


Abstract
Methotrexate (MTX) is used in patients with malignant and autoimmune diseases. This drug is primarily excreted unchanged in the urine, and its net excretion occurs via active secretory and reabsorptive processes. We characterized the interaction of MTX with human organic-anion transporting polypeptide transporter (OATP) 1A2, which is expressed in tissues important for MTX disposition and toxicity, such as the intestine, kidney, liver, and endothelial cells of the blood-brain barrier. In Xenopus laevis oocytes expressing OATP1A2, the uptake of the model substrate, estrone-3-sulfate (ES), was enhanced 30-fold compared with uninjected oocytes. MTX uptake in oocytes expressing OATP1A2 was saturable (Km = 457 ± 118 μM; Vmax = 17.5 ± 4.9 pmol/oocyte/60 min) and sensitive to extracellular pH. That is, acidic pHs stimulated MTX uptake by as much as 7-fold. Seven novel protein-altering variants were identified in 270 ethnically diverse DNA samples. Four protein-altering variants in OATP1A2 exhibited altered transport of ES and/or MTX. The common variant, protein reference sequence (p.) Ile13Thr, was hyperfunctional for ES and MTX and showed a 2-fold increase in the Vmax for ES. The common variant, p. Glu172Asp, exhibited reduced maximal transport capacity for ES and MTX. p. Arg168Cys was hypofunctional, and p. Asn277DEL was nonfunctional. Because of its expression on the apical membrane of the distal tubule and in tissues relevant to MTX disposition and toxicity, these findings suggest that OATP1A2 may play a role in active tubular reabsorption of MTX and in MTX-induced toxicities. Furthermore, genetic variation in OATP1A2 may contribute to variation in MTX disposition and response.
MTX, a folate antimetabolite and a weak bicarboxylic organic anion, has been used to treat millions of patients with malignant and autoimmune diseases. Despite its high efficacy, MTX therapy can result in severe toxicity to the kidney, liver, gastrointestinal tract, hematopoietic system, and central nervous system (Goodman et al., 2001). MTX is primarily excreted unchanged in the urine, and its net excretion is a function of active tubular secretion and reabsorption (Bourke et al., 1975; Shen and Azarnoff, 1978; Huang et al., 1979; Hendel and Nyfors, 1984). Because of poor solubility, MTX and its minor metabolite, 7-hydroxymethotrexate (7-OH-MTX), can precipitate in the distal kidney lumen, especially after high doses. This crystalluria can result in nephrotoxicity and delayed excretion, producing high systemic blood levels of MTX (el-Badawi et al., 1996; Perazella, 1999, 2003; Goodman et al., 2001). Accumulation of MTX in blood can then lead to severe system toxicity to other organs.
Organ-specific toxicity of MTX as well as other drugs is often related to high systemic or tissue-specific drug levels. There is evidence that systemic blood levels and tissue-specific levels of MTX are determined by membrane transporters such as the reduced folate carrier (SLC19A1), organic anion transporters, and ATP-binding cassette (ABC) transporters (Hooijberg et al., 2003; Ganapathy et al., 2004; Koepsell and Endou, 2004). Although several transporters that mediate MTX secretion in the proximal tubule have been identified, little is known about the transporters that are responsible for the saturable reabsorption mechanism in the distal tubule of the kidney.
Organic-anion transporting polypeptides (human, OATPs; rodents, Oatps) represent a family of membrane solute carrier proteins that are expressed in a variety of organs important for drug disposition. These proteins mediate the cellular transport of a wide range of amphipathic compounds, including drugs in clinical use and other xenobiotic substances (Hagenbuch and Meier, 2004). Within the OATP family, OATP1A2 (SLCO1A2, also referred as OATP-A or OATP1) has been shown to transport the largest number of structurally diverse compounds, including bile acids, steroid hormones, neutral and anionic peptides, and bulky organic cations (Kullak-Ublick et al., 1995, 2001). Since it is primarily expressed in epithelial cells of the kidney, liver, intestine, and in endothelial cells of the blood-brain barrier (Gao et al., 2000; Su et al., 2004; Lee et al., 2005), OATP1A2 probably plays an important role in tissue-specific disposition, pharmacokinetics, and toxicity of xenobiotics. Recently, OATP1A2 has been localized to the luminal membrane of endothelial cells at the blood-brain barrier and on the apical domain of distal tubule kidney cells, suggesting that this transporter may play a role in drug toxicity in the central nervous system and kidney, as well as in drug disposition (Konig et al., 2005; Lee et al., 2005).
In the present study, we determined that MTX is transported by OATP1A2, suggesting that this transporter is an important determinant of MTX disposition and toxicity. Transport of MTX by OATP1A2 was found to be exquisitely sensitive to extracellular pH but was not affected by the proton gradient across the membrane. Furthermore, we identified seven new protein-altering variants in OATP1A2 in ethnically diverse populations. Four of the 12 protein-altering variants studied, two of which had allele frequencies ≥1%, exhibited changes in the transport of ES and/or MTX compared with OATP1A2 reference. Interestingly, we showed that p. Ile13Thr, a common variant found in 16% of European-Americans and previously reported to have normal transport function, has a 2-fold higher transport of both ES and MTX as a result of a significant increase in Vmax. In addition, p. Glu172Asp, which is found in 2% of European-Americans, exhibited a 3- and 5-fold lower maximal transport capacity for ES and MTX, respectively, compared with OATP1A2. Our data suggest that OATP1A2 plays an important role in the tissue-specific and pH-dependent disposition of MTX in the body and genetic variants in this transporter may have significant pharmacokinetic and toxicological implications.
Materials and Methods
Materials. [3H]ES (57.3 Ci/mmol) was purchased from PerkinElmer Life Sciences (Wellesley, MA), and [3H]MTX (49.6 Ci/mmol) was obtained from Moravek (Brea, CA). All unlabeled chemicals were purchased from Sigma (St. Louis, MO), except for 7-OH-MTX, which was obtained from Schircks Laboratories (Jona, Switzerland). Human embryonic kidney cells (Flp-In-HEK293) and the mammalian expression vector, pcDNA5/FRT, were purchased from Invitrogen (Carlsbad, CA).
Xenopus laevis Oocyte Expression Assay. The original human SLCO1A2 plasmid containing the full-length cDNA in a pCMV expression vector was purchased from Genecopoeia (Germantown, MD). The SLCO1A2 cDNA was subsequently subcloned into the amphibian high-expression vector pOX. Healthy stage V and VI oocytes were harvested from X. laevis (Xenopus One, Dexter, MI) as described previously (Giacomini et al., 1994). The oocytes were injected with 50 to 75 ng of capped cRNA transcribed in vitro using T3 RNA polymerase (mMESSAGE MACHINE T3 kit; Ambion, Austin, TX) from NotI-linearized pOX plasmid containing SLCO1A2 cDNA. Before injection, an aliquot of the cRNA was run on 1% agarose gel to verify that the RNA was not degraded. RNA concentrations were determined using a spectrophotometer. The injected oocytes were incubated in modified Barth's medium [88 mM NaCl, 1 mM KCl, 0.82 mM MgSO4, 0.41 mM CaCl2, 0.33 mM Ca(NO3)2, 2.4 mM NaHCO3, 10 mM HEPES/Tris, pH 7.4, supplemented with 20 mg/l gentamicin, and 50 mg/l tetracycline] at 18°C for 72 h before transport experiments. Groups of seven to nine oocytes were exposed to Na+ buffer containing 20 nM [3H]ES or 0.2 μM [3H]MTX at room temperature for 60 min. For inhibition experiments, folate, leucovorin, MTX, and 7-OH-MTX (1 and 2 mM) were added to Na+ buffer in the presence of 20 nM [3H]ES. Uptake of both ES and MTX was linear for at least 120 min in oocytes expressing OATP1A2. The Na+ buffer contained 100 mM NaCl, 2 mM KCl, 1 mM CaCl2, 1 mM MgCl2, and 10 mM HEPES/Tris[hydroxymethyl]-aminomethane. The pH of the Na+ buffer was adjusted to 7.4 (or any other relevant pH) using Tris[hydroxymethyl]-aminomethane and/or HCl. Uptake was stopped by washing the oocytes five times with ice-cold Na+ free buffer in which Na+ was replaced by choline chloride. Oocytes were lysed individually using 100 μl of 10% SDS, and the radioactivity of each oocyte was then determined.
Saturation Kinetics of [3H]MTX. Oocytes injected with OATP1A2 cRNA were incubated with 0.2 μM [3H]MTX in Na+ buffer, pH 7.4, in the presence of various concentrations of unlabeled MTX at room temperature for 60 min. The kinetic curve for MTX transport by OATP1A2 was generated by fitting the data to a Michaelis-Menten curve by nonlinear regression, using Prism 4 software (GraphPad Software Inc., San Diego, CA). A summary of the kinetic parameters of MTX transport by OATP1A2 is given as mean ± S.E. from three independent experiments.
MTX Transport by OATP1A2 Is Sensitive to Extracellular pH. To test the effect of extracellular pH on OATP1A2-mediated transport of MTX, oocytes injected with OATP1A2 were incubated with 0.2 μM [3H]MTX in Na+ buffer in the presence of decreasing extracellular pH (pHo) at room temperature for 60 min. In addition, to determine whether MTX transport by OATP1A2 was dependent on the proton gradient across the membrane, oocytes expressing OATP1A2 were exposed to the proton ionophore, carbonylcyanide p-trifluoromethoxyphenyl hydrazone (FCCP). Oocytes were preincubated in Na+ buffer, pH 7.4, for 30 min at room temperature. Oocytes were washed three times with Na+ buffer, pH 7.4, and then incubated with 0.2 μM[3H]MTX in Na+ buffer in the absence or presence of 20 μM FCCP at pH 7.4 or pH 5.0 for 60 min. Data are normalized to uptake of MTX by OATP1A2 at pHo = 7.4 and are presented as mean ± S.E. from seven to nine oocytes. Data shown are representative of three independent experiments. p values were determined by comparing uptake in the presence of FCCP with that in the absence of FCCP at each Na+ buffer pH separately, using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance.
Identification of SLCO1A2 Variants. Genomic DNA samples were obtained from unrelated healthy individuals in the San Francisco Bay Area, as described previously (Badagnani et al., 2005). SLCO1A2 variants were identified using automated direct sequencing in an ethnically diverse population of 270 individuals: 80 African Americans, 80 European Americans, 60 Asian Americans (50 Chinese Americans and 10 Japanese Americans), and 50 Mexican Americans. The reference cDNA sequence of SLCO1A2 was acquired from GenBank (http://www.ncbi.nlm.nih.gov, accession no. NM_021094). Primers were designed manually to cover the exons and at least 50 bp of flanking intronic sequence. The primer sequences can be found at http://www.pharmgkb.org. Variant positions were determined relative to the ATG start site and were based on the reference cDNA sequence of SLCO1A2.
Construction of SLCO1A2 Variants for X. laevis Oocyte Studies. OATP1A2 cDNA (GenBank accession no. NM_021094) was subcloned into pOX to obtain OATP1A2 reference. The sequence of OATP1A2 reference corresponds to the OATP1A2 coding region that had the highest frequency in all ethnic groups. Each of the 12 protein-altering variants in OATP1A2 was constructed by site-directed mutagenesis of the OATP1A2-reference-containing plasmid, using the QuikChange mutagenesis protocol (Stratagene, La Jolla, CA).
Functional Studies of SLCO1A2 Variants. Oocytes expressing reference or variant OATP1A2 were incubated in Na+ buffer, pH 7.4, containing 20 nM [3H]ES or 0.2 μM [3H]MTX at room temperature for 60 min. The data are presented as percentage of ES or MTX uptake by OATP1A2 reference at pH 7.4. Each value represents the mean ± S.E. from seven to nine oocytes. Data are shown as a summary of three independent experiments. p values were determined in each of three experiments by comparing variant uptake with OATP1A2 reference for each substrate using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance. The maximum transport rate (Vmax) of ES by OATP1A2 reference and the common protein-altering variant, p. Glu172Asp, was characterized in oocytes injected with varying concentrations of OATP1A2 or variant cRNA. The data shown are representative of three independent experiments and were fitted by nonlinear regression analysis using GraphPad Prism 4 software. p values were determined by comparing uptake of p. Glu172Asp with that of OATP1A2 at each cRNA concentration using the unpaired Student's t test with p < 0.05 as the criterion of significance. In addition, kinetics studies of ES interaction with OATP1A2 and the common protein-altering variants, p. Ile13Thr and p. Thr668Ser, were performed in transiently transfected HEK293 cells. The plasmid containing human SLCO1A2 in pSPORT1 was a generous gift from Dr. B. Stieger (University of Zurich, Zurich, Switzerland). This SLCO1A2 clone has identical open reading frame cDNA sequence and amino acid sequence to the SLCO1A2 clone used in X. laevis oocyte expression assays. The SLCO1A2 cDNA was subsequently subcloned into the mammalian expression vector, pcDNA5/FRT. HEK293 cells were grown and propagated as described previously (Erdman et al., 2006). Cells were plated at 3 × 105 cells/well on poly-d-lysine 24-well plates (Becton Dickinson, Bedford, MA) 24 h before transfection. Transfection was performed by incubating 1 μg/well plasmid DNA with 3 μl of the lipid vehicle Lipofectamine 2000 (Invitrogen). The transfection mixture was replaced with fresh media after 24 h. Uptake experiments were performed 48 h after transfection. Uptake of ES was linear for at least 2 min. HEK293 cells transiently transfected with pcDNA5/FRT, OATP1A2, p. Ile13Thr, and p. Thr668Ser were incubated with 20 nM [3H]ES in Na+ buffer at pH 7.4 in the presence of various concentrations of unlabeled ES for 2 min at 37°C. The kinetic curves were generated by fitting the data to a Michaelis-Menten curve fit, using GraphPad Prism 4 software. A summary of the kinetic parameters of ES transport by OATP1A2 reference and variants is given as mean ± S.E. from three independent experiments. p values were determined by comparing Km and Vmax values for each variant to OATP1A2 using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance.

MTX and its metabolite, 7-OH-MTX, are transported by OATP1A2. A, inhibition of ES uptake by folates (leucovorin and folate) and folate antagonists (MTX and 7-OH-MTX) in X. laevis oocytes expressing OATP1A2. ES uptake by uninjected oocytes and oocytes expressing OATP1A2 is shown in the white and black bars, respectively. Oocytes were incubated for 60 min with [3H]ES (20 nM) in the absence or presence of various concentrations of unlabeled compounds. Uptake is expressed as percentage of ES uptake by OATP1A2 at pH 7.4. p values were determined by comparing ES uptake in the presence of inhibitors to OATP1A2-mediated ES uptake in the absence of inhibitors using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance. B, kinetics of MTX transport in X. laevis oocytes expressing OATP1A2. Oocytes were incubated for 60 min with [3H]MTX (0.2 μM) in the presence of various concentrations of unlabeled MTX. Inset, kinetic parameters of MTX transport by OATP1A2 were determined by fitting the data using nonlinear regression analysis and are shown as mean ± S.E. from three independent experiments. Data are representative of experiments carried out with three different batches of oocytes. Each value represents the mean ± S.E. from seven to nine oocytes.
Results
OATP1A2 Interacts with the Folate Antagonists, MTX and 7-OH-MTX. The role of OATP1A2 in the cellular transport of folates and folate antagonists was examined by inhibiting ES uptake in oocytes expressing OATP1A2. Initial studies demonstrated that uptake of the model substrate, ES, and MTX was enhanced 30- and 3-fold, respectively. As shown in Fig. 1A, MTX and 7-OH-MTX inhibited ES transport by 70 and 25%, respectively. On the other hand, folate and leucovorin, a drug used clinically to rescue normal cells from the cytotoxic effects of MTX, did not inhibit ES uptake, despite being structurally similar to MTX. Structure activity studies to examine the relationship between various functional groups and interaction with OATP1A2 are needed to identify the pharmacophores that are essential or preferred for interaction with OATP1A2. ES and MTX transport in oocytes expressing OATP1A2 was linear for at least 120 min (data not shown), and uptake times of 60 min were, therefore, used in subsequent experiments. The initial rate of uptake of MTX by OATP1A2 was saturable (Fig. 1B), with a mean Km of 457 ± 118 μM and Vmax of 17.5 ± 4.9 pmol/oocyte/60 min.

Transport of MTX by OATP1A2 is sensitive to extracellular pH. A, uptake of [3H]MTX (0.2 μM) in X. laevis oocytes expressing OATP1A2 (black squares) or uninjected oocytes (gray inverted triangles) as a function of decreasing extracellular pH. B, uptake of [3H]MTX (0.2 μM) in X. laevis oocytes expressing OATP1A2 (black bars) or in uninjected oocytes (white bars) in the absence or presence of 20 μM FCCP at extracellular pH 7.4 and 5.0. Oocytes were preincubated for 30 min with Na+ buffer at pH 7.4. Transport of MTX was measured in Na+ buffer over 60 min. Uptake is expressed as percentage of MTX uptake by OATP1A2 injected oocytes at pH 7.4. The pH of the Na+ buffer was adjusted with Trizma and HCl. Data are representative of three independent experiments. Each value represents the mean ± S.E. from seven to nine oocytes.
Transport of MTX by OATP1A2 Is Sensitive to pHo. To characterize the mechanism of transport of MTX by OATP1A2 in tissues, such as the kidney and intestine, where luminal pH is known to vary, MTX uptake by OATP1A2 was measured at pHs ranging from 7.4 to 5.0. MTX transport significantly increased at acidic pH in oocytes expressing OATP1A2 (Fig. 2A). The initial rate of MTX uptake in oocytes expressing OATP1A2 was 5- to 7-fold higher at pH 5.0 (0.606 ± 0.0289 pmol/oocyte/60 min) compared with pH 7.4 (0.0811 ± 0.0046 pmol/oocyte/60 min). Similar findings were obtained with ES, where uptake was enhanced by 2- to 4-fold at pH 5.0 compared with pH 7.4 (data not shown). To examine whether the increase in MTX transport by OATP1A2 at pH 5.0 was due to the H+ gradient across the membrane, the proton ionophore, FCCP, was used to collapse the pH gradient. The initial rate of uptake of MTX at pH 5.0 in oocytes expressing OATP1A2 and preincubated in pH 7.4 buffer was not significantly decreased by FCCP (20 μM) (Fig. 2B). We were unable to determine the effect of extracellular pH on the kinetics of MTX transport by OATP1A2 because MTX was insoluble at concentrations (≥500 μM at acidic pH. Altogether, these data indicate that OATP1A2-mediated uptake of MTX is sensitive to extracellular pH but that the proton gradient across the membrane does not appear to be the driving force for MTX transport by OATP1A2.
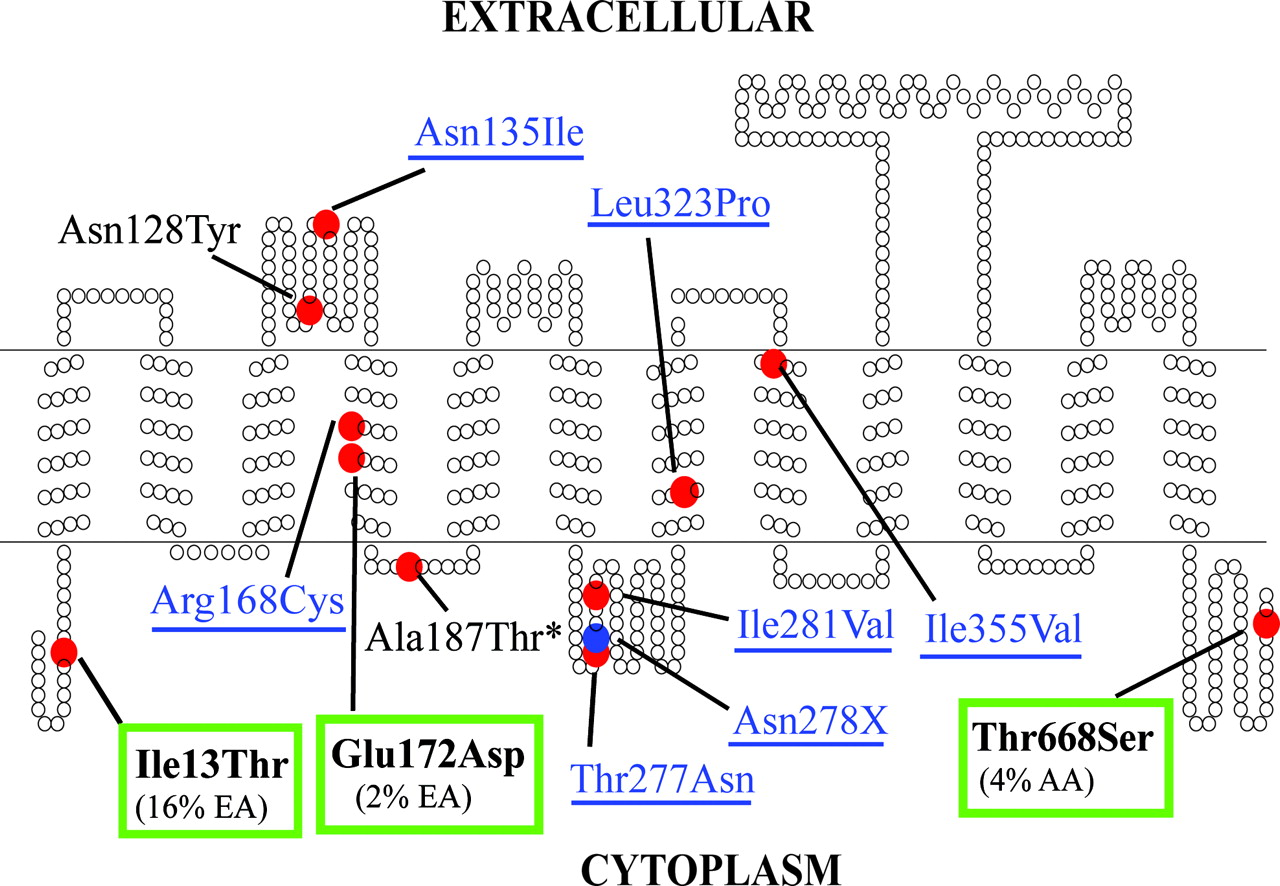
Secondary structure of OATP1A2 showing the location of the 12 protein-altering variants. The transmembrane topology diagram was rendered using TOPO (S.J. Johns, University of California, San Francisco, CA; and R. C. Speth, Washington State University, Pullman, WA), transmembrane protein display software available at the University of California San Francisco Sequence Analysis Consulting Group web site (http://www.sacs.ucsf.edu/TOPO/topo.html). Nonsynonymous variants are identified by the red circles, whereas the deletion variant (Asn278X) is shown in the blue circle. The seven newly identified protein-altering variants are labeled in blue and underlined. The asterisk denotes the previously identified variant that was not found in our resequencing project. The three most common nonsynonymous variants of OATP1A2, which were kinetically characterized, are in boldface and boxed in green.
Genetic Variation in Human SLCO1A2. Genetic variants of SLCO1A2 were identified as part of a large study, the Pharmacogenetics of Membrane Transporters project, which entails identifying variants in membrane transporter genes that may lead to differences in drug response and disposition. The 14 coding exons of SLCO1A2 and 50 to 200 bp of flanking intronic sequence were screened in a collection of 270 ethnically diverse genomic DNA samples. The data are posted in the National Institutes of Health Pharmacogenetics and Pharmacogenomics Knowledgebase, http://www.pharmgkb.org. Forty-one variants were identified; of these, 11 altered the protein sequence of OATP1A2. As shown in Fig. 3 and Table 1, the majority of nonsynonymous variants were found in intracellular or extracellular loops (seven of 11). Through this resequencing effort, seven of 11 protein-altering variants were newly identified (c. 404 A>T; c. 502 C>T; c. 830 C>A; c. 833 A>-; c. 841 A>G; c. 968 T>C; c. 1063 A>G). Six nonsynonymous variants (c. 38T>C; c. 382A>T; c. 404A>T; c. 516 A>C; c. 1063 A>G; c. 2003 C>G) had allele frequencies of ≥1% in at least one ethnic group (Table 1). c. 38 T>C (p. Ile13Thr) was found in all populations sampled, except for Asian Americans, with a particularly high allele frequency, especially in the European-American sample (16.3%). c. 516 A>C was present exclusively in the European American (1.9%) and Mexican American (2%) samples. This variant, which was found in TMD 4 (Fig. 3), has been reported previously to have an allele frequency of 5.3% in European Americans (Lee et al., 2005). c. 2003 C>G was found in the African American population at a high allele frequency (4.4%). Only one protein-altering variant, c. 841A>G, was observed in Asian Americans. c. 833 A>- (p. Asn278DEL) resulted in the early truncation of the OATP1A2 protein and retained only the first six transmembrane domains. Five of the 11 protein-altering variants, c. 502 C>T, c. 830 C>A, c. 833 A>-, c. 841 A>G, and c. 968 T>C, were singletons, meaning that they were found on only one of 540 chromosomes analyzed. The previously identified singleton variant, c. 559 G>A, was not observed in our genomic DNA samples (Lee et al., 2005). In our resequencing effort, we were able to identify 11 protein-altering variants in OATP1A2, only four of which, c. 38 T>C, c. 382 A> T, c. 516 A>C, and c. 2003 C>G, had been identified previously in a screen of 78 DNA samples from European- and African-American populations (Lee et al., 2005). Our data indicate that there is significant genetic variation in OATP1A2, with three protein-altering variants, p. Ile13Thr, p. Glu172Asp, and p. Thr668Ser, present at high allele frequencies in populations.
Summary of protein-altering variants in OATP1A2 identified in ethnically diverse populations Data are available at http://www.pharmgkb.org. The seven newly identified protein-altering variants are shown in italics. The four variants that exhibit a change in transport function compared with reference OATP1A2 are shown in boldface.

Two common (p. Ile13Thr and p. Glu172Asp) and two rare (p. Arg168Cys and p. Asn278DEL) protein-altering variants of OATP1A2 show altered transport function of ES and/or MTX. Uptake of 20 nM [3H]ES (white bars) or 0.2 μM [3H]MTX (black bars) in oocytes expressing OATP1A2 reference or the 12 protein-altering variants. Transport was measured in Na+ buffer at pH 7.4 over 60 min. Uptake is expressed as percentage of ES and MTX uptake, respectively, by OATP1A2-injected oocytes at pH 7.4. Data shown are a summary of three independent experiments and are expressed as mean ± S.E. p values were determined in each of three experiments by comparing variant uptake with OATP1A2 reference for each substrate using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance.
Functional Alterations in ES and MTX Transport by Protein-Altering Variants in OATP1A2. The 12 protein-altering variants of OATP1A2 were constructed by site-directed mutagenesis using the OATP1A2 reference template and were screened for functional activity with the organic anions, ES and MTX, in X. laevis oocytes. In our initial functional screens, 4 of the 12 protein-altering variants, p. Ile13Thr, p. Arg168Cys, p. Glu172Asp, and p. Asn278DEL, exhibited significant (p < 0.01) altered transport of ES and/or MTX compared with OATP1A2 reference (Fig. 4). Interestingly, the common variant p. Ile13Thr, which was previously reported to have a similar function as OATP1A2 reference (Lee et al., 2005), showed approximately a 2-fold increase (ES, 168.4 ± 27.9%; MTX, 174.0 ± 13.6%) in the transport of both organic anions compared with OATP1A2 (ES, 100 ± 7.6%; MTX, 100 ± 5.6%). The common variant p. Glu172Asp (MTX, 59.1 ± 5.3%) and the singleton variant p. Arg168Cys (MTX, 60.4 ± 5.3%) exhibited approximately 40% reduction in the uptake of MTX. The singleton variant p. Asn278DEL, which lacks the last 6 of 12 transmembrane domains, was nonfunctional for both organic anions (ES, 4.4 ± 0.2%; MTX, 24.7 ± 2.8%). In contrast to a previous report (Lee et al., 2005), p. Asn135Ile transported ES and MTX similarly to OATP1A2 reference. Following these initial tests for function, the three most common protein-altering variants in OATP1A2, p. Ile13Thr, p. Glu172Asp, and p. Thr668Ser, were expressed in HEK293 cells to further examine their concentration-dependent transport kinetics of ES. As shown in Fig. 5A and Table 2, cells expressing p. Ile13Thr exhibited a 2-fold increase (p < 0.05) in the Vmax (258.3 ± 32.7 pmol/mg protein/2 min) for ES, whereas the Km (13.1 ± 2.5 μM) was not significantly different compared with cells expressing OATP1A2 (Vmax = 127.0 ± 20.3 pmol/mg protein/2 min; Km = 13.6 ± 3.2 μM). There were no significant differences in the interaction affinity and maximal transport rate of ES between p. Thr668Ser and OATP1A2. Because p.Glu172Asp was nonfunctional when expressed in HEK293 (data not shown), the kinetic studies were performed in X. laevis oocytes. In RNA dose-response studies, the common protein-altering variant, p. Glu172Asp, showed differences in the maximal transport rate, Vmax, of ES compared with OATP1A2 (Fig. 5B). In this approach, oocytes are injected with increasing amounts of reference or variant RNA (10-150 ng). Injecting high doses of RNA allows maximal expression of the transporter protein and controls for RNA degradation that can occur during injection (Werner et al., 1990; Due et al., 1995). The RNA dose-response curve for p. Glu172Asp was significantly lower than that of OATP1A2 (p < 0.01) and p. Glu172Asp exhibited a 3-fold lower maximal transport capacity for ES (Fig. 5B). We also observed a similar reduction, approximately 5- to 7-fold, in the maximal transport capacity of p. Glu172Asp for MTX without any significant changes in the interaction affinity (data not shown). In contrast to many transporters, GFP tagging of OATP1A2 (at either the N or C terminus) produces a nonfunctional protein and other methods to detect the protein via tagged epitopes using confocal imaging have been unsuccessful (I. Badagnani and K. M. Giacomini, unpublished data). Thus, although differences in Vmax have been observed between OATP1A2 and the common variants, p. Ile13Thr and p. Glu172Asp, we do not know whether these differences can be explained by changes in cell surface expression. Altogether, these data suggest that OATP1A2 exhibits significant functional variation with respect to ES and MTX.
Summary of interaction kinetics of ES with OATP1A2 and the common protein-altering variants, p. Ile13Thr and p. Thr668Ser Uptake of [3H]ES (20 nM) was measured in the presence of various concentrations of unlabeled ES in Na+ buffer at pH 7.4 over 60 min. Each value represents the mean ± S.E. from three independent experiments. Transport data were fitted using nonlinear regression analysis. Significant differences in the kinetics parameters of the variant proteins compared to OATP1A2 reference are shown in boldface. p values were determined by comparing Km and Vmax values for each variant with OATP1A2 using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance.
Discussion
In the present study, we provide a comprehensive characterization of the interaction of MTX with OATP1A2, which is highly expressed in tissues important for MTX disposition and toxicity. Our data demonstrate for the first time that the bicarboxylic organic anion, MTX, and its minor metabolite, 7-OH-MTX, interact with OATP1A2 (Fig. 1, A and B). Because OATP1A2 is expressed on the apical surface of distal tubule cells, our studies suggest that OATP1A2 may play a role in the saturable reabsorption of MTX in the distal tubule (Liegler et al., 1969; Hendel and Nyfors, 1984; Lee et al., 2005). Concentrations of MTX in the distal tubule may be 100 times greater than in the plasma; therefore, a Km of 457 ± 118 μM is consistent with expected biological concentrations in the distal tubule, which could range from 10 to 200,000 μM depending on luminal pH (Christophidis et al., 1981; Hendel and Nyfors, 1984; Ferrazzini et al., 1991; Goodman et al., 2001; Chladek et al., 2002). Thus, OATP1A2 may play a role in controlling the levels of MTX and its minor metabolite in the distal tubule and may be an important determinant of its toxicity secondary to crystalluria (el-Badawi et al., 1996; Perazella, 1999, 2003).

Kinetics of interaction of ES with OATP1A2 and its three common protein-altering variants. A, interaction kinetics of [3H]ES in HEK293 cells transiently transfected with OATP1A2 reference (black square) and the common protein altering variants, p. Ile13Thr (red triangle) and p. Thr668Ser (green circle). Uptake of [3H]ES (20 nM) was measured in the presence of various concentrations of unlabeled ES in Na+ buffer at pH 7.4 over 2 min. Data are representative of three independent experiments. Each value represents the mean ± S.E. from three wells. Transport data were fitted using nonlinear regression analysis. p values were determined by comparing kinetic parameters for each variant with OATP1A2 reference using one-way ANOVA and Tukey's multicomparison test with p < 0.05 as the criterion of significance. B, RNA dose-dependent uptake of ES by OATP1A2 reference (black square) and the common protein altering variant, p. Glu172Asp (blue diamond), in X. laevis oocytes. Uptake of [3H]ES (20 nM) was measured in oocytes injected with 18.75, 37.5, 75, or 150 ng of reference or variant cRNA. Transport was measured in Na+ buffer at pH 7.4 over 60 min. Data are representative of three independent experiments. Each value represents the mean ± S.E. from seven to nine oocytes. Transport data were fitted using nonlinear regression analysis. p values were determined by comparing uptake of p. Glu172Asp with that of OATP1A2 at each cRNA concentration using the unpaired Student's t test with p < 0.05 as the criterion of significance.
Because urine pH may vary, particularly in the distal tubule, we determined the effect of extracellular pH on the interaction of MTX with OATP1A2 and observed that MTX uptake by OATP1A2 is extremely sensitive to extracellular pH (Fig. 2A). Uptake of MTX is increased by as much as 7-fold at pH 5.0 compared with pH 7.4. The apparent pH sensitivity of OATP1A2 might reflect the ionization state of MTX rather than a specific effect of pH on the carrier. Because MTX has a pKa of 4.8 and 5.5, the majority of the drug exists as an anion at both pH 7.4 and 5.0. Thus, it is possible that OATP1A2 preferentially transports the monocarboxylic form of MTX resulting in enhanced MTX uptake at acidic pH, where the monocarboxylic form predominates over the bicarboxylic acid form. It is also possible that the acidic extracellular pH may enhance MTX transport by OATP1A2 by affecting the protonation state of amino acid side chains on the extracellular loops and/or extracellular-facing portions of transmembrane domains of OATP1A2. We also observed a pH-dependent transport of ES by OATP1A2, indicating that increased transport at acidic pH is a general property of this transporter (data not shown). Because ES has a pKa of 2 (i.e., predominantly monoanionic at pH 5.0), and transport of ES by OATP1A2 was enhanced 2- to 4-fold at pH 5.0, it can be concluded that the pH-dependent transport of organic anions by OATP1A2 is probably dependent, at least in part, on the protonation state of this carrier protein. Irrespective of mechanism, the finding that MTX transport is enhanced in acidic pHs has important clinical implications in the absorption and disposition of MTX since the luminal pH of the intestine and kidney, where OATP1A2 is expressed, ranges from 4 to 7 and 4.5 to 8, respectively. Interestingly, another human OATP transporter, OATP2B1, which is highly expressed on the apical domain of enterocytes, has been recently shown to be sensitive to extracellular pH, but unlike OATP1A2, its pH-dependent transport relies on the proton gradient (Kobayashi et al., 2003; Nozawa et al., 2004).
Interindividual diversity in MTX efficacy and toxicity is a well recognized obstacle in the successful clinical management of patients with malignant and autoimmune diseases. Significant genetic and functional variation in drug uptake transporters exists and may contribute to variation in MTX disposition and toxicity (Tirona et al., 2001; Leabman et al., 2002; Shu et al., 2003; Gray et al., 2004; Badagnani et al., 2005; Owen et al., 2005; Erdman et al., 2006). In the present study, we characterized the interaction of MTX with genetic variants in OATP1A2. The extent of genetic variation in OATP1A2 seems consistent with other drug uptake transporters, such as organic anion transporter 3 and organic cation transporter 1 and 2 (Leabman et al., 2002; Shu et al., 2003; Erdman et al., 2006). In comparison with the study of Lee et al., we identified a greater number of variants in OATP1A2 including a number of novel variants, two of which, c. 404 A>T and c. 1063 A>G, had allele frequencies of 1.3 and 1.9% in the African and European American samples, respectively (Fig. 3; Table 1). In addition, we found that the allele frequency of c. 516 A>C was 1.9% in European Americans instead of 5.3% as previously reported (Lee et al., 2005). The reason for discrepancies between our study and that of Lee et al. may be due to differences in sample sets and methods used in screening. Lee et al. used single-strand conformational polymorphism analysis to initially screen DNA samples from 46 European Americans and 32 African Americans for variants in OATP1A2 (Lee et al., 2005). Additional DNA samples (n = 302) seemed to be used for estimation of allele frequencies. In contrast, we screened all 270 DNA samples from four ethnic groups by automated direct sequencing analysis. By initially screening a larger sample set in a greater number of ethnic populations, we discovered a greater number of variants.
Functional studies with the 12 protein-altering variants of OATP1A2 showed that four of these variants, p. Ile13Thr, p. Arg168Cys, p. Glu172Asp, and p. Asn278DEL, exhibited altered transport of ES and/or MTX (Fig. 4). p. Arg168Cys was hypofunctional, whereas p. Asn278DEL was nonfunctional. Interestingly, we determined that p. Ile13Thr, which is found at allele frequencies of 16% in European Americans, exhibited 2-fold higher transport of both ES and MTX (Fig. 4). It was determined in kinetic studies that the increased transport of ES by p. Ile13Thr was due to a 2-fold greater transport capacity (i.e., Vmax) with no significant effect on the interaction affinity (i.e., Km) (Fig. 5A; Table 2). Based on these findings, we can speculate that in vivo p. Ile13Thr could result in increased reabsorption of MTX from the distal tubule leading to increased exposure to MTX and subsequent systemic toxicity. However, p. Ile13Thr had been previously reported by Lee et al. (2005) to transport ES similarly to OATP1A2. The difference between our results and those of Lee et al. may be explained by differences in experimental conditions. Lee et al. used HeLa cells infected with vaccinia, whereas we used oocytes. In the study of Lee et al., ES uptake was enhanced approximately 3-fold at 30 min, a time at which ES uptake was no longer linear; therefore, differences in ES uptake between reference and variant OATP1A2 transporters may not have been detected. It is also possible that expression in oocytes may produce differences in transport of both ES and MTX between p.Ile13Thr and reference OATP1A2. To determine whether this was an artifact of the oocyte expression system, we expressed the transporter in mammalian cells and, similarly, obtained a 1.5-2-fold greater uptake and maximal transport capacity of ES by p. Ile13Thr compared with OATP1A2. These data suggest that our results demonstrating a greater uptake of ES by p. Ile13Thr are consistent between mammalian and amphibian expression systems.
p. Glu172Asp, which has an allele frequency of 1.9% in European-Americans, exhibited 40% reduced uptake of MTX (Fig. 4). We determined that the mechanism for the altered transport function of this variant was ascribed to a decrease in the maximal transport rate for both ES and MTX of 3- and 5-fold, respectively, compared with OATP1A2 (Fig. 5B). Based on these findings, we can speculate that in vivo p. Glu172Asp could result in lower reabsorption of MTX and higher MTX accumulation in the lumen of the distal tubule leading to crystalluria and subsequent nephrotoxicity. Our data are consistent with the study of Lee et al. (2005) in which p. Glu172Asp was shown to have significant loss of cell surface expression compared with OATP1A2. Altogether, the genetic and functional analyses of OATP1A2 presented in this study indicate that genetic variation in OATP1A2 may play an important role in MTX disposition and toxicity.
In summary, our studies suggest that protein-altering variants in OATP1A2 may play an important role in interindividual variation in drug disposition and toxicity. Because MTX therapy can result in severe toxicity in the kidney, liver, gastrointestinal tract, and brain, where OATP1A2 is expressed (Goodman et al., 2001; Hagenbuch and Meier, 2004), clinical studies of genetic variants in OATP1A2 are important to determine their significance to MTX therapy.
Acknowledgments
We thank Jennifer Gray, Ryan Owen, and Thomas Urban for helpful discussions.
Footnotes
-
This work was supported by the National Institutes of Health Grants GM61390 and GM36780 and by the General Clinical Research Center at San Francisco General Hospital, funded by the National Center for Research Resources, National Institutes of Health (Grant MO1-RR00083-42). I.B. was supported in part by a National Science Foundation Graduate Research Fellowship.
-
Data in this manuscript were deposited in the Pharmacogenetics and Pharmacogenomics Knowledgebase (www.pharmgkb.org). Some parts of this study were presented as an abstract; Interaction of methotrexate with SLCO1A2 (OATP-A) and its 12 protein-altering variants: clinical implications for delayed excretion and nephrotoxicity; International Society for the Study of Xenobiotics Annual Meeting; 2005 October 23-27; Maui, HI. International Society for the Study of Xenobiotics, Washington, DC.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.104364.
-
ABBREVIATIONS: MTX, methotrexate; 7-OH-MTX, 7-hydroxymethotrexate; SLC, solute carrier; OATP, organic-anion transporting polypeptide; ES, estrone-3-sulfate; p., protein reference sequence; HEK293, human embryonic kidney cell line; cRNA, complementary RNA; FCCP, carbonylcyanide p-trifluoromethoxyphenyl hydrazone; ANOVA, analysis of variance; c., coding DNA reference sequence; TMD, transmembrane domain.
- Received March 10, 2006.
- Accepted May 11, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}