


Abstract
Cyclosporin A (CyA), prednisolone (Pred) and sirolimus (Sir) inhibit lymphocyte proliferation at the cytokine transcription (CyA and Pred) or signal transduction (Sir) levels. Their double and triple interactions were studied on lectin-induced proliferation of whole blood lymphocytes (WBLP) from male and female humans, rabbits and rats. Isobols along with the Universal Response Surface Approach were used to describe and quantify the nature and intensity of drug interactions by determining α values. CyA was always less potent than Pred and Sir while these two compounds were relatively equipotent. Species-related differences were observed with single drugs. Rabbit WBLP were resistant to Pred action (Imax = 67%) and rats were more sensitive to Pred (IC50 = 9.1 nM in females) and Sir (2.8 nM) actions than humans (32 and 55 nM). Gender differences were observed but were not consistent across species. All double-drug combinations were synergistic, and combinations containing Pred were 10 to 100 times more synergistic in rabbits (αPred/Sir = 213 and αCyA/Pred = 147 in males) than in rats (12 and 2.1) or humans (3.7 and 5.7) in relation to the lower efficacy of Pred. Double-combination α values were able to describe CyA/Pred/Sir triple combination effects. These studies indicate that CyA, Pred and Sir act and synergistically interact in vitro in species- and gender-dependent fashions. Adrenalectomized rats better resemble humans in these responses. WBLP are useful in various species in determining immunosuppressive drug action and interactions.
Immunosuppressive compounds decrease cell-mediated reactivity by blocking rate-limiting steps in the immune response produced by T-helper and T-cytotoxic lymphocytes upon recognition of foreign or altered material (fig.1). These compounds are used in the prevention and treatment of graft rejection as well as in controlling autoimmune diseases like lupus erythematosus and rheumatoid arthritis (Scudeletti et al., 1990; Morris, 1992). However, their extended usage often is limited by severe side effects such as nephrotoxicity (CyA), diabetes mellitus (steroids) and hyperlipidemia (Sir) (Hricik et al., 1994; Murgia et al., 1996;Shoker, 1996). Therefore, combination therapy may be required to produce adequate immunosuppression with acceptable toxicity and should include compounds that are additive or synergistic only in their therapeutic effects. One strategy is to combine drugs blocking either T-cell activation or proliferation steps, which leads to several possible associations. For example, the classic maintenance therapy involves CyA, Pred and azathioprine (Shoker, 1996).

Immunosuppressive compounds currently used (bold) or in development for organ transplantation. Each compound is presented at its major site of action on lymphocyte activation and proliferation (S, synthesis; M, mitosis; D, differentiation).
Sirolimus is a new immunosuppressive compound with antifungal and antitumor activities. Sir acts through an original mechanism as it stops lymphocyte reactivity at the mid-late G1phase by blocking transductional signals produced by fixation of cytokines (e.g., IL-1, -2, -6) or growth factors to their membrane receptors (Sehgal, 1995; Metclafe and Richards, 1990) (fig.2). Sir interacts synergistically with CyA as they act through distinct mechanisms at the cytokine gene transcription or signal transduction levels. Indeed, CyA acts at the G0 phase by inhibiting calcineurin/calmodulin phosphatase activity required for IL-2 gene transcription by T-helper lymphocytes (Metclafe and Richards, 1990) (fig. 2). The CyA/Sir synergistic combination may be improved by addition of steroids or brequinar (Kahan et al., 1991, 1993). Corticosteroids act by decreasing cytokine (e.g., IL-1, -2, -6) and cell surface molecule (e.g., ICAM-1, LFA-1) gene transcription (Scudeletti et al., 1990; Paliogianni et al., 1993); brequinar bears antimetabolic properties blocking cells at the S phase (Klingermann, 1995). Therefore, as corticosteroids act through different mechanisms than CyA and Sir, it was hypothesized that their triple combination may produce additional additive or synergistic therapeutic effects compared with the double combinations.

Detailed mechanism of action of CyA (square), Pred (diamond) and Sir (triangle) in antigen-presenting cells and T lymphocytes. Inhibition is represented by a negative sign, stimulation by a positive sign within the drug symbol (Cyp, cyclophilin; TcR, T-cell receptor; Pg-P, P-glycoprotein; GRE, glucocorticoid repressive element; FRAP, FKBP-rapamycin-associated protein).
A convenient and controlled way to determine the nature and intensity of drug action and interaction is by studying in vitrosystems mimicking in vivo settings. The most widely used methods to assess immunosuppressive compound activity are the mitogen-induced lymphocyte proliferation assays usually performed on splenocytes (mouse, rat, rabbit, pig) or whole blood isolated lymphocytes (rabbit, pig, human) (Metclafe and Richards, 1990; Kayet al., 1991). These methods require cell separation and animal death because either spleens or large quantities of blood are needed, rendering them inadequate for multiple measurements. WBLP assays are a preferred choice because they better mimic the natural environment (Bloemena et al., 1989; De Groote et al., 1992), require very small blood volumes (10 μl per point) and are applicable to double-drug interaction assessment (Piekoszewskiet al., 1994). Therefore, WBLP interaction studies were extended successfully from humans to rabbits by Piekoszewski et al. (1994) and to rats in this study. It was important to developin vitro methods applicable to several species to select the most representative one, because species-related differences were often detected.
Currently, in animal studies, the nature and intensity of interactions observed between two or more compounds usually have been described and quantified by two methods: the Chou and Talalay combination index and Drewinko score methods (Stepkowski and Kahan, 1993; Piekoszweskiet al., 1994). Both methods lack in fully describing the interaction surface by not providing a reliable statistical summary parameter (Greco et al., 1995). Therefore, the isobologram method (Gessner, 1974) was used to describe the double-drug interactions and the URSA (Greco et al., 1990) was applied to quantify the extent of double- and triple-drug interactions. The major advantages of the last method are its full description of the interaction surface, its determination of a statistical summary parameter, α, along with its interpolating and extrapolating capabilities.
This paper presents the assessment of CyA/Pred/Sir interactions in humans, rabbits and rats with in vitro lectin-induced WBLP assays. To uncover possible species- and gender-related differences in these interactions, the data were analyzed with isobols and the URSA method.
Materials and Methods
Animals
Adrenalectomized Sprague-Dawley male (n = 6) and female (n = 6) rats weighing 250 to 320 g were purchased from Harlan Sprague-Dawley Inc. (Indianapolis, IN). New Zealand male (n = 4) and female (n = 5) rabbits weighing 4 to 5 kg were purchased from Becken Farms (Sanborn, NY). Animals were housed in a 12-h light/dark cycle, constant temperature environment (22°C). Rats had free access to rat chow (Agway RMH 1000) and physiological saline water. Rabbits received 125 g of rabbit chow (PMI Feeds Inc., St. Louis, MO) per day and had free access to tap water. Animals were acclimatized for 10 days before experiments and were kept in a stress-free environment throughout these studies. Analysis of plasma from the rats showed no corticosterone present. This research adhered to the principles of Laboratory Animal Care (NIH publication no. 85–23, revised 1985).
Humans
Caucasian male (n = 6) and female (n = 3) drug-free volunteers were included in our study; women receiving birth control pills were excluded. Volunteers were in good health and they were between 25 and 40 years of age. Blood was collected at 9 a.m. on the day of the experiment.
Reagents
CyA and Sir were obtained as gifts from Sandoz Research Institute (E. Hanover, NJ) and Wyeth-Ayerst Research (Princeton, NJ). Pred and PHA-L were purchased from Sigma Chemical Co. (St. Louis, MO) and concanavalin-A from ICN Biomedicals Inc. (Aurora, OH). Stock solutions of each drug were made in ethanol and kept at −20°C (CyA and Pred) or −80°C (Sir) for the duration of the study. Serial dilutions of the stock solutions with complete media at pH 7.3 to 7.5 were made freshly on each experimentation day in borosilicate sterile glass tubes. Final ethanol concentrations in incubation wells were always less than 0.23%.
Whole Blood Lymphocyte Proliferation
Lymphocyte proliferation was induced by the nonspecific T-cell mitogens PHA-L and concanavalin-A. The assay was optimized for humans, rabbits and rats, in terms of whole blood volume in the well, incubation time with and without [3H]thymidine and mitogen concentrations. Blood was kept at room temperature for use within 6 h of collection.
Human.
Blood was collected in evacuated heparinized glass tubes and diluted 1:20 (v/v) with whole blood human complete medium [RPMI 1640 supplemented with 2 mM l-glutamine, 20 mM HEPES, 100 U/ml penicillin, 100 μg/ml streptomycin, 0.25 mM 2-mercaptoethanol (Gibco)]. Diluted blood (160 μl) was plated in 96-well plates after addition of the different drug combinations. Proliferation was induced by PHA-L at an optimized final concentration of 3 μg/ml. All cultures were performed in triplicates in a total volume of 200 μl per well. After incubating cultures for 72 h at 37°C in a 7.5% CO2-humidified air incubator, cultures were pulsed with 1 μCi of [3H]thymidine per well (New England Nuclear, Boston MA) for 20 h. The cells were then harvested onto microplates (Packard Instrument Company, Meriden CT), washed with 3% hydrogen peroxide, dried and counted in liquid scintillation fluid (Microscint-20, Packard) in a Top Count Microplate Scintillation Counter (Packard).
Rabbit.
Blood was collected from the ear artery into glass tubes containing heparin and diluted 1:10 (v/v) with rabbit WBLP medium [human WBLP medium supplemented with 2.5% of heat-inactivated fetal calf serum (Gibco)]. Lymphocyte proliferation was induced by PHA-L at a final concentration of 5 μg/ml. At 72 h, cultures were pulsed with [3H]thymidine for 16 h to obtain the maximum response. Because macrophages were present, addition of a trypsin wash was necessary during the harvesting phase.
Rat.
Adrenalectomized rats were chosen because the maximum proliferation was insufficient in blood from normal rats. Blood was collected from the tail artery of anesthetized rats (ketamine/xylazine mixture) and diluted 1:15 (v/v) with rat WBLP medium [human WBLP medium with 2.5% of heat-inactivated new born calf serum (Gibco)]. Lymphocyte proliferation was induced by concanavalin-A instead of PHA-L at an optimized final concentration of 10 μg/ml because experiments with PHA-L showed insufficient mitogenic response. At 96 h after induction of lymphocyte proliferation, cultures were pulsed with [3H]thymidine for 24 h before the harvesting procedure.
Interaction Assays
Interaction experiments were conducted in 96-well plates. Each plate contained 96 different conditions (1/well): 90 drug combinations, 4 maximum stimulation controls (Smax) and 2 background (no drug and no mitogen) samples. Triplicates were determined by preparing three identical plates. The CyA/Pred/Sir single-drug concentration-effect relationship was defined with ten different concentrations spanning 4 log-units (30 conditions). To properly define each interaction with isobols and the URSA method, five combination ratios at the IC50 (1:1, 1:2, 2:1, 1:4 and 4:1) were studied and characterized with three combination concentrations (15/interaction), chosen to theoretically produce 25, 50 and 75% inhibition of Smax assuming additive interaction. For CyA/Pred/Sir triple interaction, five ratios at the IC50 were 1:1:1, 4:1:1, 1:4:1, 1:1:4 and 4:1:4. All drug combinations were prepared in a stock plate and dispensed with multichannel pipettes. The use of stock plates allowed for studying several subjects under identical conditions.
Data Analysis
Single-drug inhibition.
Observed effects were expressed directly as percent of Smax because background noise was negligible (<0.1%Smax). The inhibitory concentration-effect relationship was modeled with the sigmoidImax model:
Double and triple interaction with the URSA.
Combination data were analyzed with the isobologram method (Gessner, 1974) and URSA (Greco et al., 1990). The URSA equation applied to the CyA/Pred double interaction is:
For the triple-drug combination data, the expected effect was calculated with equation 2 written for each compound and the three double interactions. If the difference between observed and expected effects was not significantly different from zero, it was concluded that there was no alteration of the three double-drug interactions inside the triple combination. If the difference was significantly greater than zero, additional synergism is found within the triple combination; if the difference is negative, antagonism is produced.
Statistical Analysis
URSA analysis was performed by means of nonlinear regression fitting by the ADAPT II program (Biomedical Simulations Resource, Los Angeles, CA) applying the GLS and the bisection technique described byGreco et al. (1990) and modified by Ferron et al.(1998). An advantage of GLS over the maximum likelihood procedure is its computation of confidence intervals around parameter estimates. Single-drug parameters (Back, IC50, γ) were obtained initially and used as initial estimates in the quantitation of double-drug combination data (Back, IC50, γ and α). Assuming a normal distribution of the errors from the observed data and predicted values, the variance (Vari) was defined as: Vari = ς1·M(θ,Ci)ς2, where ς1 and ς2 are the vectors of variance, θ is the vector of parameters for the dynamic model (Back, IC50, γ and α) andM(θ,Ci) are the predicted values from the URSA at combined concentration Ci.
Statistical results are expressed as mean ± S.D. or median (range). Parametric and nonparametric tests were used to study gender, species and compound differences. Relations between parameters were quantified with orthogonal regressions because each estimated parameter has a certain degree of error. Data analysis was performed with SAS software (Version 6; SAS Institute, Cary, NC) and the level of significance of P < .05 was chosen.
Results
Single-drug inhibitions.
CyA, Pred and Sir single-drug inhibition curves for selected males and females for the three species are presented in figure 3. The effect-concentration relationship can be described by the sigmoidImax model (eq. 1). Maximum stimulation (Smax) was achieved at low drug concentrations. With increased concentrations, lower mitogenic responses were obtained in relation to drug immunosuppressive properties. The maximum inhibition (Imax) usually was observed at drug concentrations between 100 and 10,000 nM and concentrations needed to achieve 50% ofImax (IC50) were greater for CyA than Pred and Sir regardless of gender and species, with the exception of male rabbits. Moreover, rats were more sensitive than humans and rabbits to Pred action (fig. 3, lower center panel). Single-drug parameters (Back, IC50 and γ) for CyA, Pred and Sir are reported in table1 for each gender and species. Individual parameter values are displayed in figure4, allowing for direct visualization of the spread of individual estimates.

CyA, Pred and Sir inhibition of WBLP induced by lectins in humans (triangle), rabbits (circle) and rats (square). The upper panels display representative results from females (white); the bottom panels from males (gray). Fitted lines were drawn with estimated parameter values.
Summary parameters for CyA, Pred and Sir single-drug inhibition of lymphocyte proliferation

Summary of CyA, Pred and Sir single-drug parameters in females (open) and males (gray), for human, rabbit and rat WBLP assays. Horizontal lines present median values.
CyA acts at the lymphocyte activation step blocking cells at the G0 phase. This drug typically inhibited more than 96% of the maximum proliferative response. Species and gender differences were observed. In males, CyA was 4 times more potent in rabbits and 1.75 times more potent in rats than in humans, in whom the mean CyA IC50 was 396 nM. In female rats and rabbits, with IC50 approximately 80 nM, CyA was three to four times more potent than in women. Females were more sensitive to CyA action than males, and this finding may be because of the higher hematocrits in males.
Pred acts at the G1 phase by inhibiting cytokine gene transcription in antigen presenting cells (e.g., IL-1) and T lymphocytes (e.g., IL-2). Rabbits were more resistant to steroid action: mean Imax was 67% and similar among males and females. In other species, Pred inhibited more than 98% of the maximum proliferative response. Female rats were found to be significantly more sensitive to Pred action (IC50 = 9.12 nM) than rabbits (22.8 nM) and humans (31.6 nM). However, in males, rabbits were the less sensitive species with a mean IC50 of 339 nM.
Sir acts at the mid-late G1 phase by inhibiting the cascade of events induced by the binding of cytokines to their membrane receptors (e.g., IL-1 and -2). High concentrations of Sir inhibited 78 to 99% of the maximum proliferative response, depending on gender and species. Humans were the least sensitive species with a mean IC50 of 37.8 nM in men compared with 29.4 nM in male rats and 6.38 nM in rabbits. In females, rats were more sensitive with an IC50 of 2.77 nM.
All compounds inhibited lymphocyte proliferation from 67 to 99%, and CyA potency was found significantly lower than Pred and Sir potencies, except in male rabbits. Similar potency of Sir and Pred was observed with the same exception. No pattern for the steepness parameter γ was discernible.
Double- and triple-drug interactions.
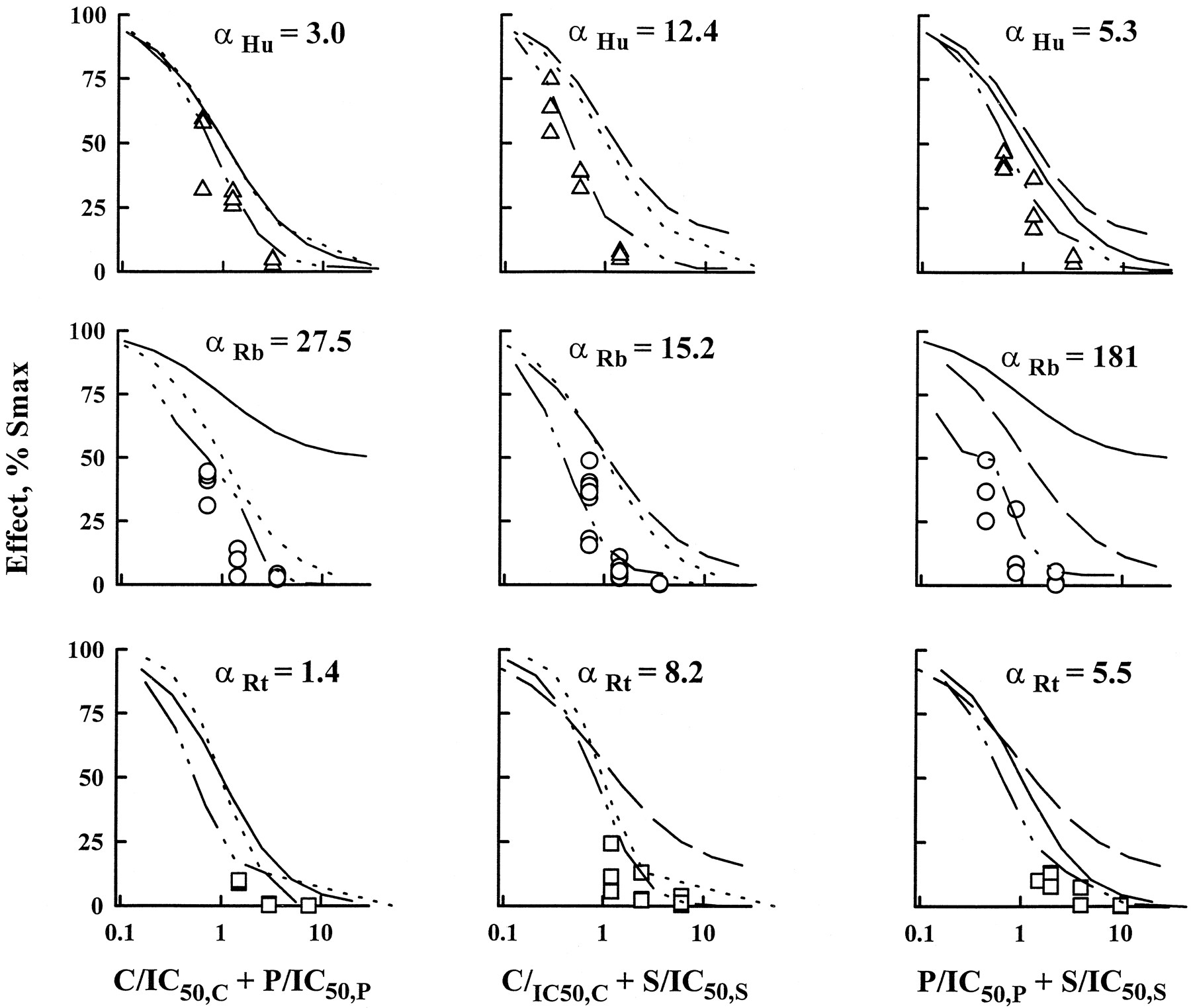
The major results for this study are in the full characterization of CyA/Pred/Sir double and triple combinations by the isobologram method and URSA. All combinations were mainly synergistic. Most of the ratios were found in the synergistic region of the isobolograms drawn at the IC50 and IC90 (fig.5). The combination effect curves with IC50 normalized concentrations were shifted to the left of single-drug effect curves (fig.6) and α values were always greater than zero (table 2 and fig.7). The relationship between isobologram shapes and URSA α values can be seen in figure 5 because greater bowing in the isobols translated into larger α values; this is especially clear with rats (bottom of the figure). When combination data are presented as an interaction surface (fig.8), the extent to which the three-dimensional fishnet curves scooped out represents the degree of the synergistic interaction. Higher synergism will be seen by a tightening of the fishnet into the bottom far left corner of the cube.
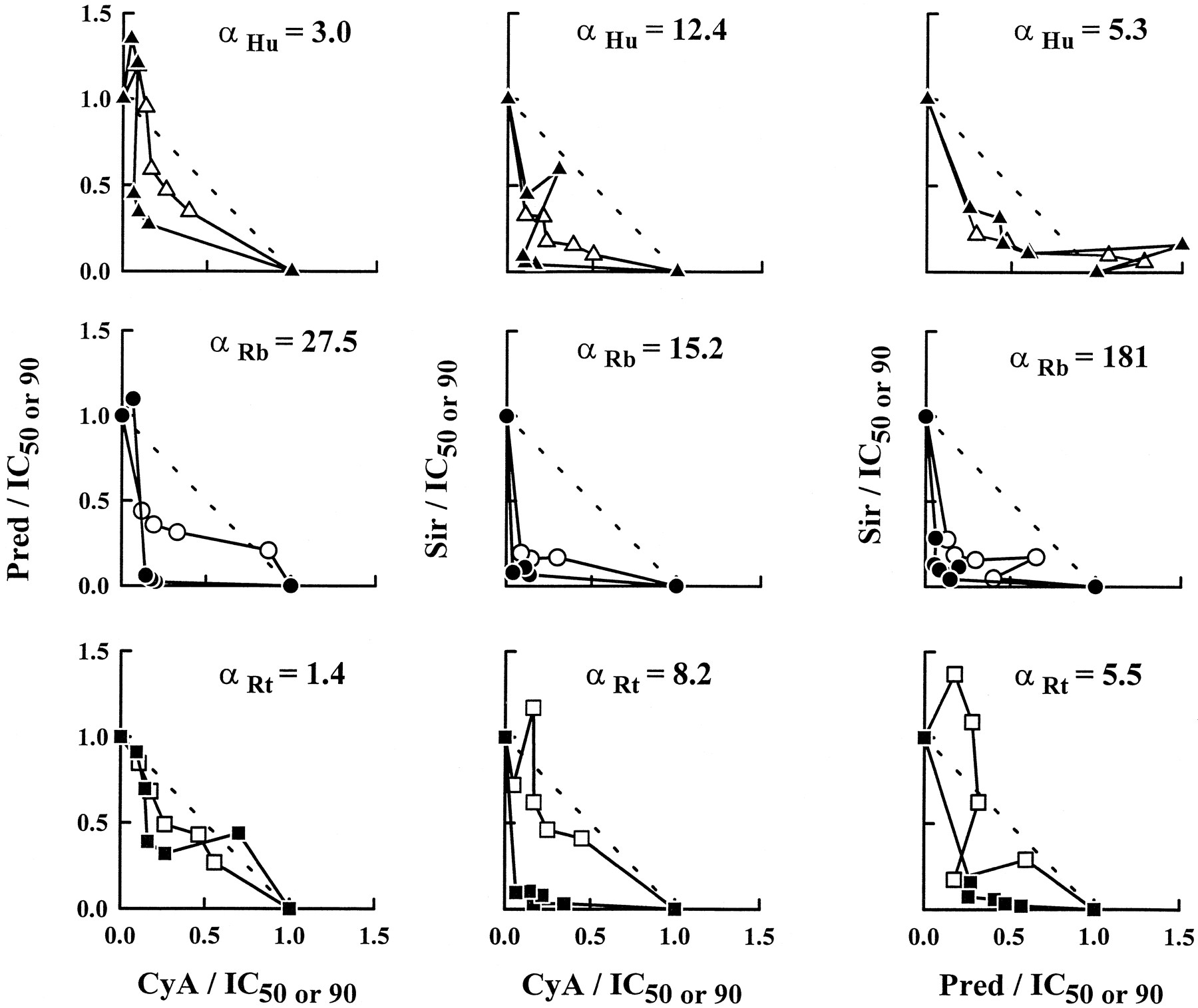
Isobolograms at the IC50 (open symbol) and IC90 (solid symbol) for the three double-drug interactions constructed with the five drug combination ratios at the IC50 (1:1, 1:2, 2:1, 1:4 and 4:1) in representative humans (upper panels), rabbits (middle) and rats (lower). Diagonal lines depict additivity. α values were obtained with URSA.
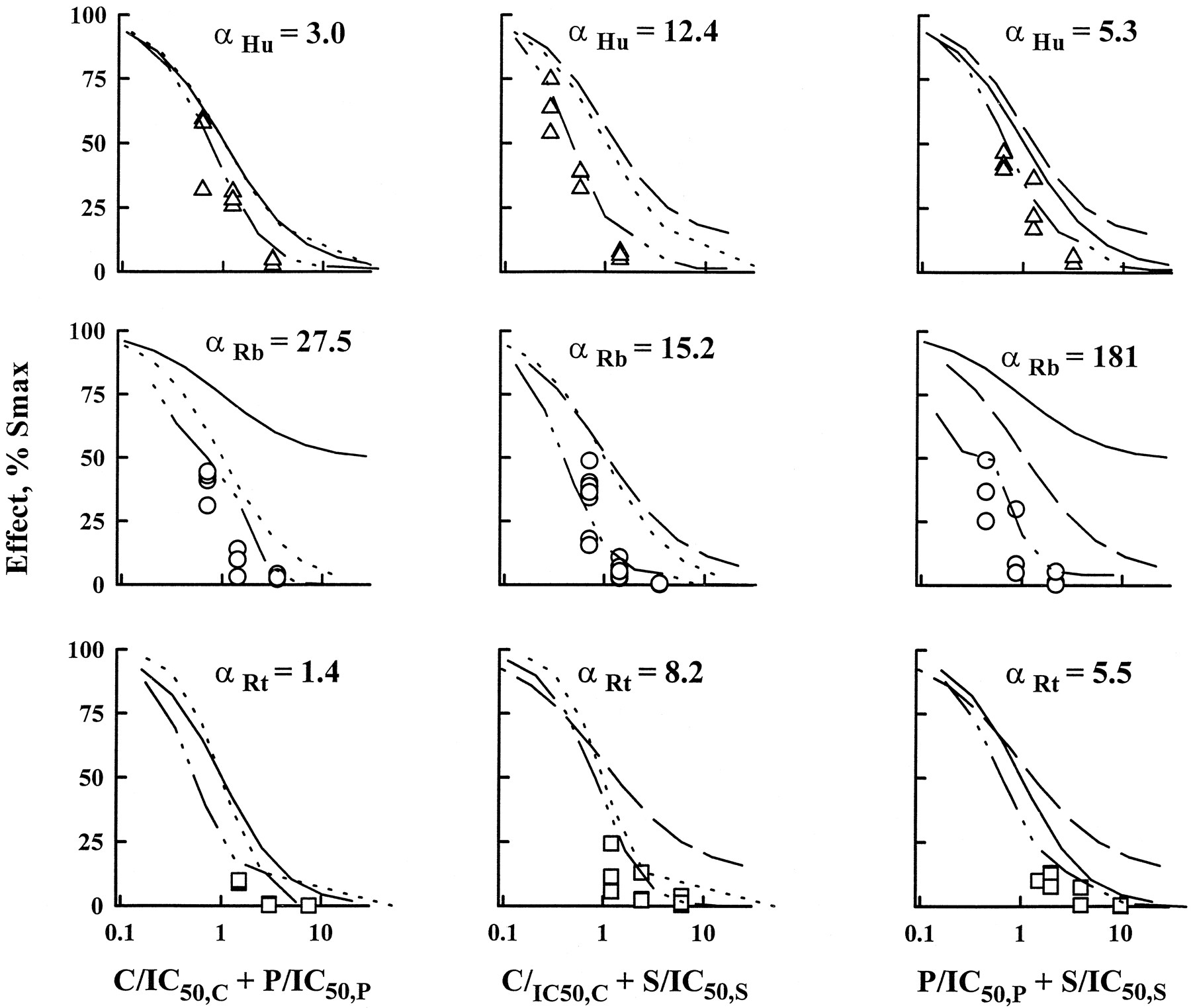
URSA for the three double-drug interactions in the representative subjects used in figure 3. Data obtained for drug ratios 1:1 at the IC50 are displayed along with the predicted effect obtained from URSA. Single-agent inhibition curves are presented: CyA (dotted line), Pred (solid line) and Sir (dashed line). Drug concentrations are normalized by their respective IC50values. A shift of the interaction curve to the left of single-drug curves indicates synergism (α > 0) and the absence of shift, additivity (α = 0). α values are reported.
Summary of α values for double-drug combinations

Three-dimensional representation of URSA for Pred/Sir interaction in humans (left), rabbits (middle) and rats (right) along with the isobolograms at the IC50. Seventeen different ratios were studied with 5 to 8 points per ratio. The fishnets are the best fitting surface. The 90 combination data set is plotted on the same graph, with vertical lines which indicate the distance between the mean data points and the surface. Solid symbols are above, and open symbols, below the surface.

Summary of CyA, Pred and Sir double-interaction parameter (α) in females (open symbol) and males (gray symbol), for human, rabbit and rat WBLP assays. Horizontal lines are median values.
Similarly to single-drug inhibition data, gender- and species-related differences were observed in double-drug interactions (table 2, fig.7). Rabbits differed significantly for combinations including Pred as larger α values were determined. As an example, the mean α for Pred/Sir interaction in males was 214 in rabbits, 3.7 in humans and 12 in rats. This finding may be in relation to the lower efficacy of Pred in rabbits as displayed on figure 9. With higher Pred Back or IC50 values, two parameters significantly correlated across species, greater α values were estimated. Confirming the visual inspection, significant correlations were found between CyA/Pred α values and Pred Back and IC50 values with or without log transformation. Moreover, CyA/Pred and Pred/Sir α values were correlated significantly (P = .0025) (fig. 9). Rats and humans gave similar α values for the three double-drug interactions, and the only gender effect was detected in humans with the Pred/Sir combination because it was two times more synergistic in women.

Relation between CyA/Pred or Pred/Sir α values and Pred Back or IC50 estimates. All species and genders are plotted with the symbols described previously. Lines were obtained from orthogonal regressions.
Comparing the three double-drug interactions, greater synergism was obtained for combinations involving Pred in rabbits or Sir in humans and rats. For example, the CyA/Sir combination was five times more synergistic than CyA/Pred, but similar to the Pred/Sir interaction in male rats.
For the triple interaction, all differences were not significantly different from zero. Therefore, double-combination α values were reasonably able to describe CyA/Pred/Sir interaction effects (fig.10). No additional synergism was found within the triple combination because estimated and observed effects were similar. Indeed, a constant departure of observed effects to the left (vs. right) of predicted values will have signified additional synergism (vs. antagonism) within the triple combination. The extrapolative advantage of URSA was demonstrated by simulations where the concentrations of each agent combining to produce 50 or 90% inhibition of the maximum stimulation were estimated (fig.11).

URSA applied to triple-drug interaction obtained in the representative subjects of figure 4. CyA (dotted line), Pred (solid line) and Sir (dashed line) single-agent effects are simulated along with the triple-combination effects assuming double-drug interaction α values. Observed effects are reported for the ratio 1:1:1 at the IC50. Drug concentrations were normalized by their respective IC50 values.

URSA simulations of CyA, Pred and Sir concentrations to combine in order to obtain 50% inhibition (gray surface) or 90% inhibition of the maximum response. Simulations were performed with mean parameter values determined in males (table 1).
Discussion
Uncontrolled immunologic responses account for graft failures and for the development and worsening of autoimmune diseases (Shoker, 1996). Therefore, immunosuppressive therapy is used to decrease humoral and cell-mediated reactivity by partially blocking rate-limiting steps in the immune response produced by T and B lymphocytes on recognition of foreign or altered materials. However, extended usage of immunosuppressive compounds often is limited by severe side effects such as nephrotoxicity (CyA), diabetes mellitus (steroids) and hyperlipidemia (Sir) (Hricik et al., 1994; Murgia et al., 1996; Shoker, 1996). Therefore, combination therapy may be required to produce adequate immunosuppression with acceptable toxicity and should include compounds that are additive or synergistic only in their therapeutic effects.
A convenient and useful way to determine the nature and intensity of drug action and interaction is by studying in vitro systems mimicking in vivo settings. The most widely used methods to assess immunosuppressive compound activity are the mitogen-induced lymphocyte proliferation assays usually performed on splenocytes or whole blood isolated lymphocytes (Metclafe and Richards, 1990; Kayet al., 1991). These methods require animal death or large quantities of blood, rendering them inadequate for multiple measurements. WBLP assays are preferred because they better mimic the natural environment (Bloemena et al., 1989; De Grooteet al., 1992) and require very small blood volumes. In the present paper, WBLP assays were extended successfully from single-drug effect characterization in humans to double and triple interaction assessment in humans, rabbits and rats. It was important to developin vitro methods applicable to several species because many animal models are used to study these agents.
Species-related differences were detected for CyA, Pred and Sir used as single agents. The major difference was observed in the resistance of rabbit cells to Pred action as the maximum inhibition was 67%, confirming results previously obtained with methylprednisolone, another synthetic glucocorticoid (Piekoszewski et al., 1994). This finding may be in relation to the higher macrophage density present in rabbit lymphocyte cultures compared with humans and rats where these were fewer. Macrophages are required for lymphocyte proliferation particularly in rabbits (Kapsenberg et al., 1984). Indeed, adherent cells were present in detectable quantities at the end of the culture, which required an additional trypsin wash during the harvesting phase. Adherent cells accounted in preliminary studies for 30 to 40% of the total mitogenic effect measured. Thus, the observed resistance of rabbit WBLP to Pred action may be related to lower efficacy of steroids on macrophage proliferation.
Differences in lymphocyte sensitivity between humans, rabbits and rats were found for all compounds studied. In all species, CyA was the least effective agent in inhibiting lymphocyte proliferation induced by lectins. Rat was confirmed as a more sensitive species for steroid action than humans and rabbits. This may be partially because of the difference in T-lymphocyte mitogens used, because concanavalin-A (rats) and PHA-L (rabbits and humans) may stimulate different lymphocyte subsets. However, in vitro rat splenocyte proliferation stimulated with PHA-L showed sensitivity to Pred similar to rat WBLP stimulated with Con-A (Meno-Tetang and Jusko WJ, unpublished observations). Moreover, human lymphocytes were less sensitive to CyA and Sir action than rabbits and rats. These two immunosuppressive agents are highly distributed into red blood cells because of their respective intracellular bindings to cyclophilin and FKBP12. In vitro whole blood distribution studies with CyA showed whole blood/plasma ratios lower than two, and no differences were observed among rats, dogs and humans (Takada et al., 1993). Thein vitro whole blood/plasma ratio of Sir was approximately 10 in humans (Yatscoff et al., 1993), 5 in rabbits and 1 in rats; these ratios were not altered by the presence of high concentrations of Pred or CyA (Ferron and Jusko, 1998). Similar mean hematocrits were obtained in humans (0.47) and rabbits (0.45), but lower values were observed in rats (0.39). Therefore, variabilities in hematocrit or red blood cell binding capabilities across species may partially account for sensitivity differences.
CyA, Pred and Sir act through distinct mechanisms at the cytokine gene transcription or signal transduction levels (figs. 1 and 2). CyA acts at the G0 phase by inhibiting calcineurin/calmodulin phosphatase activity required for IL-2 gene transcription by T-helper lymphocytes (Metclafe and Richards, 1990). Corticosteroids decrease cytokine (e.g., IL-1, -2, -6) and cell surface molecule (e.g., ICAM-1, LFA-1) gene transcription (Scudeletti et al., 1990; Paliogianni et al., 1993) partially by increasing the production of NF-κB inhibitory factor, Iκ-Bα (Auphan et al., 1995; Scheinmanet al., 1995). Sir arrests lymphocytes at the mid-late G1 phase by blocking transductional signals produced by fixation of cytokines (e.g., IL-1, -2, -6) or growth factors to their membrane receptors (Sehgal, 1995; Metclafe and Richards, 1990). Moreover, Sir and CyA bind to the cytoplasmic glucocorticoid receptor complex, directly (via hsp59) or indirectly through binding to cyclophilin-40, possibly altering receptor functionality (Ning and Sanchez, 1993, 1994). In addition, CyA and Sir inhibit P-glycoprotein function (Ning and Sanchez, 1996). Therefore, because CyA, Sir and Pred acted through distinct mechanisms and displayed species-related differences as single agents, it was hypothesized that similar differences will be discovered for their double and triple combinations.
CyA, Pred and Sir double and triple combinations were assessed successfully in humans, rabbits and rats, with a sparse design, WBLP assays and the isobologram method coupled with the URSA. Combining careful graphical examination with the statistical analysis led to an adequate description of the nature and intensity of each interaction. With isobols, each combination ratio was studied and most isobols lacked symmetry (fig. 5) in relation to a ratio dependence in the nature of drug interactions. In Pred/Sir combinations (fig. 7), synergism was found for low concentrations of Pred, but antagonism was obtained for higher Pred concentrations. A similar trend was discovered for CyA/Pred combinations (fig. 5). CyA and Sir interact with Pred at the P-glycoprotein level by partially blocking the pump and decreasing steroid efflux (Ning and Sanchez, 1996). Preponderance of Pred over CyA or Sir may overflow this inhibitory mechanism, creating lower synergism or even antagonism. Global description of drug interactions by URSA was obtained by determining α values (figs. 7 and 8), and all combinations were synergistic in relation to the different levels of drug action (fig. 2). Species-related differences were found because α values for drug associations involving Pred were significantly higher in rabbits. This may be caused partly artificially by the lower efficacy of Pred in rabbits, because similar isobols were obtained for humans, rabbits and rats (fig. 8). Indeed, it can be noted on figure 7that the three-dimensional interaction surfaces were smoother in humans and rats than in rabbits. Pred combinations may only be two to three times more synergistic in rabbits with a full design instead of the 20-times factor obtained from the sparse design. Taking the three double-drug interactions together, higher synergism was found in rabbits for the triple combination (fig. 10) compared with rats and humans which behaved similarly. Therefore, combining single-drug effects with interaction data, adrenalectomized rats seem to be a better species than rabbits for studying CyA, Pred and Sir actions and interactions at the immunodynamic level.
Gender-related differences were observed with single drugs as well as in interactions but were not constant across species. For CyA, female rats and humans were more sensitive to CyA action than males, but no gender differences were found in rabbits. Moreover, only female rabbits showed greater sensitivity than males to Pred action, because this previously reported gender effect for humans unexpectedly was not recovered from our experiments (Lew et al., 1993). Male humans and rabbits as well as female rats were more sensitive to Sir action. Because gender differences were not consistent across species, extrapolations from one species to another are tenuous.
Within the triple interaction, no additional synergism was detected (fig. 10) and URSA extrapolating capabilities helped in the computation of combined CyA, Pred and Sir concentrations to produce 50 or 90% inhibition of the maximum stimulation (fig. 11). Because CyA, Pred and Sir display nonoverlapping side effects, their combination is promising in transplantation as well as in prevention of autoimmune diseases or allergic reactions (Sehgal, 1995; Haczku et al., 1994). Moreover, combinations with Sir are of particular interest as Sir may reduce chronic rejection by inhibiting arterial smooth muscle cell proliferation (Shoker, 1996; Schmidbauer et al., 1994). Additional pharmacodynamic and pharmacokinetic interactions may arisein vivo because CyA, Pred and Sir are metabolized by CYP3A4 and steroids alter lymphocyte trafficking, decreasing the number of T-helper lymphocytes present in the blood stream in humans, rabbits and rats (Lew et al., 1993; Piekoszewski et al., 1994). Decreased Pred clearance with increased Sir exposure was observed in renal transplant patients (Jusko et al., 1996). Therefore, greater synergistic interactions between the three compounds may occur in vivo (Kahan, 1997).
In conclusion, CyA/Pred/Sir double and triple interactions were examined in humans, rabbits and rats using in vitro whole blood lymphocyte proliferation assays. All combinations were found to be synergistic. Differences were found especially between species, whereas few gender-related differences were observed. Rats, albeit adrenalectomized, seem to resemble humans better than rabbits in assessing CyA, Pred and Sir action and interaction on lymphocytes. Moreover, WBLP lymphocyte cultures are extremely useful in assessing the nature and intensity of drug interactions in vitro. The applicability of these methods to ex vivo WBLP cultures from treated animals will be tested further.
Acknowledgments
The authors thank Dr William R. Greco and his team for synergistic discussions and Nancy N. Pysczcynski for excellent technical assistance.
Footnotes
-
Send reprint requests to: William J. Jusko, Ph.D., Department of Pharmaceutics, State University of New York at Buffalo, 565 Hochstetter Hall, Buffalo, NY 14260.
-
↵1 This work was supported by grant GM 24211 from the National Institutes of Health, Bethesda, MD and Fellowship support for G.M.F. from Wyeth-Ayerst Research, Radnor, PA.
- Abbreviations:
- α
- double drug interaction parameter using URSA
- Back
- % of Smax not affected by the drug
- CyA
- cyclosporin A
- γ
- slope of the concentration effect relationship
- GLS
- general least squares procedure
- IC50
- drug concentration producing response equal to 50% ofSmax − Back
- HEPES
- N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid
- ICAM-1
- intracellular adhesion molecule-1
- IL
- interleukin
- Imax
- maximum inhibition
- LFA-1
- lymphocyte function-associated antigen-1
- PHA-L
- phytohemagglutinin-L
- Pred
- prednisolone
- Sir
- Sirolimus (Rapamycin)
- Smax
- maximum stimulation (100%)
- URSA
- universal response surface approach
- WBLP
- whole blood lymphocyte proliferation
- Received September 8, 1997.
- Accepted March 10, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}