each coronal section was evaluated by a histologist in our laboratory who was blinded to animal treatment. The area of ischemic damage in the cerebral hemisphere, cerebral cortex and striatum was calculated with a computer-aided image analyzer system (Luzex III, Nireco Co., Tokyo, Japan). The volume of ischemic damage was calculated from the area of damage of the 12 coronal sections and their stereotaxic coordinates.
Data analysis.
Data are presented as the mean ± S.E. For cardiovascular, respiratory variables and CBF, statistical comparison among the four experimental groups was performed with two-way repeated measures ANOVA. The volumes of ischemic damage in the four groups were compared with one-way ANOVA followed by Dunnett’s multiple range test. The area of ischemic damage at each coronal plane was compared among the four experimental groups by one-way ANOVA followed by Dunnett’s multiple range test.
Results
Physiological variables and CBF measured by LDF.
There were no significant differences between saline- and YM872-treated cats in the key physiological variables which may affect the amount of ischemic damage, e.g., hematocrit, rectal temperature or MABP, throughout the pre- and postischemic periods (table1, a and b). In saline-treated cats, PaCO2 at the time of MCA occlusion was 31.5 ± 1.1 mm Hg, PaO2 was 148 ± 6 mm Hg and pH was 7.42 ± .02 (table 1a). There were no significant differences in these variables during the succeeding 6-h period, nor between saline- and YM872-treated cats at any time point (table 1a). In the present study, brain temperature was monitored to examine whether or not the neuroprotective effect of YM872 was caused by its lowering effect on the brain temperature. Both in saline-treated and in YM872-treated cats brain temperature shifted in parallel with rectal temperature and there were no significant differences between cats treated with saline and those treated with YM872 (1 or 2 mg/kg/h), in which the volume of ischemic damage was significantly reduced (table1b).
Physiological variables and CBF before and after treatment with YM8721-a
There were no significant differences in CBF measured by LDF between saline- and YM872-treated cats throughout the postischemic period (table 1b). Occlusion of the left MCA led to an immediate drop in CBF to 31 ± 3% of the preocclusion base-line value in saline-treated cats, and to 31 ± 7% (0.5 mg/kg/h), 28 ± 4% (1 mg/kg/h) and 28 ± 3% (2 mg/kg/h) of the base-line value in YM872-treated cats (table 1b). In saline-treated cats, CBF remained depressed at this level throughout the rest of the experiment (29 ± 4% at 6 h after MCA occlusion), whereas in YM872-treated cats it exhibited modest, but not significant, increases at the end of the 6-h observation period (table 1b).
Neuroprotective effect of YM872 on ischemic damage.
Intravenous infusion of YM872 starting 10 min after MCA occlusion at a rate of 1 or 2 mg/kg/h significantly reduced the volume of ischemic damage in the cerebral hemisphere and cortex in a dose-dependent manner (fig. 2). In particular, YM872 at a rate of 2 mg/kg/h reduced the volume of ischemic damage by 61% in the cerebral hemisphere (from 2604 ± 202 mm3 in saline-treated cats to 1025 ± 277 mm3 in YM872-treated cats) and by 63% in the cerebral cortex (from 2418 ± 191 mm3 in saline-treated cats to 897 ± 258 mm3 in YM872-treated cats) (fig. 2). In contrast, YM872 showed no significant protective effect on ischemic damage in the striatum (fig. 2). In the cerebral hemisphere and cortex, YM872 reduced the area of ischemic damage in all coronal sections, with this reduction being statistically significant in coronal sections at 1 to 19 mm stereotaxic distance (2 mg/kg/h) (fig.3). The area of ischemic damage in the striatum was not ameliorated by treatment with YM872 (fig. 3).

Neuroprotective effect of YM872 on the volume of ischemic damage in the cerebral hemisphere (Hemisphere), cerebral cortex (Cortex) and striatum (Striatum) at 6 h after MCA occlusion in cats. Each value represents the mean ± S.E. (n = 10 in saline-treated group,n = 8 in each YM872-treated group). Saline or YM872 was intravenously infused starting 10 min after the onset of ischemia and continuing for 6 h. *P < .05, **P < .01 for comparison between saline- and YM872-treated cats (one-way ANOVA followed by Dunnett’s multiple range test).

Area of ischemic damage in the cerebral hemisphere (A, hemisphere), cerebral cortex (B, cortex) and striatum (C, striatum) at 12 defined coronal sections in saline-treated (control) and YM872-treated cats. Each value represents the mean ± S.E. (n = 10 in saline-treated group,n = 8 in each YM872-treated group). Saline or YM872 was intravenously infused starting 10 min after the onset of ischemia and continuing for 6 h. *P < .05, **P < .01 for comparison between saline- and YM872-treated cats (one-way ANOVA followed by Dunnett’s multiple range test).
Plasma levels of YM872 (0.5, 1 and 2 mg/kg/h) increased continuously, reaching 1.27 ± 0.14 μg/ml, 3.18 ± 0.27 μg/ml and 9.40 ± 1.38 μg/ml, respectively, at the end of the experiments (fig. 4). CSF levels of YM872 were 12.7 ± 2.0 ng/ml (0.5 mg/kg/h), 23.4 ± 1.0 ng/ml (1 mg/kg/h) and 71.2 ± 15.2 ng/ml (2 mg/kg/h), which indicates that the CSF/plasma ratio was approximately 1:100.
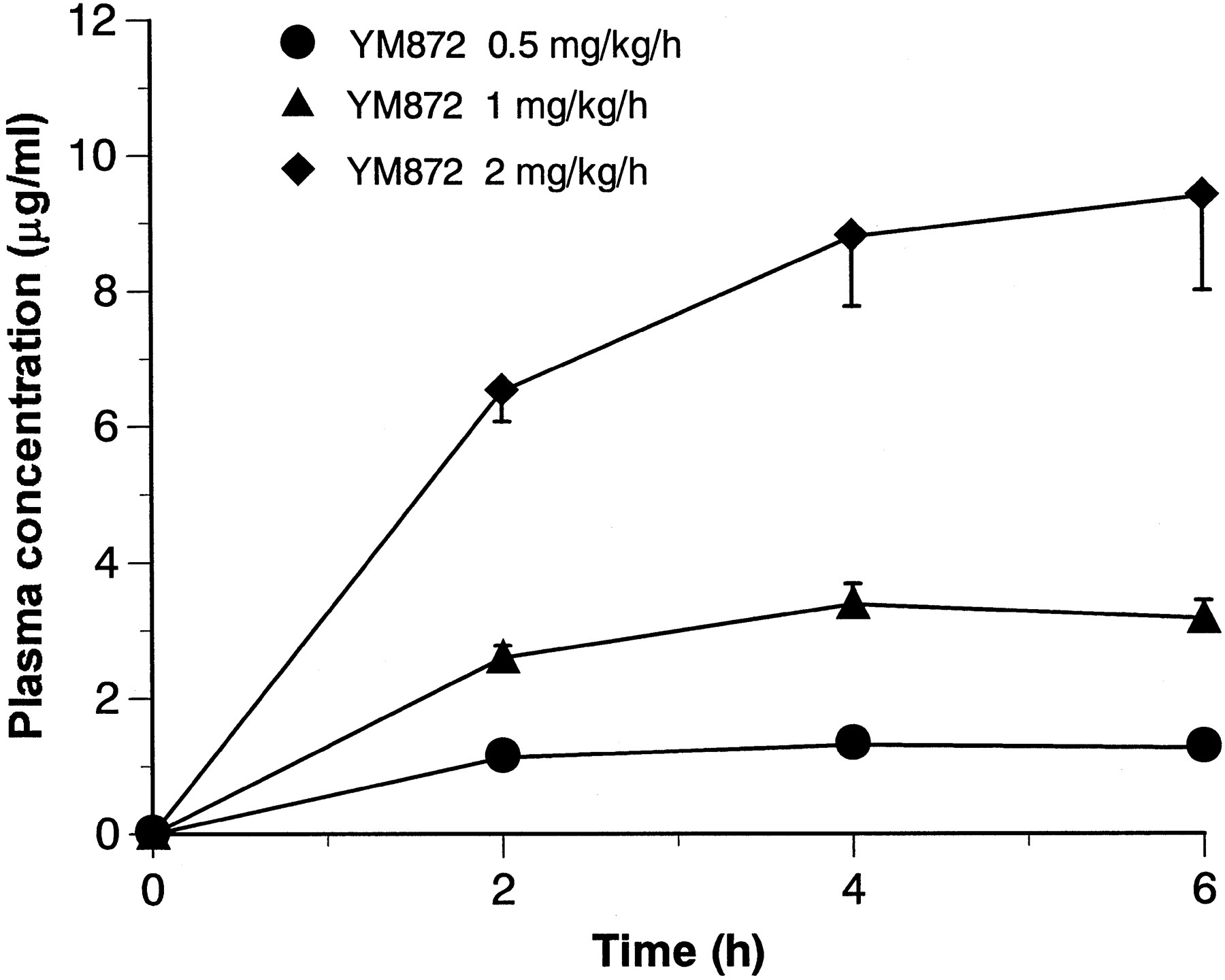
Plasma levels of YM872 during the postischemic period (MCA occlusion at time 0). YM872 was infused intravenously starting 10 min after the onset of ischemia and continuing for 6 h. Each value represents the mean ± S.E. (n = 8 in each group).
Neither drug precipitation nor associated tissue damage was observed in the kidney of YM872-treated animals under light microscopy.
In preliminary studies, the effect of YM872 on the behavior of normal cats was observed. At a rate of 0.5 mg/kg/h, YM872 showed no obvious effect on behavior, but at 2 mg/kg/h, it induced sleep (data not shown).
Discussion
Most ischemic episodes in humans occur as a result of occlusion(s) of the MCA or one of its penetrating branches (Karpiak et al., 1989). In the cat the transorbital approach produces consistent infarction (although of variable infarct size) with commensurate neurological deficits that are analogous to the condition seen after stroke in humans (Bose et al., 1984; Berkelbachet al., 1988; Karpiak et al., 1989). In the present study the MCA was occluded via this method. The Eur (Tif) cats used in this study were genetically more homogeneous than conventional mongrel animals. In addition, to standardize the severity of ischemia, only those cats in which mean ipsilateral EEG amplitude decreased by 70% or more of preocclusion level immediately after the onset of ischemia without recovery during the first 10 min of ischemia were used. Sixty-three percent of the cats in this study satisfied this condition. It has been reported that this change in EEG amplitude is a good index for predicting ischemic damage in MCA-occluded cats and this protocol provides relatively larger and more uniform infarcts than those of other methods (Uematsu et al., 1989a; Yatsugiet al., 1996). These findings indicate that our experimental conditions are suitable for the evaluation of neuroprotective agents against focal cerebral ischemia in cats.
The results of the present study demonstrated that the AMPA antagonist YM872 markedly reduced the volume of ischemic damage in a focal cerebral ischemia model using a gyrencephalic species. This finding confirms previous work with other AMPA antagonists and contributes several new aspects. The relative neuroprotective potency of AMPAversus NMDA receptor blockade in focal cerebral ischemia remains to be fully established. Various studies on the efficacy of antagonists of these two subtypes in focal cerebral ischemia models in rodents (Gill et al., 1991, 1992; Buchan et al., 1992; Shimizu-Sasamata et al., 1996) and cats (Park et al., 1988a; Bullock et al., 1990, 1994;Yatsugi et al., 1996) have shown that the neuroprotective effects of AMPA antagonists are relatively modest compared with those of NMDA antagonists. For example, in cat MCA occlusion models in which the histological end point of 6 h after the onset of ischemia was commonly used, AMPA antagonists LY-293558 and YM90K reduced the volume of ischemic damage in the cerebral hemisphere by only 18% and 38%, respectively (Bullock et al., 1994; Yatsugi et al., 1996), whereas NMDA antagonists MK-801, remacemide hydrochloride and d-CPP-ene reduced it by 50%, 49% and 65%, respectively (Park et al., 1988a; Ozyurt et al., 1988; Bullock et al., 1990; Bannan et al., 1994). In contrast, our present results demonstrated that YM872 administered after ischemic insult reduces the volume of ischemic damage in the cerebral hemisphere by 61%. This finding suggests that the neuroprotective effects of AMPA antagonists in focal cerebral ischemia in gyrencephalic species are equipotent to NMDA antagonists. Although NMDA antagonists have the above-mentioned neuroprotective potency, it is well known that they produce such adverse effects as psychotomimetic effects (Koek et al., 1988), cognitive impairment (Morris et al., 1986) and neurotoxicity (Olneyet al., 1989, 1991) in experimental models and clinical trials (Dorman et al., 1996), which limits their clinical usefulness. However, AMPA antagonists do not share these unfavorable properties of NMDA antagonists (Parada et al., 1992;Izumisawa et al., 1995; Graham et al., 1996; Liet al., 1996). The neuroprotective potency of YM872 therefore suggests the advantage of AMPA antagonists over NMDA antagonists in clinical use.
The reportedly low water solubility of first-generation compounds such as NBQX has led to concerns about drug precipitation in the renal tubules (Xue et al., 1994). This characteristic may have limited doses for experimental and clinical trials of these compounds, which may itself explain why AMPA antagonists have shown relatively modest neuroprotective effects in animal focal cerebral ischemia models. Improving the solubility of AMPA antagonists is thus an important matter. YM872, an N1 carboxymethylated derivative of YM90K, is a significantly more water-soluble compound than YM90K and NBQX (Kohara et al., 1996), and in the present study markedly reduced the volume of ischemic damage by 61% at a dose of 2 mg/kg/h, at which no precipitation in the kidney was observed. To our knowledge, this is the greatest neuroprotective efficacy by an AMPA antagonist in a cat MCA occlusion model. Administration of YM872 at a rate of 2 mg/kg/h for 6 h starting after occlusion of MCA produced a CSF concentration of 71.2 ng/ml (0.22 μM), at which the drug in vitro selectively blocks AMPA receptors without blocking NMDA receptors (Kohara et al., 1996). This suggests that the permeability of YM872 into the brain is sufficient for the exertion of its neuroprotective effect and that YM872 is active as a selective antagonist of AMPA receptors in vivo.
The present data provide evidence that the neuroprotective effect of AMPA antagonists in focal cerebral ischemia models cannot be attributed to drug-induced or artifactual alterations in key physiological variables, such as MABP, blood gas and brain temperature. The monitoring and control of physiological variables which may affect neuronal damage is essential in the evaluation of putative neuroprotective agents (Ginsberg and Busto, 1989). In particular, because hypothermia during or after ischemia may ameliorate ischemic injury (Busto et al., 1989; Kuroiwa et al., 1990;Welsh et al., 1990; Xue et al., 1992), both brain and rectal temperature were monitored directly throughout the pre- and postischemic period. Neither temperature was influenced by administration of YM872, which indicates that its neuroprotective effect is not caused by any decrease in brain temperature. Although AMPA antagonists such as NBQX have been reported to cause a significant decrease in temperature (Nurse and Corbett, 1993, 1996), YM872 in preliminary studies did not induce hypothermia in normal nonanesthetized cats at doses which cause sedative effects, nor in MCA-occluded rats at doses at which it exerts its neuroprotective effects (data not shown). The difference is now under investigation. As a AMPA antagonist which does not induce hypothermia, YM872 may be a useful tool in evaluating the pathophysiological role of the AMPA receptor in ischemic stroke. CBF measured by LDF in YM872-treated cats exhibited a modest but not a significant increase at the end of the 6-h observation period as compared with cats given saline. This may be a result of histological improvement, because in another series of experiments in normal cats intravenous infusion of YM872 at 2 mg/kg/h did not affect CBF measured by LDF in the ectosylvian gyri (data not shown). As in saline-treated cats, EEGs in YM872-treated cats were not restored during the ischemic period. This may be because of the sedative effect of YM872, as demonstrated in preliminary studies in which YM872 given to normal cats at a rate of 2 mg/kg/h induced sleep.
The AMPA receptor is the primary postsynaptic excitatory receptor in brain. Thus, blockade of the AMPA receptor prevents neuronal depolarization induced by excessive extracellular glutamate. It is well known that the elevation of intracellular Ca++concentration after excessive synaptic release of glutamate plays a critical role in ischemia-induced neuronal damage (Siesjö, 1981;Uematsu et al., 1989a). This elevation of intracellular Ca++ concentration is mediated by many pathways: 1) the NMDA receptor, which is modulated voltage dependently by Mg++ under resting conditions (Wong and Kemp, 1991); 2) VSCCs on postsynaptic membranes (Miller, 1991); 3) reversal of Na+-Ca++ exchanger (Siesjo and Bengtsson, 1989); and 4) AMPA receptor lacking the GluR2 subunit, which is also permeable to Ca++(Hollmann et al., 1991; Verdoorn et al., 1991). It is possible that the accumulation of toxic levels of Ca++ results from Ca++entry through a combination of all these routes and that blockade of only one provides at least some degree of protection. In fact, an NMDA antagonist and VSCC antagonist reduced ischemic damage in the cat MCA model, and also attenuated the elevation of intracellular Ca++ concentration (Uematsu et al., 1989b, 1991). Blocking of the AMPA receptor by antagonists could prevent opening of the NMDA receptor channel and could also attenuate the activation of VSCCs after depolarization of the cell membrane. This would prevent Ca++ entry into neuronsvia these two pathways. In addition, AMPA antagonists may also prevent Ca++ entry by reversal of the Na+-Ca++ exchanger and through AMPA receptors lacking the GluR2 subunit. Such a decrease in Ca++ entry through these pathways by blockade of the AMPA receptor may be one neuroprotective mechanism of AMPA antagonists such as YM872 in focal cerebral ischemia.
In conclusion, we have demonstrated that YM872, a selective and potent AMPA antagonist with improved water solubility, markedly reduced neuronal damage in focal cerebral ischemia in cats. The present data further support the notion that the AMPA receptor plays an important role in the progression of focal ischemic damage in a gyrencephalic model. YM872, a highly water-soluble compound, may be a useful tool in investigating the pathophysiological role of the AMPA receptor in ischemic stroke without concern about hypothermia and nephrotoxicity. The efficacy of YM872 in gyrencephalic species suggests its therapeutic potential in the treatment of acute stroke in humans.
Acknowledgment
The authors thank Drs. K. Oikawa and A. Miyashita at the Institute for Drug Development Research for their determination of plasma and CSF levels of YM872, and Dr. T. Hanada at the Institute for Drug Development Research for his preparation of brain slices. The authors also thank Drs. T. Mase and S. Usuda at the Institute for Drug Discovery Research for their invaluable advice.
Footnotes
-
Send reprint requests to: Masayasu Takahashi, M.S., Neuroscience Research, Pharmacology Laboratories, Institute for Drug Discovery Research, Yamanouchi Pharmaceutical Co., Ltd., 21 Miyukigaoka, Tsukuba, Ibaraki 305, Japan.
- Abbreviations:
- AMPA
- α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid
- CBF
- cerebral blood flow
- CGS19755
- cis-4-phosphonomethyl-2-piperidine carboxylic acid
- CSF
- cerebrospinal fluid
- d-CPP-ene
- d-(E)-4-(3-phosphonoprop-2-enyl)piperazine-2-carboxilic acid
- EEG
- electroencephalogram
- HPLC
- high-performance liquid chromatography
- LDF
- laser Doppler flowmetry
- LY-293558
- (3SR,4aRS,6RS,8aRS)-6-[2-(1H-tetrazol-5-yl)ethyl] decahydroisoquinoline-3-carboxylic acid
- MCA
- middle cerebral artery
- NBQX
- 2,3-dihydroxy-6-nitro-7-sulfamoyl-benzo (F) quinoxaline
- NMDA
- N-methyl-d-aspartate
- VSCC
- voltage-sensitive calcium channel
- YM90K
- [6-(1H-imidazol-1-yl)-7-nitro-2, 3(1H, 4H)-quinoxalinedione monohydrochloride
- MABP
- mean arterial blood pressure
- ANOVA
- analysis of variance
- Received February 12, 1997.
- Accepted October 27, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}