


Abstract
A novel member of the organic anion transporter (OAT) family, Oat5 (Slc22a19), has been reported to transport a naturally occurring mycotoxin, ochratoxin A (OTA). However, neither its endogenous substrate and driving force nor physiological functions have been determined. Herein, we report the functional characterization of rat Oat5 (rOat5), as well as its intrarenal distribution and membrane localization. When expressed in Xenopus laevis oocytes, rOat5 mediated the transport of sulfate conjugates of steroids such as estrone-3-sulfate (E1S; Km = 18.9 ± 3.9 μM) and dehydroepiandrosterone sulfate (Km = 2.3 ± 0.2 μM) in a sodium-independent manner, in addition to OTA. The rOat5-mediated E1S transport was strongly inhibited by four-carbon (C4) dicarboxylate succinate and longer dicarboxylates (C7–C9). The uptake of [3H]E1S via rOat5 was significantly trans-stimulated by succinate, and the efflux of [14C]succinate was significantly trans-stimulated by E1S. A similar trans-stimulatory effect of preloaded succinate on E1S uptake was also detected in cells stably expressing rOat5 (S2 rOat5). rOat5 interacted with chemically heterogenous anionic compounds. The rOat5-mediated E1S transport was inhibited by several sulfate conjugates, such as 4-methylumbelliferyl sulfate and β-estradiol sulfate, but not by glucuronide conjugates. An immunohistochemical study showed that rOat5 was localized at the apical membrane of renal proximal tubules in the corticomedullary region. rOat5 mRNA was expressed in the late segments (S2 and S3) of proximal tubules. These results indicate that rOat5 is renal organic anion/dicarboxylates exchanger and, under physiological conditions, may function as an apical reabsorptive pathway for organic anions in proximal tubules driven by an outward gradient of dicarboxylates.
The kidney plays an important role in the elimination of harmful endogenous compounds and xenobiotics from the body. The proximal tubule is the primary site where numerous organic anions are taken up from the blood and excreted into the urine (Wright and Dantzler, 2004). The transcellular secretion of organic anions in the proximal tubule involves a two-step process: 1) the uptake of organic anions across the basolateral membrane of proximal tubular cells and 2) their excretion across the apical membrane into the tubular lumen. The former is energetically uphill and is accomplished by a tertiary active process via the organic anion/dicarboxylate exchanger that uses the outwardly directed gradient of five-carbon (C5) dicarboxylate α-ketoglutarate, one of the Krebs-cycle intermediates (Burckhardt et al., 2001). Two organic anion transporter (OAT) proteins, OAT1 (Sekine et al., 1997; Sweet et al., 1997; Wolff et al., 1997) and OAT3 (Kusuhara et al., 1999; Bakhiya et al., 2003; Sweet et al., 2003), have been proposed to be responsible for this step. The latter process is believed to be transporter-mediated, although this process is energetically downhill. In general, the efflux system for para-aminohippurate (PAH; a prototypical substrate for the organic anion transport system) in the brush-border (apical) membrane has been investigated using brush-border membrane vesicles (BBMVs). The electroneutral anion exchange systems and voltage-driven transport systems are considered to play a physiological role not only in the efflux of various organic anions but also in the reabsorption of some organic anions. However, the precise mechanism underlying the apical organic anion transport still remains unclear.
To date, several cDNAs encoding renal organic anion transporters have been successively identified in the kidney, including OAT, organic anion-transporting polypeptide (oatp), sodium-dependent inorganic phosphate transporter (NPT), and multidrug resistance-associated protein (MRP) families (Burckhardt et al., 2001; Russel et al., 2002). OAT4 (human), Oatp1 (rodent), Oat-k1/k2 (rodents), and NPT1 (human and mouse) are localized at the apical membrane of proximal tubules and are the possible candidates for an anion-exchange system. Regarding the ATP-driven organic anion transport system, MRP2 and MRP4 are also localized at the apical membrane of proximal tubules, although the relative contributions of both transporters to the apical efflux of organic anions have yet to be identified (Russel et al., 2002; van Aubel et al., 2002; Smeets et al., 2004). Recently, we have identified a novel voltage-driven organic anion transporter (OATV1) at the apical membrane of pig renal proximal tubules (Jutabha et al., 2003). OATV1 seems to be the long-hypothesized potential-driven facilitator in pig BBMVs (Werner et al., 1990; Krick et al., 2000). OATV1 exhibits the highest amino acid sequence identity to NPT1 (60–65%), although the functional properties of both transporters are different. Therefore, it should be elucidated whether OATV1 is an ortholog of human NPT1 (Uchino et al., 2000). NPT1 was suggested to be a good candidate for the apical potential-driven facilitator in human proximal tubules characterized in earlier experiments on BBMVs.
The number of OAT isoforms is rapidly increasing to date. Among them, both human and rodent cDNAs for OAT1 to OAT3 were reported, whereas only human cDNA was cloned for OAT4 (Koepsell and Endou, 2004). Very recently, Youngblood and Sweet (2004) reported the novel murine organic anion transporter mOat5 (Slc22a19) expressed in the kidney. They demonstrated that mOat5 mediates the transport of mycotoxin ochratoxin A (OTA), but their study was very limited. For example, they could not determine its endogenous substrate and driving force or its intrarenal localization related to its physiological functions. In the present study, we isolated this novel OAT member Oat5 from the rat kidney by a homology search of an expressed sequence tag (EST) database using a human OAT4 sequence. Rat Oat5 (rOat5) mediates the sodium-independent transport of sulfate conjugates of steroids, such as estrone-3-sulfate (E1S) and dehydroepiandrosterone sulfate (DHEAS) as well as OTA. The most surprising finding of our study was that rOat5 interacted with succinate, which has not been reported as a counterion for the classical renal organic anion transport system to date, to mediate organic anion/succinate exchange. These data indicate that rOat5 is renal organic anion transporter that may mainly function as an apical pathway for the reabsorption of some organic anions driven by an outward gradient of dicarboxylates, including succinate. The functional characterization of rOat5 deepens our understanding of renal apical transport mechanisms of organic anions, including xenobiotics and endogenous compounds.
Materials and Methods
Materials. [3H]E1S (2.0 TBq/mmol) and [3H]DHEAS (520 GBq/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [3H]Ochratoxin A (OTA) (547.6 GBq/mmol) was purchased from Moravek Biochemicals (Brea, CA). [14C]Succinate (4.07 GBq/mmol) was from American Radiolabeled Chemicals, Inc. (St. Louis, MO). [32P]dCTP was obtained from GE Healthcare (Little Chalfont, Buckinghamshire, UK). All other chemicals and reagents were of analytical grade and obtained from commercial sources.
Isolation of rOat5 cDNA. The EST database was searched for “query human OAT4 (hOAT4)”, and an EST clone (GenBank accession number AI058341) was identified. The [32P]dCTP-labeled probe was synthesized from this clone (I.M.A.G.E. ID No. UI-R-C1-kn-d-03-0-UI) and used for the screening of a rat kidney cDNA library. A nondirectional cDNA library was prepared from rat kidney poly(A)+ RNA using the Superscript Choice System (Invitrogen, Carlsbad, CA). The cDNA library was screened as described previously (Kusuhara et al., 1999). rOat5 cDNA was sequenced in both directions by the dye terminator cycle-sequencing method using an ABI PRISM 3100 genetic analyzer (Applied Biosystems, Foster City, CA).
Functional Characterization in Xenopus laevis Oocytes. cRNA synthesis and radiolabeled substrate uptake measurements were performed as described elsewhere (Kusuhara et al., 1999). Capped cRNA was synthesized in vitro using SP6 RNA polymerase from plasmid DNA linearized with RsrII. Defolliculated stage IV and stage V oocytes were injected with 25 ng of capped rOat5 cRNA and incubated in modified Barth's solution [88 mM NaCl, 1 mM KCl, 0.33 mM Ca(NO3)2, 0.4 mM CaCl2, 0.8 mM MgSO4, 2.4 mM NaHCO3, and 10 mM HEPES] containing 50 μg/ml gentamicin at 18°C. After two to three days of incubation, uptake and efflux experiments were performed at room temperature using ND96 solution (96 mM NaCl, 2 mM KCl, 1.8 mM CaCl2, 1 mM MgCl2, and 5 mM HEPES, pH 7.4).
Kinetic parameters for the uptake of E1S, DHEAS, and OTA via rOat5 were estimated from the following equation: v = Vmax × S/(Km + S), where v is the rate of substrate uptake (picomole/oocyte/hour), S is the substrate concentration in the medium (in micromolars), Km is the Michaelis-Menten constant (in micromolars), and Vmax is the maximum uptake rate (picomole/oocyte/hour). These kinetic parameters were determined with the Eadie-Hofstee equation.
To examine the trans-stimulatory effect of both the uptake and efflux of radiolabeled substrates, 50 nl of [14C]succinate (∼2 mM) or cold succinate (50 mM) was injected into oocytes expressing rOat5 with a fine-tipped glass micropipette as described previously (Jutabha et al., 2003). Individual oocytes then were washed with ice-cold ND96 solution twice, placed on ice for 5 min, incubated with ND96 at room temperature for 1 h, and then finally transferred into a medium with or without radiolabeled E1S or unlabeled E1S and incubated at room temperature for 1 h. Radioactivity in both the medium and oocytes was determined after a 1-h incubation.
For the uptake and efflux measurements in the present study, 8 to 10 oocytes were used for each data point. The values are expressed as mean ± S.E. We repeated each experiment at least twice to confirm the results. The results from representative experiments are shown in the figures.
Cell Culture and Establishment of S2 rOat5. The establishment of cells stably expressing rOat5 (S2 rOat5) was carried out as described previously (Takeda et al., 2002). These cells were grown in a humidified incubator at 33°C and under 5% CO2 using the RITC 80-7 medium containing 5% fetal bovine serum, 10 μg/ml transferrin, 0.08 U/ml insulin, 10 ng/ml recombinant epidermal growth factor, and 400 μg/ml geneticin. The cells were subcultured in a medium containing 0.05% trypsin-EDTA solution (containing 137 mM NaCl, 5.4 mM KCl, 5.5 mM glucose, 4 mM NaHCO3, 0.5 mM EDTA, and 5 mM HEPES, pH 7.2) and subjected to 25 to 35 passages.
Uptake Study Using S2 rOat5. Uptake experiments were performed as described previously (Takeda et al., 2002). Organic anion transport in S2 rOat5 was estimated by measuring the uptake rates of [3H]E1S and [14C]succinate. S2 rOat5 or S2 pcDNA 3.1(+) (S2 mock) cells (1 × 105 cells) were plated on 24-well plates and cultured for two days. After the medium was removed, cell monolayers were washed twice with Dulbecco's modified phosphate-buffered saline (D-PBS) (containing 137 mM NaCl, 3 mM KCl, 8 mM Na2HPO4, 1 mM KH2PO4, 1 mM CaCl2, and 0.5 mM MgCl2, pH 7.4) supplemented with 5.5 mM d-glucose, and preincubated for 10 min. The monolayers then were incubated with 500 μl of D-PBS containing 50 nM [3H]E1Sor10 μM[14C]succinate for 2 min at 37°C. The cells were washed three times with ice-cold D-PBS and solubilized in 0.5 ml of 0.1 N sodium hydroxide. The amount of substrate accumulated within the cells was then determined by measuring radioactivity.
To clarify the driving force and its transport direction, we examined the trans-stimulatory effect of succinate on rOat5-mediated E1S transport as described elsewhere (Ekaratanawong et al., 2004). For the experiments on the uptake of radiolabeled substrates, the cells were prepared as described above. In particular, the cells were preloaded with 0.5 ml of D-PBS with or without unlabeled substrates (5 mM succinate or glutarate) at 37°C for 30 min before uptake experiments.
Preparation of Anti-rOat5 Antibody. We generated a rabbit anti-rOat5 polyclonal antibody raised against a keyhole limpet hemocyanin-conjugated synthesized peptide REVKKDAVAKVTPF (538–551 of the amino acid sequence) corresponding to the 14 amino acids of the COOH terminus of rOat5.
Transfection, Crude Membrane Preparation, and Western Blot. Cell culture and transfection of HEK293 cells and the preparation of its crude membrane fractions were carried out as described previously (Anzai et al., 2004). pcDNA3.1-rOat5 or pcDNA3.1 vector (2 μg) was transfected into HEK293 cells.
Immunohistochemical Analysis. After anesthetizing the rats, their kidneys were perfused in situ with 4% paraformaldehyde-phosphate-buffered saline and excised. Kidney slices (2–3-mm thick) were fixed in the same solution at room temperature overnight and embedded in paraffin. Two-micrometer-thick sections were prepared, and after deparaffinization, the sections were incubated in 3% H2O2 for 15 min to abrogate endogenous peroxidase activity. Blocking was performed with 10% goat serum for 20 min. Then, the sections were further incubated overnight at 4°C with the anti-rOat5 antibody or anti-rOat3 antibody at 1:2000 dilution (Kojima et al., 2002). Thereafter, they were rinsed with 0.05 M Tris-buffered saline containing 0.1% Tween 20 and incubated with Envision(+) rabbit peroxidase (DakoCytomation California Inc., Carpinteria, CA) for 30 min. After treatment with diaminobenzidine (0.8 mM; Dojindo Laboratories, Kumamoto, Japan), the sections were counterstained with hematoxylin and examined under a light microscope.
Microdissection of Rat Nephron and Reverse Transcription-Polymerase Chain Reaction. The nephron segments of the rats were microdissected as described previously (Nonoguchi et al., 1986) to obtain the following structures: glomeruli; proximal tubules (S1,S2,S3); medullarly thick ascending limb (MAL) and cortical thick ascending limb; and cortical collecting duct (CCD) and outer and inner medullary collecting ducts. Three glomeruli and 2 mm of each dissected tubule were used for reverse transcription (RT)-polymerase chain reaction (PCR).

rOat5-mediated uptake of organic anions. A, the uptake rates of radiolabeled compounds (E1S, 100 nM; DHEAS, 100 nM; and OTA, 100 nM) by control oocytes (open column) and rOat5-expressing oocytes (closed column) for 1 h were measured (mean ± S.E.M.; n = 8–10). B, direct comparison of representative substrate E1S transport mediated by rOat3, OAT4, and rOat5. The uptake rates of [3H]E1S (100 nM) by rOat3, OAT4, and rOat5-expressing oocytes were determined in the absence (closed column) or presence of 5 μM (open column), 50 μM (gray column), and 500 μM (dotted column) unlabeled E1S.
RT-PCR was performed using a cDNA synthesis kit and a PCR master kit as described previously (Ikebe et al., 2001). Specific primers for rOat3 and rOat5 were designed as follows: rOat3 sense primer, 5′-CCA GTC TTC CCA ATG ACA CCC AGA GG-3′, and antisense primer, 5′-CCA GAT AGA ACC AGC CAG CGT ATG GA-3′; and rOat5 sense primer, 5′-GGG AGG AAG TTC ATT GTG-3′, and antisense primer, 5′-GCA CAC CAC ACC ATA GAG-3′. The cDNA products of rOat3 and rOat5 were 556 and 656 base pairs in length, respectively.
Results
By the EST database search, we found one EST (GenBank accession number AI058341) whose translated amino acid sequence is highly homologous to OAT1, OAT2, OAT3, and OAT4. A rat kidney cDNA library was screened using the corresponding cDNA clone I.M.A.G.E. ID No. UI-R-C1-kn-d-03-0-UI as a probe, and cDNA encoding a membrane protein that represents the rat ortholog of the recently identified rOat5 was isolated. rOat5 cDNA consisted of 1995 base pairs encoding a 551-amino acid protein (GenBank/EBI Data Bank with accession number AB051836). rOat5 showed amino acid sequence identities of 39% to rat Oat1 (Sekine et al., 1997; Sweet et al., 1997), 37% to rat Oat2 (Sekine et al., 1998b) and rat Oat3 (Kusuhara et al., 1999), 47% to human OAT4 (Cha et al., 2000), and 44% to a recently cloned OAT member, human urate/anion exchanger URAT1 (Enomoto et al., 2002). rOat5 is obviously not the ortholog of the human OAT5 (hOAT5) (Sun et al., 2001). Kyte-Doolittle hydropathy plot analysis (Kyte and Doolittle, 1982) predicted 12-membrane-spanning domains in rOat5 (hydropathy plot not shown). As in the case for members of the OAT and OCT families, N-glycosylation (residues 39, 56, 62, and 102) and protein kinase C-dependent phosphorylation (residues 216, 272, 279, 313, and 536) sites were predicted on the basis of the rOat5 sequence.
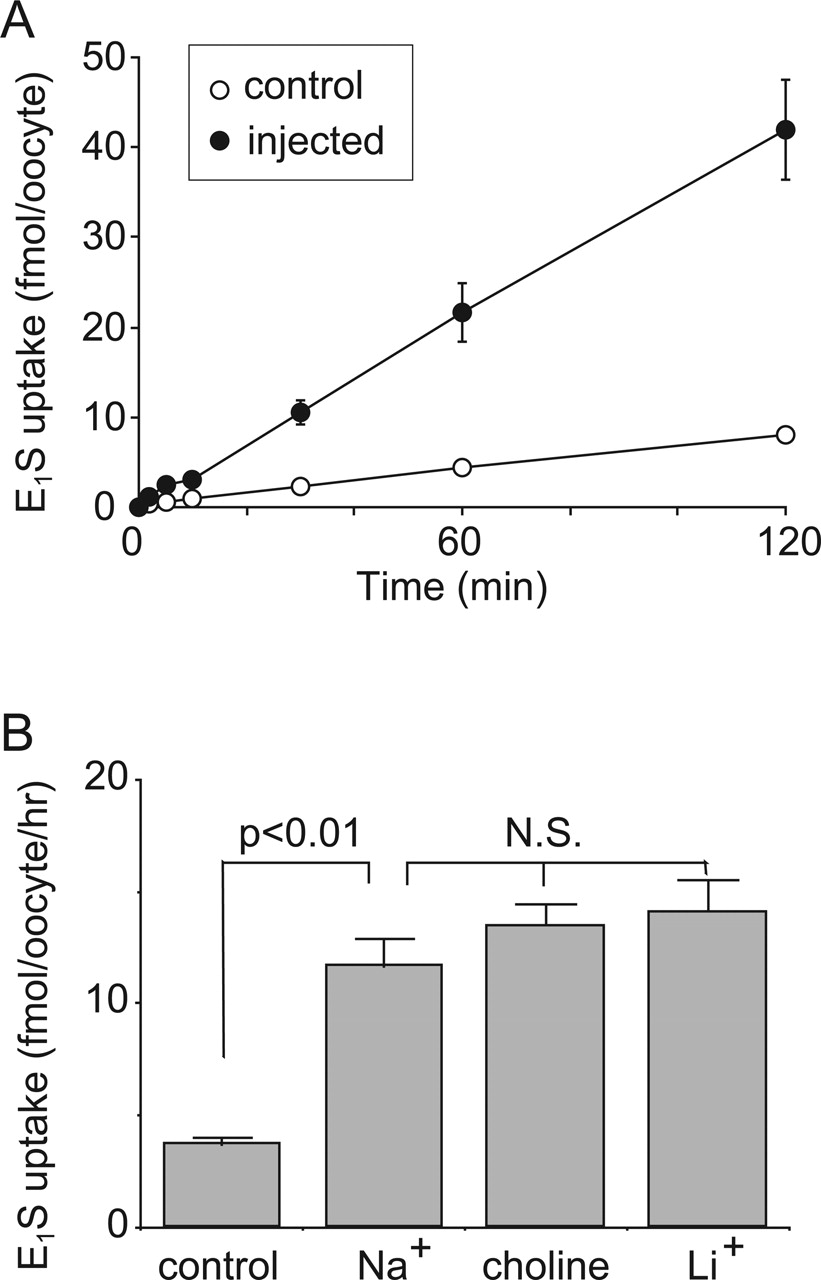
Transport properties of E1S via rOat5. A, the rates of uptake of 100 nM [3H]E1S by control oocytes (open circle) and rOat5-expressing oocytes (closed circle) were measured after 2 h of incubation. B, effect of extracellular cations on [3H]E1S uptake in X. laevis oocytes expressing rOat5. The rate of [3H]E1S (100 nM) uptake by control (open column) or rOat5-expressing (closed column) oocytes for 1 h was measured (mean ± S.E.M.) in the presence or absence of extracellular Na+. Extracellular Na+ was replaced with lithium or choline of equimolar concentration. N.S., not significant.
Using the X. laevis oocyte expression system, we investigated the transport of organic anions by rOat5 (Fig. 1). The uptake rates of [3H]E1S, [3H]DHEAS, and [3H]OTA in oocytes expressing rOat5 were 2- to 4-fold higher than those in control oocytes (Fig. 1A). rOat5 did not show any uptake of PAH (a representative substrate of OAT1), urate, salicylate, prostaglandins E2 and F2α, estradiol-17β-glucuronide, and tetraethylammonium (data not shown). To directly compare the transport rates of rOat5 with other renal OATs, we performed uptake studies of E1S, one of the representative substrates for OATs, with or without unlabeled E1S using rOat3-, OAT4-, and rOat5-expressing oocytes. As shown in Fig. 1B, although E1S uptake via rOat5 was strongly inhibited by 50 and 500 μM cold E1S, its rate of uptake was smaller than the rates of rOat3 and OAT4. In addition, the inhibition by 5 μM E1S to Oat5-mediated uptake was weak, whereas 5 μM E1S strongly inhibited rOat3 and OAT4-mediated uptake.

Representative rates of concentration-dependent uptake of [3H]E1S (A), [3H]DHEAS (B), and [3H]OTA (C) via rOat5. The rates of uptake of E1S, DHEAS, and OTA at various concentrations by control or rOat5-expressing oocytes for 1 h were measured (mean ± S.E.M.; n = 8–10). rOat5-mediated transport was determined by subtracting transport velocity for control oocytes from that for rOat5-expressing oocytes. Each value represents the mean ± S.E.M. (n = 8–10) from one typical experiment of three separate experiments.
Figure 2 shows the properties of E1S transport via rOat5. The cell-associated count of [3H]E1S increased linearly until 2 h of incubation in rOat5-expressing oocytes. This result indicates that rOat5 not only binds but also translocates E1S into the cytoplasm (Fig. 2A). The rate of E1S uptake via rOat5 was not affected by the replacement of extracellular sodium with lithium or choline (Fig. 2B).
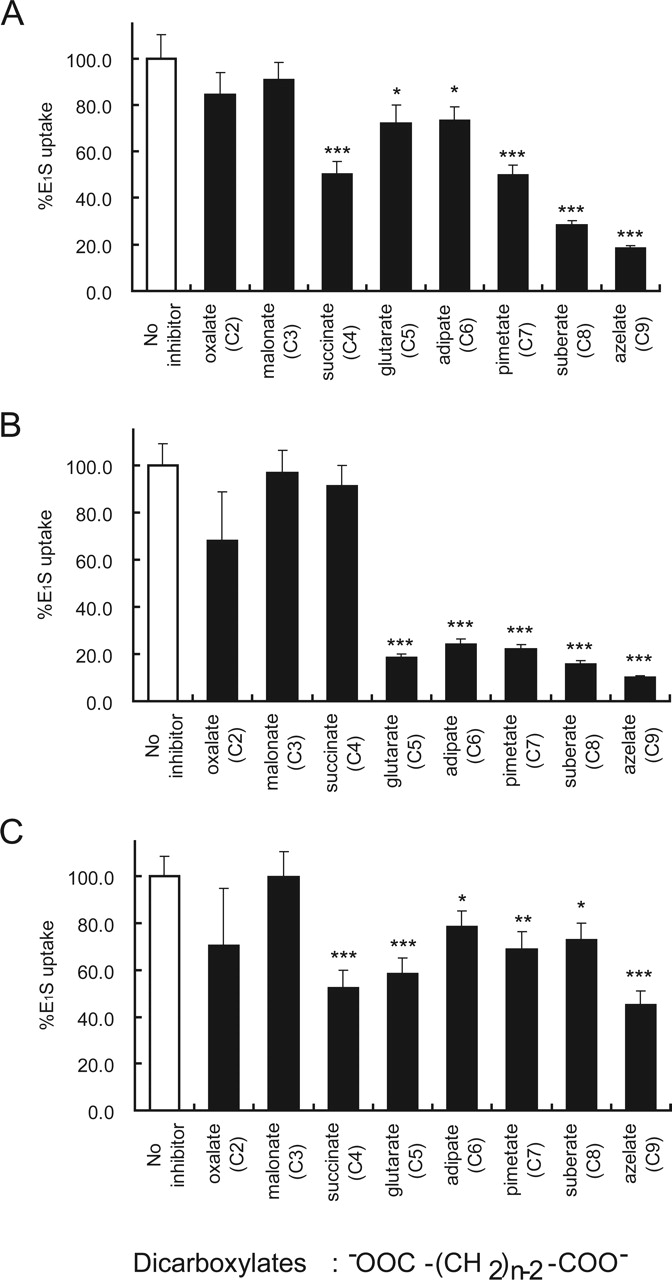
Inhibition of [3H]E1S uptake via rOat5 (A), rOat3 (B), and OAT4 (C) by dicarboxylates. The [3H]E1S concentration used was 100 nM. The inhibitor concentration was 1 mM (gray column). The value was expressed as a percentage of [3H]E1S uptake for 1 h in rOat5-, rOat3-, or OAT4-expressing oocytes in the absence of the inhibitor (open column) (mean ± S.E.M.; n = 8–10). *, p < 0.05; **, p < 0.01; ***, p < 0.001.
We then examined the concentration-dependent uptake of [3H]E1S, [3H]DHEAS, and [3H]OTA via rOat5 (Fig. 3). The rOat5-mediated uptake of these three compounds manifested saturable kinetics and followed the Michaelis-Menten equation. Eadie-Hofstee equation yielded Km values of 18.9 ± 3.9, 2.3 ± 0.2, and 0.34 ± 0.06 μM and Vmax values of 6.6 ± 1.2, 0.25 ± 0.03, and 0.26 ± 0.05 pmol/oocyte/h for E1S, DHEAS, and OTA, respectively, which were determined using the same batch of oocytes.
Recently, we have reported that OAT4, which is expressed at the apical membrane of human renal proximal tubules, mediates organic anion transport in exchange for five-carbon (C5) dicarboxylate glutarate (Ekaratanawong et al., 2004). These results indicate that OAT4 is an organic anion/dicarboxylate exchanger similar to the basolateral membrane isoforms OAT1 (Sekine et al., 1997; Sweet et al., 1997) and OAT3, which have been clarified as organic anion/dicarboxylate exchangers (Bakhiya et al., 2003; Sweet et al., 2003). To investigate the possibility that rOat5 functions as an organic anion/dicarboxylate exchanger and to search the candidate(s) for its counterion, we first examined the interaction of several dicarboxylates with rOat5, although agents that inhibit are not always exchanged. We performed inhibition experiments in which the uptake of 100 nM [3H]E1S was measured in the presence of various dicarboxylates (1 mM) ranging from C2 oxalate to C9 azelate (Ullrich et al., 1987). Among these, C3 to C5 dicarboxylates exist as endogenous organic anions in the cells. rOat5-mediated [3H]E1S uptake was inhibited strongly by succinate (C4) and by all longer dicarboxylates (C7-C9) and moderately by glutarate (C5) and adipate (C6) (Fig. 4A).

trans-Stimulatory effect on rOat5-mediated transport in X. laevis oocytes. A, trans-stimulatory effect on the uptake of [3H]E1S via rOat5. Control (open column) and rOat5-expressing (solid column) oocytes were injected (50 nl) with 50 mM unlabeled succinate (right two lanes) or water (left two lanes) and incubated for 1 h at room temperature. Then, the oocytes were incubated with [3H]E1S (100 nM), and the amount of [3H]E1S accumulated for 1 h was determined. B, trans-stimulatory effect on the efflux of [14C]succinate via rOat5. Control (open column) and rOat5-expressing (hatched column) oocytes were injected (50 nl) with [14C]succinate (∼2 mM). After washing, the oocytes were incubated at room temperature for 1 h and then incubated with 0.1 mM unlabeled E1S (right lanes) or without a substrate (left lanes) and the amount of [14C]succinate effluxed for 1 h was determined. Each value represents the mean ± S.E.M. (n = 8–10) from one typical experiment of two separate experiments. ***, p < 0.001.

Transport of E1S and succinate in S2 cells expressing OATs. Representative studies of the uptake of 100 nM [3H]E1S (A) and 10 μM [14C]succinate (B) in rOat5-expressing S2 cells (S2 rOat5) (closed circles) and in S2 cells transfected with the expression vector pcDNA3.1 alone (S2 mock) (open squares). The cells were incubated for 1 to 15 min at 37°C. C, trans-stimulatory effects of succinate and glutarate on rOat5- and rOat3-mediated E1S transport. The concentrations of preloaded glutarate (gray columns) and succinate (black columns) were 1 mM. Values represent mean ± S.E.M. of two to three independent experiments with four determinations. *, p < 0.05; ***, p < 0.001.
To compare the inhibition profiles of other renal OATs, such as the basolateral membrane isoform rOat3 and the apical membrane isoform OAT4, we also performed inhibition experiments using dicarboxylates for E1S transport in rOat3 and OAT4-expressing oocytes as described above. The basolateral membrane isoform rOat3-mediated [3H]E1S uptake was inhibited strongly by glutarate (C5) and by all longer dicarboxylates (C6–C9) but not by succinate (C4) (Fig. 4B). In contrast, the apical membrane isoform OAT4-mediated [3H]E1S uptake was inhibited strongly by succinate (C4), glutarate (C5), and azelate (C9) and moderately by adipate (C6), pimetate (C7), and suberate (C8) (Fig. 4C).

Inhibition of rOat5-mediated [3H]E1S uptake by various compounds. The rate of [3H]E1S (100 nM) uptake by rOat5-expressing or noninjected oocytes was determined in the absence or presence of 50 μM (gray column), 100 μM (closed column), and 500 μM (open column) inhibitors. The value was expressed as a percentage of [3H]E1S uptake by rOat5-expressing oocytes in the absence of the inhibitor (mean ± S.E.M.; n = 8–10).
Then, we tested the trans-stimulatory effect of succinate and E1S on the uptake and efflux of radiolabeled substrates via rOat5. We injected the unlabeled anionic compounds directly into the oocytes as previously described (Enomoto et al., 2002). The uptake of [3H]E1S was significantly trans-stimulated by unlabeled succinate injected into the oocytes (Fig. 5A), and the efflux of injected [14C]succinate was significantly trans-stimulated by unlabeled E1S in the medium (0.1 mM) (Fig. 5B).
To confirm the succinate transport via rOat5 and the trans-stimulatory effect of succinate on rOat5-mediated E1S transport, we established the cells stably expressing rOat5 (S2 rOat5). The expression of rOat5 was detected by Western blot analysis using the crude membrane fractions from S2 rOat5 cells and not from S2 mock cells (data not shown). Figure 6, A and B, shows the time profiles of the rates of uptake of [3H]E1S (A) and [14C]succinate (B) by S2 rOat5 and S2 mock. Their rates of uptake by S2 rOat5 were greater than those of S2 mock and increased linearly until 2 min of incubation. A similar [14C]succinate uptake by S2 OAT4 cells has recently been identified (N. Anzai, unpublished observation). The uptake of [3H]E1S via rOat5 was significantly trans-stimulated by preloaded succinate (5 mM) (Fig. 6C). In contrast, the uptake of [3H]E1S via rOat3 was trans-stimulated by preloaded glutarate (5 mM), but not by preloaded succinate (5 mM) (Fig. 6C). On the other hand, similar to a previous report on S2 OAT4 cells (Ekaratanawong et al., 2004), the uptake of [14C]succinate was not trans-stimulated by preloaded E1S and the efflux of preloaded [3H]E1S was not stimulated with unlabeled succinate in the medium (5 mM) (data not shown).
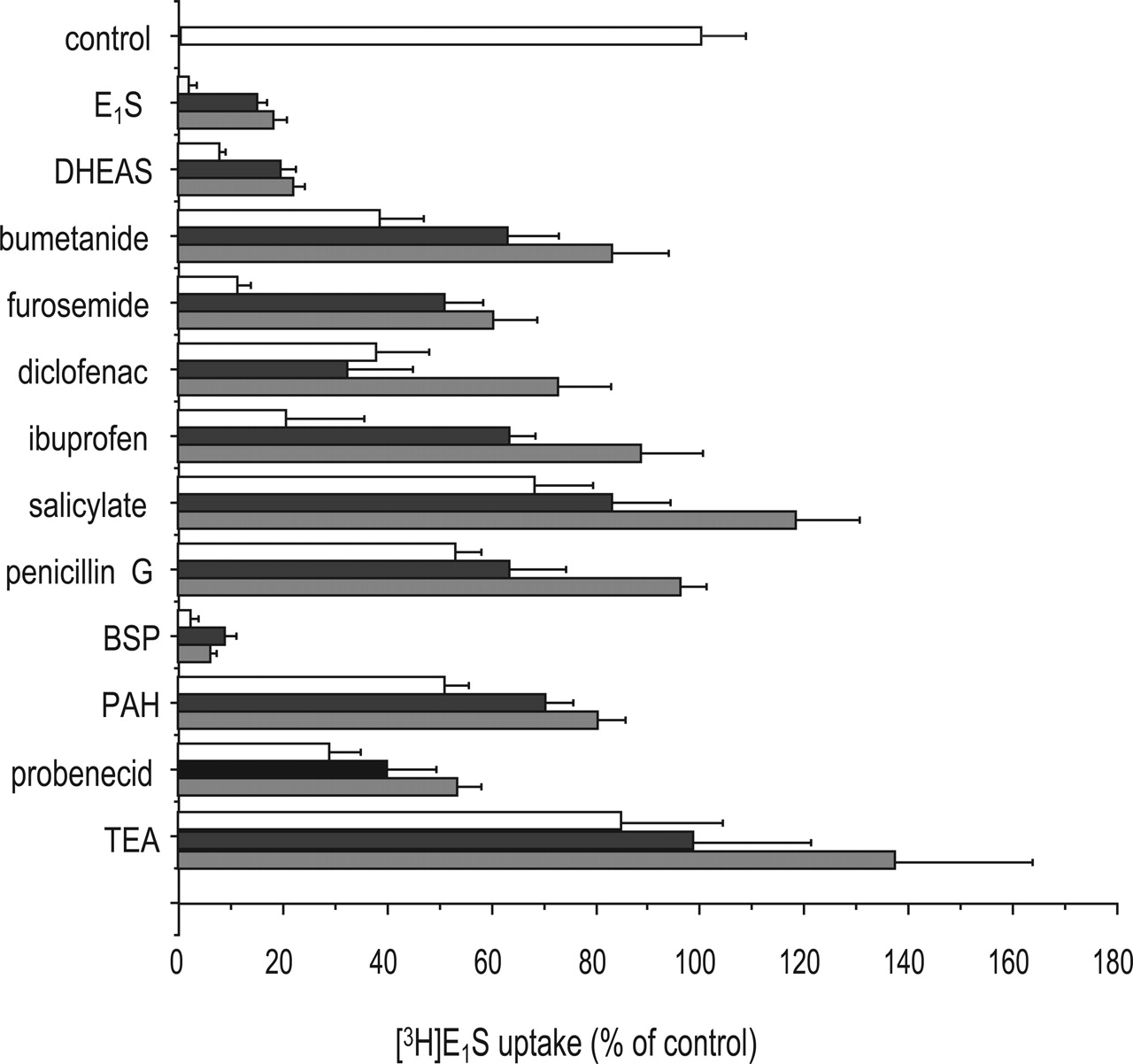
To further investigate the substrate selectivity of rOat5, an inhibition study was performed using rOat5-expressing oocytes. The cis-inhibitory effect of various compounds at 50, 100, and 500 μM on rOat5-mediated [3H]E1S (100 nM) uptake was investigated (Fig. 7). Unlabeled E1S, DHEAS, and sulfobromophthalein at 50 μM each showed a definite inhibitory potency. Bumetanide, furosemide, diclofenac, ibuprofen, and probenecid showed a moderate but dose-dependent inhibitory effect. Salicylate, penicillin G, and PAH showed a much weaker inhibitory effect. In contrast, tetraethylammonium (an organic cation) did not show any inhibitory effect on rOat5-mediated [3H]E1S uptake.
The interaction of rOat5 with several sulfate and glucuronide conjugates was also examined (Fig. 8). Unlabeled E1S, DHEAS, 4-methylumbelliferyl sulfate, and β-estradiol sulfate showed a strong inhibitory effect on the rOat5-mediated uptake of [3H]E1S, whereas p-nitrophenyl sulfate and vinblastine sulfate did not (Fig. 8A). In contrast, we could not find the strong inhibitors in all of the glucuronide conjugates tested, which are the well known substrates for OATPs and MRPs (Fig. 8B).
To determine the localization of rOat5 in the kidney compared with rOat3, we immunostained serial kidney sections with the anti-rOat5 and anti-rOat3 antibodies. First of all, to ascertain the specificity of the anti-rOat5 antibody, crude membrane fractions of transfected HEK293 cells were examined. Single band of ∼65 kDa was detected in HEK293 cells transfected with rOat5 expression vector (Fig. 9A, lane 1), but no band was detected in cells transfected with vector alone (pcDNA3.1) (Fig. 9A, lane 2). We then examined the expression of rOat5 in rat kidney by diaminobenzidine reaction. Low-magnification light microscopy of 2-μm-thick paraffin sections demonstrated that there is a specific immunostaining for rOat5 in proximal tubular cells, mainly in the outer stripe of the outer medulla as well as in the medullary ray of the cortex (Fig. 9B). No immunostaining for rOat5 was detected in the inner medulla and inner stripe of the outer medulla. In contrast, the broad distribution of rOat3 was detected in tubules from the inner medulla to the cortex, as reported previously (Kojima et al., 2002). rOat3 signals in proximal tubular cells were stronger in the cortical labyrinth than in the medullary ray (Fig. 9C). Under a high magnification, rOat5 immunostaining was detected mainly at the apical membrane of proximal straight tubules (S3) (Fig. 9D), whereas rOat3 signals were observed mainly at the basolateral membrane of proximal convoluted tubules (S2) (Fig. 9E). In the corticomedullary junction, the overlapping expressions of apical membrane rOat5 and basolateral membrane rOat3 were detected in some proximal tubular cells (Fig. 9, D and E). The specificity of the anti-rOat5 antibody was confirmed by the reduced immunoreactivity followed by the preincubation of the antibody with the synthetic rOat5 peptide (200 μg/ml) (data not shown).

Inhibition of rOat5-mediated [3H]E1S uptake by sulfate and glucuronide conjugates. The [3H]E1S concentration used was 100 nM. The inhibitor concentration was 100 μM. The value was expressed as a percentage of [3H]E1S uptake in rOat5-expressing oocytes in the absence of the inhibitor (mean ± S.E.M.; n = 8–10).

Immunohistochemical analysis of rOat5 and rOat3 in serial sections of rat kidney. A, Western blot analysis of rOat5 protein. Membrane proteins prepared from HEK293 cells transfected with pcDNA3.1-rOat5 (10 μg) (lane 1) and empty vector (pcDNA3.1) (10 μg) (lane 2) were electrophoresed on a 10% SDS-polyacrylamide gel. Proteins were detected using an enhanced chemiluminescence system. B to E, immunohistochemical labeling of rOat5 and rOat3 by diaminobenzidine reaction of rat kidney tissue cryosections. B, rOat5 was detected in proximal tubules, in the outer stripe of the outer medulla, and in juxtamedullary cortex. C, rOat3 was present in the proximal tubules and tubules in the outer and inner medullae. The apical membrane of proximal tubule was immunostained with the anti-rOat5 antibody (D) and no immunostaining was observed in the basolateral membrane and glomeruli. rOat3 was detected in the basolateral membrane of proximal tubules. E, in some tubules, the overlapping expressions of both rOat3 and rOat5 were observed. *, location of the same renal tubule in serial kidney sections. These figures are representative of typical section samples. Magnifications, ×100 (B, C) and ×400 (D, E).
In addition, we performed RT-PCR analysis to determine the intranephron distributions of rOat5 and rOat3 mRNAs. rOat5 mRNA was expressed strongly in the S2 and S3 segments of the proximal tubule, moderately in glomeruli and CCD, and weakly in MAL. rOat3 mRNA was expressed strongly in the S1 and S2 segments of the proximal tubule, moderately in the S3 segment, and weakly in MAL and collecting ducts. The overlapping expressions of rOat5 and rOat3 were observed in the S2 and S3 segments (Fig. 10).
Discussion
In this study, we identified, for the first time, the four-carbon dicarboxylate succinate (C4) as one of the counterions for the OAT family as shown in Figs. 4A, 5, and 6. It has been established that the classical renal organic anion transport system at the basolateral membrane uses five-carbon dicarboxylate α-ketoglutarate (C5) as a counterion for its organic anion exchange (Burckhardt et al., 2001). The first identified that OAT family members OAT1 and fROAT are PAH/α-ketoglutarate exchangers (Sekine et al., 1997; Sweet et al., 1997; Wolff et al., 1997). Recently, rat and human OAT3s have been shown to be organic anion/α-ketoglutarate exchangers as well (Bakhiya et al., 2003; Sweet et al., 2003). To date, there have been no reports regarding the role of succinate as a counterion for renal organic anion transporters. In a previous study, Youngblood and Sweet (2004) reported that the mOat5-mediated transport was neither cis-inhibited nor trans-stimulated by the dicarboxylate glutarate and succinate. As we previously discussed in our report on OAT4, 500 μM glutarate is not sufficient to inhibit the OAT4-mediated transport and the preincubation of dicarboxylates is not suitable for inducing the trans-stimulatory effect in oocyte system (Ekaratanawong et al., 2004). Therefore, in this study, we used 1 mM succinate, which is 4-fold higher than that used by Youngblood and Sweet, and we directly injected cold succinate into the oocytes instead of using the preincubation method they used for mouse Oat5. These modifications might lead to the identification of the organic anion/succinate exchange property of rOat5.
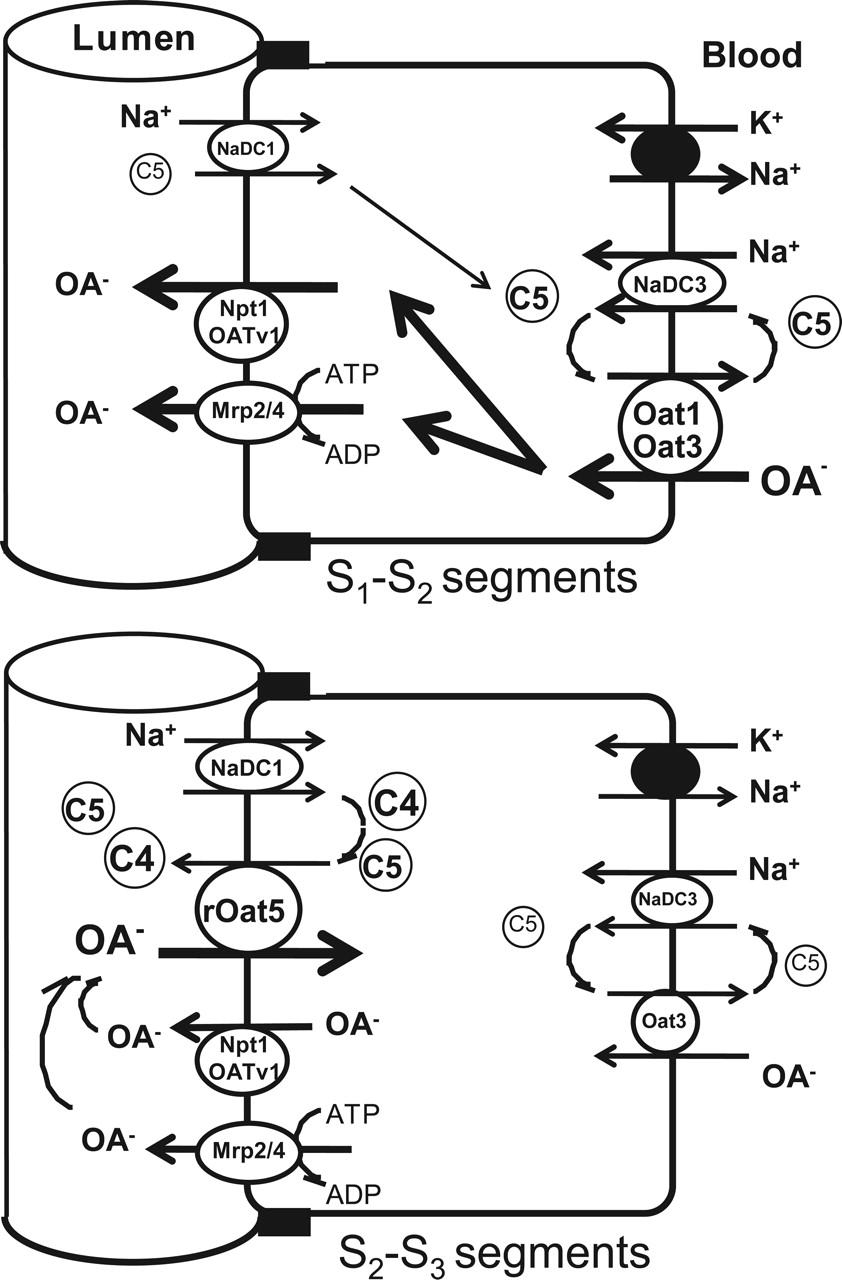
It was reported that glomerular-filtrated α-ketoglutarate is abundantly reabsorbed from the lumen in the early segments of proximal tubules (Ferrier et al., 1985) and that a portion of this reabsorbed α-ketoglutarate is considered to contribute to the basolateral uptake of organic anions via organic anion/α-ketoglutarate exchangers, such as OAT1 and OAT3 (Burckhardt and Pritchard, 2000). However, our previous report indicated that organic anion/α-ketoglutarate exchange activity exists not only in the basolaterally localized OAT1 and OAT3 but also in the apically localized OAT4 of the same proximal tubular cells (Ekaratanawong et al., 2004). As Schmitt and Burckhardt (1993) discussed in the case of bovine BBMVs, it seems unlikely that two exchangers with the same properties existing on both sides of membranes accomplish the vectorial transport of organic anions. However, our current finding that succinate is the most likely counterion for the rOat5-mediated transport may explain this contradiction. To separate or isolate the transport of organic anions at the apical membrane of proximal tubules from that at the basolateral membrane, the apical membrane isoforms of OAT seem to use four-carbon dicarboxylate succinate as a counterion, whereas the basolateral membrane OAT isoforms use five-carbon dicarboxylate as a counterion (Fig. 11). This idea is supported by the results shown in Fig. 4, B and C, that the basolateral membrane isoform rOat3-mediated transport was inhibited strongly by glutarate, but not by succinate, and that the apical membrane isoform OAT4-mediated transport was inhibited by succinate as well as glutarate. It is noteworthy that urate transport via the urate/anion exchanger URAT1 localized at the renal apical membrane was also inhibited by succinate at a millimolar concentration in the X. laevis oocyte system (Enomoto et al., 2002). It has been suggested that the succinate exchange property is a common mechanism for the apical membrane isoforms of OAT family members.
Dicarboxylic and tricarboxylic acids are actively taken up by proximal tubular cells from both peritubular fluid and glomerular filtrate (Simpson, 1983). The transport processes have been assumed to be mediated by two distinct Na+-dicarboxylate pathways, the luminal and the basolateral membrane Na+-dicarboxylate transporters, NaDC1 and NaDC3 (Wright, 1985; Wright and Wunz, 1987). One proposed function of these two transporters is to supply Krebs cycle intermediates, such as citrate, α-ketoglutarate, and succinate, to support the oxidative metabolism in proximal tubular cells. Another function that has been proposed is to sustain the outward dicarboxylate gradient that drives the organic anion uptake into proximal tubular cells. Because α-ketoglutarate is the most abundant within a proximal tubular cell (∼200 μM) and is found in blood circulation with mean plasma level of ∼10 μM (Pritchard, 1995), α-ketoglutarate is considered to be the most likely candidate for the physiological counterion participating in organic anion transport, particularly at the basolateral membrane. However, succinate has recently been identified by He et al. (2004) as a natural ligand for GPR91, an orphan G-protein-coupled receptor in the proximal and distal tubules. They showed that succinate increases blood pressure in mice and that the succinate-induced hypertension involves the renin-angiotensin system. Succinate is also found in blood circulation with a mean plasma level of ∼5 μM (Kushnir et al., 2001). These findings raise the possibility that succinate level surrounding proximal tubular cells, determined by the balance between the uptake of dicarboxylates via NaDCs and its efflux via OATs, is an important factor that regulates renovascular hypertension. Succinate is a prototypical substrate of NaDCs, and it is also reabsorbed in proximal tubules with α-ketoglutarate. It is noteworthy that the apical membrane isoform of this transporter NaDC1 is expressed in the S2 and S3 segments in rats (Sekine et al., 1998a) and that this distribution profile is similar to that of rOat5 (Figs. 9 and 10). These results raise another possibility that rOat5 participates in the dicarboxylates efflux (including succinate) from tubular cells using the outward dicarboxylate gradient in the tubular cells.

Intranephron distribution of rOat5 and rOat3 in rat kidney. rOat5 mRNA was expressed strongly in the S2 and S3 segments of proximal tubules, moderately in glomeruli and CCD, and weakly in MAL (top panel). rOat3 mRNA was expressed strongly in the S1 and S2 segments of proximal tubules, moderately in the S3 segment, and weakly in MAL and collecting ducts (bottom panel).
rOat5 is unique among OATs in several respects. rOat5 is functionally distinct from OAT4 in substrate specificity, affinity, and intranephron localization. First, rOat5 did not transport PAH (data not shown), although one of the common characteristics of the four OAT isoforms (OAT1 to OAT4) identified to date is the transport of PAH, a prototypical substrate for renal organic anion transport. In particular, OAT1 is considered to be a classical PAH transporter because it mediates the high-affinity transport of PAH in exchange for intracellular α-ketoglutarate (Burckhardt et al., 2001). The lack of PAH recognition by rOat5 seems to be one reason that the apical organic anion/dicarboxylate-exchange system was not found by previous classical transport studies using isolated rat renal tubules or brush-border membrane vesicles preparation. Second, the affinity for E1S transport is different between rOat5 and other OAT members. As reported previously, both OAT3 and OAT4 manifested a high affinity for E1S. Km for E1S transport for rOat3 was 2.3 μM (Kusuhara et al., 1999), that for hOAT3 was 3.1 μM (Cha et al., 2001), and that for OAT4 was 1.01 μM (Cha et al., 2000). On the other hand, Km for E1S transport for rOat5 was 18.9 μM, which was approximately 10- to 20-fold higher than that by other OATs (Fig. 3). Third, rOat5 was expressed mainly in the late segments (S2 and S3) of proximal tubules (Figs. 9 and 10). On the other hand, another renal apical membrane OAT isoform OAT4 was expressed throughout the proximal tubules (from S1 to S3) (Ekaratanawong et al., 2004). It is noteworthy that our previous study demonstrated that OAT4 immunoreactivities completely overlap with those of OAT1 and OAT3 in the S1 and S2 segments of proximal tubules (Ekaratanawong et al., 2004), whereas rOat5 immunoreactivity overlapped partly with that of rOat3 (Fig. 9).

Proposed scheme of organic anion (OA-) transport in rodent renal proximal tubules. In the early (S1 to S2) segments of the proximal tubules (top), OAs, including xenobiotics (e.g., PAH), enter into proximal tubular cells via basolateral Oat1 and/or Oat3 using the outward gradient of α-ketoglutarate (C5) created by NaDC3 and they are secreted into the urine via apical efflux transporters, such as Mrp2/4 and Npt1(OATV1). In contrast, in the late (S2 to S3) segments of the proximal tubules (bottom), because the apical membrane rOat5 expression level is high and the basolateral membrane Oat3 expressions level is low, some organic anions, such as steroid sulfates that are glomerular filtrated or tubular secreted or effluxed by Mrp2/4 and/or Npt1(OATV1), are reabsorbed by rOat5 using the outward gradient of succinate (C4) andα-ketoglutarate (C5) generated by NaDC1. Some organic anions may be accumulated into proximal tubular cells and may induce xenobiotic-induced nephrotoxicity.
As described above, rOat5 shows a remote amino acid sequence similarity to OAT4 (47%) and it appears to be different from OAT4. However, rOat5 still shares some common characteristics with other OATs. rOat5, similar to OAT3 and OAT4, and exhibits narrow substrate selectivity for sulfate conjugates (Fig. 8) (Kusuhara et al., 1999; Cha et al., 2000). The inhibition study revealed that the pharmacological profile of rOat5 is similar to that of OAT3 and OAT4 in which rOat5-mediated E1S uptake was inhibited by bumetanide, furosemide, penicillin G, probenecid, and sulfobromophthalein (Fig. 7). Furthermore, the inhibition profile of rOat5 for sulfate and glucuronide conjugates was similar to that of OAT4 (Fig. 8). rOat5 preferentially accepted sulfate conjugates rather than glucuronide conjugates, whereas it did not accept small sulfoconjugates, such as p-nitrophenyl sulfate (molecular weight: 257.3), and large sulfoconjugates, such as vinblastine sulfate (molecular weight: 909.1). Therefore, it will be interesting to determine the common inherent structural traits responsible for such similar substrate selectivities of different transporters.
In summary, we describes the intrarenal localization and the functional characterization of an organic anion transporter rOat5 (Slc22a19). The observations in this study suggest that the renal apical transporter rOat5 functions as an apical backflux pathway (Ekaratanawong et al., 2004) for organic anions (e.g., sulfate conjugates) in the terminal segment of renal proximal tubules, driven by an outward gradient of succinate under physiological conditions (Fig. 11). rOat5 may contribute to selectively reabsorb sulfate conjugates excreted into luminal fluid by the apical organic anion efflux transporters (e.g., MRP2/4 and NPT1/OATV1) in the early segment of renal proximal tubules. Although the relative contribution of rOat5 to renal organic anion and/or dicarboxylate handling needs to be clarified, this study provides new insights into the functional basis and the integration of the apical transport pathway of organic anions, including xenobiotics and endogenous compounds as well as dicarboxylates, such as succinate, in the rat kidneys.
Acknowledgments
We thank A. Toki for technical assistance and Drs. A. Bahn, A. M. Torres, and T. Sekine for helpful discussions. The anti-rOat5 and anti-rOat3 polyclonal antibodies were supplied by Transgenic Inc. (Kumamoto, Japan).
Footnotes
-
This work was supported in part by grants from the Japanese Ministry of Education, Culture, Sports, Science and Technology (grants-in-aid for Scientific Research and High-Tech Research Center), the Science Research Promotion Fund of the Japan Private School Promotion Foundation, Research on Health Sciences Focusing on Drug Innovation from the Japan Health Sciences Foundation, grants-in-aid for Scientific Research from the Japan Society for the Promotion of Science (JSPS), grants-in-aid from the Salt Science Research Foundation (no. 0524), and Health and Labor Sciences Research Grants for Research on Advanced Medical Technology: Toxicogenomics Project. P.J. is a research fellow supported by the Labor Sciences Research Grants for Research on Advanced Medical Technology: Toxicogenomics Project.
-
Part of this work was presented at the 36th Annual Meeting and Scientific Exposition of the American Society of Nephrology, San Diego, CA, October 2003.
-
N.A. and P.J. contributed equally to this work.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.105.088583.
-
ABBREVIATIONS: OAT/Oat, organic anion transporter; PAH, para-aminohippuric acid; rOat5, rat Oat5; BBMV, brush-border membrane vesicle; NPT/Npt, sodium-dependent inorganic phosphate transporter; MRP/Mrp, multidrug resistance-associated protein; OATP/oatp, organic anion-transporting polypeptide(s); MAL, medullarly thick ascending limb; OATV1, voltage-driven organic anion transporter; EST, expressed sequence tag; E1S, estrone-3-sulfate; DHEAS, dehydroepiandrosterone sulfate; OTA, ochratoxin A; NaDC, Na+-dependent dicarboxylate transporter; HEK, human embryonic kidney; PCR, polymerase chain reaction; S1, S2, S3, the first, second, and third segments of proximal tubule, respectively; hOAT, human OAT; D-PBS, Dulbecco's modified phosphate-buffered saline; RT, reverse transcription; CCD, cortical collecting duct.
- Received April 25, 2005.
- Accepted August 1, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}