


Abstract
The aim of this study was to investigate the functional importance of multidrug resistance-associated protein (Mrp)3/Abcc3 and Mrp4/Abcc4 in the pharmacokinetics of methotrexate. Compared with the corresponding wild-type mice, the plasma concentrations of methotrexate given orally were similar in Abcc4-/- mice and were significantly lower in Abcc3-/- mice. Pharmacokinetic parameters related to hepatobiliary transport were determined under steady-state conditions in wild-type and Abcc3-/- mice that were given a constant intravenous infusion of methotrexate. The biliary clearance, based on the plasma concentration, was 1.6-fold greater in Abcc3-/- mice than in wild-type mice (23 and 15 ml/min/kg, respectively, P < 0.05). Because the basolateral uptake and canalicular efflux clearances of methotrexate were similar in wild-type and Abcc3-/- mice, this result suggests that the basolateral efflux clearance of methotrexate is decreased in the liver of Abcc3-/- mice. Furthermore, a lower fraction of absorption of methotrexate (Fa Fg) was suggested in Abcc3-/- mice (0.49 and 0.29 in wild-type and Abcc3-/- mice, respectively). The mucosal-to-serosal transport rate of methotrexate, determined in vitro using everted sacs, was highest in the duodenum and was significantly decreased in Abcc3-/- mice compared with wild-type mice. This is ascribed to the reduced intrinsic efflux clearance of methotrexate across the serosal membrane (22 and 5.3 μl/min/sac in wild-type and Abcc3-/- mice, respectively, P < 0.05). These results suggest that Mrp3 mediates basolateral efflux of methotrexate in the liver and duodenum, thereby serving to increase systemic exposure, whereas Mrp4 is likely to play only a limited role in the systemic methotrexate exposure.
Multidrug resistance-associated proteins (MRPs) belong to the ATP-binding cassette (ABC) family of membrane transporters and function as efflux pumps, coupling substrate transport to ATP hydrolysis. Of the members of the MRP family, MRP3/ABCC3 and MRP4/ABCC4 are expressed on the basolateral membrane of hepatocytes (Rius et al., 2003; Zelcer et al., 2006) and are considered to mediate the efflux of endo- and xenobiotic compounds and their metabolites from the liver into the blood under normal and cholestatic conditions (Suzuki and Sugiyama, 2000; Borst et al., 2007). In recent studies, Mrp3 and Mrp4 genes were disrupted in mice by homologous recombination (Leggas et al., 2004; Belinsky et al., 2005; Zelcer et al., 2005). Glucuronide conjugates of drugs accumulate in the liver of Abcc3-/- mice after the administration of the parent drug, resulting in lower plasma concentrations of the glucuronide conjugates (Manautou et al., 2005; Zelcer et al., 2005; Zamek-Gliszczynski et al., 2006). Furthermore, the hepatic clearance (presumably, biliary clearance) of morphine-3-glucuronide and the systemic clearance of morphine-6-glucuronide based on the plasma concentration after intraperitoneal administration of respective glucuronides were greater in Abcc3-/- mice (Zelcer et al., 2005). As for the role of Mrp4 in the liver, in situ liver perfusion experiments in Abcc4-/- mice showed that Mrp4 mediates the basolateral efflux of some sulfate conjugates into the blood from the liver (Zamek-Gliszczynski et al., 2006). Moreover, Mrp4 plays a unique and essential protective role in the liver by extruding bile acids into the blood under cholestatic conditions (Mennone et al., 2006).
Methotrexate (N-{4-[(2,4-diaminopteridin-6-ylmethyl)-(methyl)amino]benzoyl}-l-glutamic acid), an inhibitor of dihydrofolate reductase, has been used clinically to treat neoplastic and autoimmune diseases including rheumatoid arthritis and psoriasis. Methotrexate is well absorbed from the small intestine into the systemic circulation (Tracy et al., 1994) followed by excretion into the urine in the unchanged form in humans (Egan et al., 1999). The accumulated evidence thus far suggests that multiple transporters are involved in the disposition of methotrexate. Proton-coupled folate transporter (PCFT)/SLC46A1 (Nakai et al., 2007) and reduced folate carrier (RFC)-1 (Said et al., 1996) are involved in folate uptake from the intestinal lumen via the brush border membrane. For renal elimination of methotrexate, which involves tubular secretion as well as glomerular filtration (Tracy et al., 1994), organic anion transporter (Oat)-3/Slc22a8 mediates uptake into the kidney via the basolateral membrane of the proximal tubules (Nozaki et al., 2004; VanWert and Sweet, 2008), and subsequent luminal efflux in the kidney is mediated by multiple ABC transporters, such as breast cancer resistance protein (Bcrp)/Abcg2 in rodents (Breedveld et al., 2004) and MRP2/ABCC2 in humans (Rau et al., 2006; Nozaki et al., 2007). In rodents, although methotrexate is predominantly excreted in the unchanged form as in humans, biliary excretion makes a greater contribution than urinary excretion (Masuda et al., 1997; Breedveld et al., 2004). In hepatobiliary transport in rodents, the uptake of methotrexate across the basolateral membrane is mediated by a transporter (Ueda et al., 2001), and subsequent excretion into the bile across the canalicular membrane is mainly mediated by Mrp2 in rats (Masuda et al., 1997) and both Bcrp (Breedveld et al., 2004) and Mrp2 (Vlaming et al., 2006) in mice. In addition, both Mrp3 and Mrp4 accept methotrexate as a substrate (Hirohashi et al., 1999; Zeng et al., 2001; van Aubel et al., 2002), although their roles in the disposition of methotrexate remain to be examined.
The purpose of this study was to investigate the role of Mrp3 and Mrp4 in determining systemic exposure of methotrexate. Using Abcc3-/- and Abcc4-/- mice and their corresponding wild-type mice, we carried out in vivo pharmacokinetic studies to examine the roles of Mrp3 and Mrp4 in the biliary excretion and intestinal absorption of methotrexate. To examine the role of Mrp3 in the intestinal absorption in vitro, mucosal-to-serosal transport of methotrexate in everted intestinal sacs was compared between wild-type and Abcc3-/- mice.
Materials and Methods
Chemicals and Reagents. Unlabeled methotrexate was purchased from Dr. B. Schircks (Schircks Laboratories, Jona, Switzerland). [3′,5′,7-3H(N)]Methotrexate (46.8 Ci/mmol) and [24-14C]taurocholate (50 mCi/mmol) were purchased from Moravek Biochemicals (Brea, CA) and American Radiolabeled Chemicals, Inc. (St. Louis, MO), respectively. All other chemicals were commercially available, reagent grade products.
Animals. Male Abcc3-/- (Zelcer et al., 2005) and wild-type FVB mice (10–14 weeks old) and male Abcc4-/- (Leggas et al., 2004) and wild-type C57BL/6 mice (9 weeks old) were used. All animals were kept under standard conditions with a dark-light cycle and were treated humanely. Food and water were available ad libitum. The studies were carried out in accordance with the guidelines provided by the Institutional Animal Care Committee (Graduate School of Pharmaceutical Sciences, The University of Tokyo, Tokyo, Japan).
In Vivo Pharmacokinetic Analyses of Methotrexate in Abcc3-/-and Abcc4-/-Mice. Unanesthetized wild-type, Abcc3-/-, and Abcc4-/- mice were given methotrexate orally (2.2 μmol/kg, 100 mM phosphate buffer, pH 7.0). Blood (approximately 30 μl) was taken from the tail artery at 0.5, 1, 1.5, 2, 3, and 4 h after administration. In another set of studies, under pentobarbital anesthesia, wild-type and Abcc3-/- mice, whose bile duct and bladder were cannulated, received methotrexate by a constant intravenous infusion at the rate of 0.22 μmol/h/kg with a priming dose of 0.22 μmol/kg. Blood (approximately 30 μl) was taken from the jugular vein at 60, 80, 100, and 120 min. Bile and urine were collected every 20 min.
Plasma Protein Binding and Blood-Plasma Partitioning. The free fraction (fp) of methotrexate in mouse plasma (2.2 μM) was determined in vitro by ultrafiltration using a micropartition system (MPS-3; Millipore, Billerica, MA). The free fraction (fp) was given as the ratio of concentrations in ultrafiltrate and plasma. Blood-plasma partitioning (Rb) was determined in freshly collected blood. Methotrexate was added to blank blood to give 2.2 μM concentration in the blood. After incubation for 30 min at 37°C, blood specimens were centrifuged to obtain plasma. Rb was calculated as the concentration ratio in whole blood and plasma.
Kinetic Parameters Related to Hepatobiliary and Urinary Excretion and Bioavailability. Total body clearance (CLtot,p), biliary clearance (CLbile,p), and renal clearance (CLr,p) based on the plasma concentrations and intrinsic biliary clearance based on the liver concentration (CLbile,liver) were calculated from the following equations: 


 where I, Cp,ss, vbile, vurine, and Cliver represent the infusion rate, plasma concentration, the rates of biliary and urinary excretion, and hepatic concentration at steady state, respectively. Kpliver and Kpkidney were defined as the ratio of drug concentration in the organs to the plasma concentration.
where I, Cp,ss, vbile, vurine, and Cliver represent the infusion rate, plasma concentration, the rates of biliary and urinary excretion, and hepatic concentration at steady state, respectively. Kpliver and Kpkidney were defined as the ratio of drug concentration in the organs to the plasma concentration.
Bioavailability (F), hepatic availability (Fh), and the product of fraction absorbed and intestinal availability (FaFg) were calculated from the following equations: 

 where Dosepo represents the dose after oral administration, AUCpo represents the area under the plasma concentration-time curve, CLh,p represents hepatic clearance based on the plasma concentration, and Qh represents hepatic blood flow rate. AUCpo was calculated by the linear trapezoidal rule up to 4 h (AUC0–4) and then extrapolated to infinity using the plasma concentration at 4 h (Cp,4h) and elimination rate constant (ke) as described below.
where Dosepo represents the dose after oral administration, AUCpo represents the area under the plasma concentration-time curve, CLh,p represents hepatic clearance based on the plasma concentration, and Qh represents hepatic blood flow rate. AUCpo was calculated by the linear trapezoidal rule up to 4 h (AUC0–4) and then extrapolated to infinity using the plasma concentration at 4 h (Cp,4h) and elimination rate constant (ke) as described below.  ke was obtained from the slope of the terminal phase of the log-transformed plasma concentration-time curve, where the last 3 points were considered as the terminal phase. Hepatic blood flow rate was cited from previous literature (106 ml/min/kg) (Brown et al., 1997).
ke was obtained from the slope of the terminal phase of the log-transformed plasma concentration-time curve, where the last 3 points were considered as the terminal phase. Hepatic blood flow rate was cited from previous literature (106 ml/min/kg) (Brown et al., 1997).
Hepatic Uptake of Methotrexate in Wild-Type and Abcc3-/- Mice. An integration plot was performed as described previously (Kuwabara et al., 1995; Ueda et al., 2001). In brief, methotrexate (1.1 μmol/kg) was administered intravenously to wild-type and Abcc3-/- mice. Arterial blood and liver specimens were collected within a period of 2 min by sacrificing the mice at different time points (0.5, 1, 1.5, and 2 min). Four animals were used to determine the liver concentration at each time point. Methotrexate concentrations in the specimens were determined as described before. When Kpliver determined at the time of sacrifice (t) was plotted against the area under the plasma concentration-time curve (AUCp,0-t) from time 0 to t divided by the plasma concentration at time t, the slope and y-intercept represent the CLuptake,p and distribution volume at time 0[Vd(0)].  For comparison with the hepatic blood flow rate, the uptake clearance based on plasma concentration (CLuptake,p) was divided by Rb to give the uptake clearance based on the blood concentration (CLuptake,b).
For comparison with the hepatic blood flow rate, the uptake clearance based on plasma concentration (CLuptake,p) was divided by Rb to give the uptake clearance based on the blood concentration (CLuptake,b).
For intraportal administration, a carotid artery of wild-type and Abcc3-/- mice was cannulated for the collection of arterial blood. [3H]Methotrexate and [14C]taurocholate dissolved in mouse plasma were given simultaneously to the mice by an intraportal bolus injection at doses of 50 and 10 μCi/kg, respectively. Arterial blood was collected at 5 to 10 and 10 to 15 s after bolus administration of radiolabeled compounds, and, after 17 s, the portal vein and the hepatic artery were ligated and the liver was excised. The radioactivities associated with plasma and liver specimens were determined in a liquid scintillation analyzer (1900TR; PerkinElmer Life and Analytical Sciences, Waltham, MA).
Mucosal-to-Serosal Transport of Methotrexate in Everted Sacs from Intestine. Mucosal-to-serosal transport of methotrexate in everted sacs from mouse intestines was determined as reported previously (Enokizono et al., 2007). Krebs-Ringer bicarbonate buffer (118 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, 25 mM NaHCO3, and 11 mM d-glucose, pH 6.8) was used in all experiments. The duodenum (1–8 cm from the stomach), jejunum (10–18 cm from the stomach), ileum (8 cm above the cecum), and colon (7 cm below the cecum) of the wild-type and Abcc3-/- mice were excised, trimmed to 6 cm, turned inside out, and immersed into the mucosal buffer (50 ml, 37°C) aerated by 95% O2 + 5% CO2. Serosal buffer was perfused through the everted bowels at 100 μl/min. After 15 min of preincubation, methotrexate was added to the mucosal buffer at a concentration of 22 or 2200 μM, and the total volume of perfused serosal buffer was collected every 5 min. The methotrexate concentration in the perfusate was determined by liquid chromatography/mass spectrometry. The transport rate of methotrexate in the mucosal-to-serosal direction was obtained by multiplying the concentration in the perfusate by the flow rate. The permeability-surface area product for the net methotrexate permeation from the mucosal side to the serosal side (PSnet) was calculated by dividing the transport rate at 40 to 45 min in the mucosal-to-serosal direction by the concentration of methotrexate in the mucosal medium (22 or 2200 μM).
Quantification of mRNA of Transporters Expressed in the Intestine. The mRNA levels of Mrp3, Rfc-1, Pcft, and glyceraldehyde-3-phosphate dehydrogenase (G3pdh) were quantified by real-time polymerase chain reaction (PCR), using the following primers: Mrp3, 5′-GCTCTCACAAGGTGGTACAA-3′ (sense) and 5′-CAGGTTGAAACAGGCACTCA-3′ (antisense); Rfc-1, 5′-GTGGAGTGTCATCTTGGCCCG-3′ (sense) and 5′-CGTTCCAGGAGGAAGGGTGTG-3′ (antisense); Pcft, 5′-GGGAATGGTGGTCTTTGCGT-3′ (sense) and 5′-AGGCCACAGCAGAGAACAGA-3′ (antisense); and G3pdh, 5′-AACGACCCCTTCATTGAC-3′ (sense) and 5′-TCCACGACATACTCAGCAC-3′ (antisense). The mucosa was scraped to obtain an epithelial cell-enriched fraction to prepare total RNA using ISOGEN (Wako Pure Chemicals, Osaka, Japan). Real-time PCR was performed with an SYBR Premix Ex Taq (Takara Bio Inc., Otsu, Japan) and a LightCycler system (Roche Diagnostics, Mannheim, Germany). An external standard curve was generated by dilution of the target PCR product, which was purified by agarose gel electrophoresis. All gene expressions were normalized by the expression of G3pdh.
Western Blotting of Mrp3 in the Kidney. Crude membrane fractions (50 μg of protein) from the kidney and liver of wild-type and Abcc3-/- mice were subjected to electrophoresis and electroblotted to a polyvinylidene difluoride membrane. Then, the polyvinylidene difluoride membrane was incubated with 250 times diluted anti-mouse Mrp3 antiserum (A66) (Zelcer et al., 2006) or 500 times diluted anti-actin antibody (C4; Millipore Bioscience Research Reagents, Temecula, CA) overnight at 4°C. Then, the membrane was incubated with 5000 times diluted horseradish peroxidase-labeled anti-rabbit or anti-mouse IgG (GE Healthcare, Little Chalfont, Buckinghamshire, UK), respectively. An ECL plus system (GE Healthcare) was used for detection, and the intensity of the bands was measured using an image analyzer (LAS-3000 mini; Fuji Film, Tokyo, Japan).
Statistical Analysis. All data represent at least three independent experiments and are expressed as the mean ± S.D. Any statistical significance in the differences of the means was assessed using a two-tailed Student's t test. P < 0.05 was considered statistically significant.
Results
Pharmacokinetic Analyses of the Disposition of Methotrexate in Abcc3-/-and Abcc4-/-Mice. Plasma concentrations of methotrexate after oral administration were significantly lower in Abcc3-/- mice than in wild-type FVB mice, whereas Abcc4-/- mice showed almost the same plasma concentration-time profile as wild-type C57BL/6 mice (Fig. 1). There was some strain difference between wild-type FVB and wild-type C57BL/6 mice in methotrexate pharmacokinetics such as a higher maximal concentration (0.176 ± 0.024 and 0.369 ± 0.054 nmol/ml) and a lower total clearance after oral administration (70.9 ± 16.9 and 46.9 ± 2.3 ml/min/kg) in C57BL/6 mice.
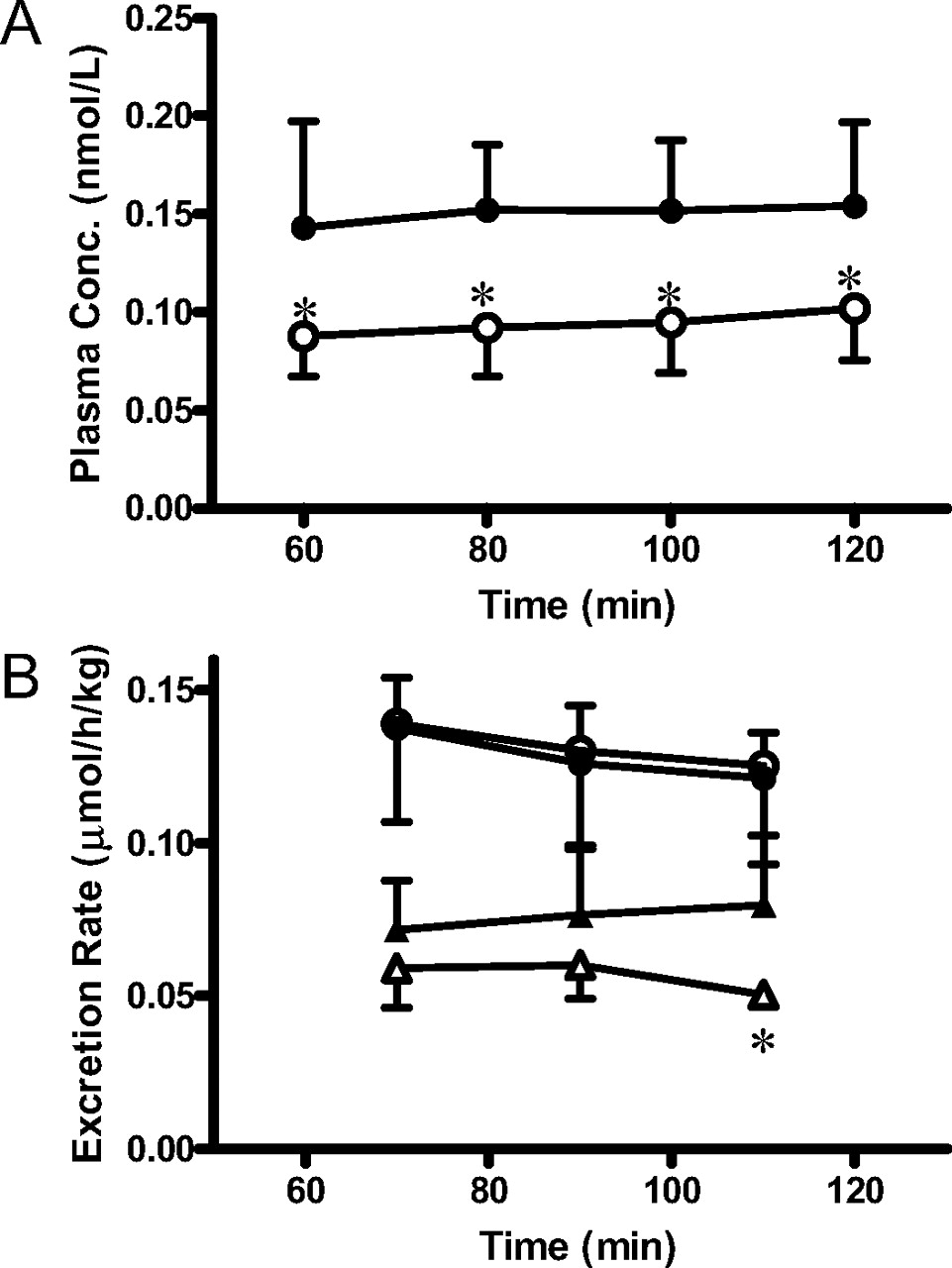
Methotrexate was also given by a constant intravenous infusion to wild-type and Abcc3-/- mice to determine the biliary and urinary excretion rates and tissue concentrations under steady-state conditions (Fig. 2). Pharmacokinetic parameters are listed in Table 1. Hepatic clearance (CLh,p) and biliary clearance based on the plasma concentration (CLbile,p) were significantly greater in Abcc3-/- mice than in wild-type mice. Intrinsic biliary clearances based on the liver concentration (CLbile,liver), representing efflux transport activity at the canalicular membrane, were similar in wild-type and Abcc3-/- mice. Renal clearance (CLr,p) of methotrexate in Abcc3-/- mice was similar to that in wild-type mice, and it was not much different from fpGFR. As an indicator of GFR, the creatinine clearance was determined by liquid chromatography/mass spectrometry to be 13.9 ± 4.1 and 18.3 ± 2.9 ml/min/kg in wild-type and Abcc3-/- mice, respectively, and the difference was not statistically significant. Because these values are similar to the reported GFR (14.0 ml/min/kg) (Davies and Morris, 1993), normal physiological function was maintained. There was no difference in the free fraction in plasma (fp) or blood-plasma partitioning (Rb) between the two strains (Table 1).
Pharmacokinetic parameters of methotrexate in wild-type and Abcc3-/-mice given by intravenous infusion and single oral administration Data were taken from intravenous infusion and single oral dose studies, and ex vivo plasma protein binding and blood to plasma partition assays. The details of pharmacokinetic analyses are described under Materials and Methods. CLtot,p, CLbile,p, CLr,p, and CLbile,liver were calculated from eqs. 1, 2, 3, and 4, respectively. AUCpo was calculated from eq. 8, and F, Fh, and FaFg were calculated from eqs. 5, 6, and 7, respectively. Each value represents the mean ± S.D. of at least four independent experiments.
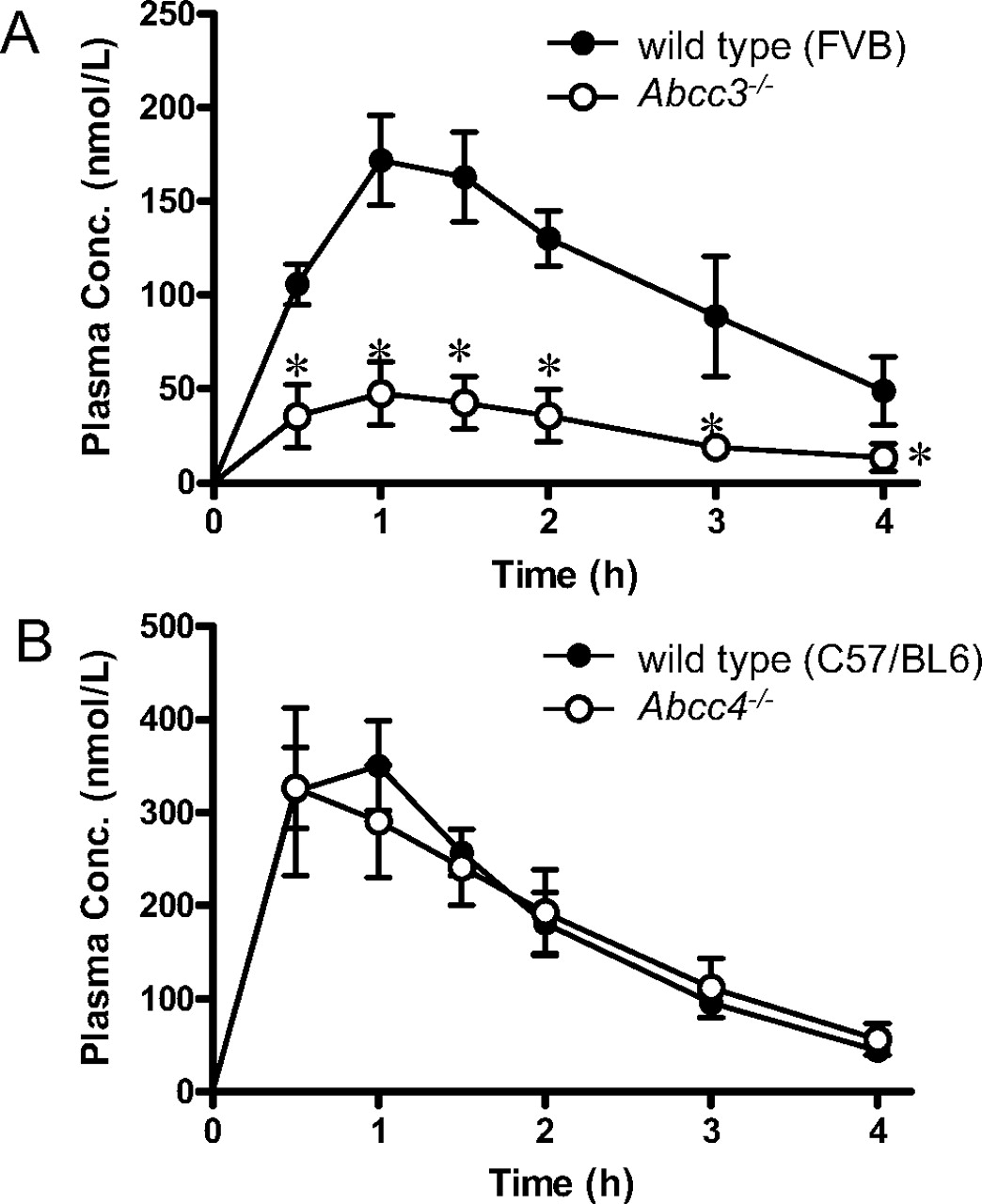
Time profiles of the plasma concentrations of methotrexate after oral administration in wild-type, Abcc3-/-, and Abcc4-/- mice. A, methotrexate (2.2 μmol/kg) was given orally to wild-type FVB (•) and Abcc3-/- (○) mice. The plasma concentrations were determined at the designated times. Each point represents the mean ± S.D. (n = 4 or 5). *, P < 0.05, significantly different from wild-type mice. B, methotrexate (2.2 μmol/kg) was given orally to wild-type C57BL/6 (•) and Abcc4-/- (○) mice. The plasma concentrations were determined at the designated times. Each point represents the mean ± S.D. (n = 5).
Bioavailability (F) was calculated from AUCpo and CLtot,p (Table 1). Because bioavailability is a product of the fraction of absorption (Fa) and availability in the small intestine and liver (Fg and Fh, respectively), dividing F by Fh, calculated from the hepatic clearance (CLh,p) and the hepatic blood flow rate (106 ml/min/kg) (Brown et al., 1997), assuming that nonrenal clearance is hepatic clearance and there is no excretion via the gut, gave the product of fraction absorbed and intestinal availability (FaFg). Both Fh and FaFg were smaller in Abcc3-/- mice than in wild-type mice without statistical significance (Table 1).
Integration Plot Analysis of the Hepatic Uptake of Methotrexate. The hepatic uptake clearance was determined by integration plot analysis in wild-type and Abcc3-/- mice (Fig. 3). As shown in eq. 9, the slopes of the integration plots represent the hepatic uptake clearances based on plasma concentration (CLuptake,p). CLuptake,p values were 0.785 ± 0.026 and 0.923 ± 0.014 ml/min per gram of liver in wild-type mice and Abcc3-/- mice, respectively. The hepatic uptake clearances based on the blood concentration (CLuptake,b), obtained by dividing CLuptake,p by Rb, were 1.38 and 1.51 ml/min per gram of liver, respectively, which are approximately three quarters of the previously reported hepatic blood flow rate (1.94 ml/min per gram of liver) (Brown et al., 1997).

Time profiles of the plasma concentrations of methotrexate after intravenous administration, and biliary and urinary excretion rates of methotrexate in wild-type and Abcc3-/- mice. A, methotrexate was given to wild-type FVB (•) and Abcc3-/- (○) mice, whose bile duct and bladder had been cannulated for collection of bile and urine, by a constant intravenous infusion at the rate of 0.22 μmol/h/kg with a priming dose of 0.22 μmol/kg. The plasma concentrations were determined at the designated times. B, the excretion of methotrexate into the bile (circles) and urine (triangles) was determined at 20-min intervals in wild-type (closed symbols) and Abcc3-/- (open symbols) mice. Each point represents the mean ± S.D. (wild type: n = 6; Abcc3-/-: n = 4 or 5). *, P < 0.05, significantly different from wild-type mice.
Hepatic Extraction of Methotrexate after Intraportal Bolus Injection. Wild-type and Abcc3-/- mice (n = 3) were given methotrexate by an intraportal bolus administration to determine the hepatic uptake during a single pass. Taurocholate was simultaneously administered as an indicator of extremely efficient hepatic uptake. There was no significant change in the ratio of methotrexate in the liver relative to the dose injected, 58.3 ± 6.6 and 56.9 ± 9.3% in wild-type and Abcc3-/- mice, respectively. The corresponding values for taurocholate were 110 ± 3 and 104 ± 4% in wild-type and Abcc3-/- mice, respectively. There was no difference in arterial blood concentration between the two strains.
Mucosal-to-Serosal Transport of Methotrexate in Everted Sacs from Intestine. The mucosal-to-serosal transport of methotrexate in the intestine was determined in vitro using everted sacs prepared from the duodenum, jejunum, ileum, and colon of wild-type and Abcc3-/- mice (Fig. 4; Table 2). The permeability-surface area product of the net methotrexate transport from the mucosal side to the serosal side (PSnet) was saturable in the duodenum (Table 2). The PSnet in other intestinal segments at a low methotrexate concentration (22 μM) was similar or smaller than the PSnet in the duodenum at a high methotrexate concentration (2200 μM). PSnet was significantly lower in the duodenal everted sacs of Abcc3-/- mice compared with wild-type mice at a low methotrexate concentration (22 μM), and the methotrexate concentration in the tissue was 1.9-fold higher in Abcc3-/- mice than in wild-type (3.04 ± 1.21 and 5.90 ± 1.46 μMin wild-type and Abcc3-/- mice, respectively, P < 0.05). At a high methotrexate concentration (2200 μM), there was no significant difference in PSnet between wild-type and Abcc3-/- mice (Table 2). Intrinsic serosal efflux clearance, obtained by dividing the mucosal-to-serosal transport rate by tissue concentrations at a low methotrexate concentration, represents the efflux transport activity in the basolateral membrane of the enterocytes. The intrinsic serosal efflux clearance of methotrexate was significantly lower in the duodenum of Abcc3-/- mice compared with wild-type mice (22.4 ± 5.4 and 5.26 ± 1.29 μl/min/sac in wild-type and Abcc3-/- mice, respectively, P < 0.05, at 22 μM in the mucosal buffer). In other segments, there were no significant differences in PSnet between wild-type and Abcc3-/- mice.
Transport rate and permeability-surface area product of methotrexate across the intestinal everted sac prepared from wild-type and Abcc3–/– mice Data were taken from Fig. 4. The transport rate was obtained by multiplying the concentration in the perfusate at 40 to 45 min by the flow rate. The permeability-surface area product for the net methotrexate permeation from the mucosal side to the serosal side (PSnet) was calculated by dividing the transport rate at 40 to 45 min by the concentration in the mucosal medium. Each value represents the mean ± S.D. (duodenum: n = 6 or 7; jejunum: n = 7; ileum: n = 4; colon: n = 3 or 4).
Distribution of Folate Transporter mRNA in the Intestine. Intestinal mRNA expression levels of Mrp3, Pcft, and Rfc-1 were quantified in wild-type and Abcc3-/- mice (Fig. 5). Pcft and Rfc-1 are suggested to be involved in the intestinal uptake of methotrexate. The expression of Mrp3 was highest in the colon, and expression was almost equal in other segments. Pcft was predominantly expressed in the duodenum, and Rfc-1 was most highly expressed in duodenum. Expression levels of these transporters were much lower in other intestinal segments. Pcft expression was slightly higher in the duodenum and ileum of Abcc3-/- mice compared with wild-type mice, but there was no significant difference in Rfc-1 expression between strains.
Expression of Mrp3 Protein in the Kidney. In the kidney, the expression of Mrp3 protein was detected by Western blotting (Fig. 6). The band associated with the Mrp3 protein was detected in the membrane fraction from wild-type mice, and it was not detected in membranes from Abcc3-/- mice. Expression levels in the kidney were much lower than those in the liver.
Discussion
In this study, we investigated the roles of Mrp3 and Mrp4 in the disposition of methotrexate using Abcc3-/- and Abcc4-/- mice. It was found that the plasma concentration of methotrexate after oral administration was lower in Abcc3-/- mice compared with that in wild-type mice (Fig. 1A). This suggests a greater systemic clearance and/or lower oral absorption in Abcc3-/- mice. First, the role of Mrp3 in the hepatobiliary and urinary excretion of methotrexate was examined by comparing its pharmacokinetic parameters between wild-type and Abcc3-/- mice receiving a constant intravenous infusion of methotrexate. Although the plasma concentrations were significantly lower in Abcc3-/- mice compared with wild-type mice (Fig. 2), the biliary excretion rate was similar in wild-type and Abcc3-/- mice. That is, the biliary clearance based on the plasma concentration (CLbile,p) was greater in Abcc3-/- mice than in wild-type mice, whereas the renal clearance (CLr,p) was similar (Table 1). The biliary clearance based on the plasma concentration (CLbile,p) consists of hepatic blood flow rate (Qh), unbound fraction in plasma (fp), blood-plasma partitioning (Rb), and three intrinsic parameters, representing basolateral uptake (CLinf), basolateral efflux (CLeff), and canalicular efflux (CLbile,liver). 
Because there were no significant differences in the unbound fraction, blood-plasma partitioning, basolateral uptake, and canalicular efflux between wild-type and Abcc3-/- mice (Table 1), it is most likely that the basolateral efflux clearance of methotrexate is reduced in Abcc3-/- mice to account for the increase in the biliary clearance. This is also supported by the greater liver-to-plasma concentration ratio observed in Abcc3-/- mice, indicating that total efflux from the liver was decreased in Abcc3-/- mice. In Abcc3-/- mice, the expression of Mrp4 protein, another basolateral efflux transporter for methotrexate in the liver, was similar to that of wild-type mice (Matsushima et al., 2008), and the decrease in the basolateral efflux of methotrexate in the liver was probably ascribed to the lack of Mrp3-mediated efflux.
It is known that methotrexate is metabolized to 7-hydroxymethotrexate. In this study, the pharmacokinetic disposition of 7-hydroxymethotrexate was not determined, and the clearance obtained by subtracting the biliary clearances from the nonrenal clearance was regarded as the metabolic clearance. The biliary clearance based on the plasma concentration is similar to the nonrenal clearance in wild-type mice, indicating that metabolic clearance makes only a minor contribution, 10% at most, if any, to the hepatic clearance in wild-type mice. In Abcc3-/- mice, the metabolic clearance of methotrexate based on the plasma concentration was 3.4-fold greater than the corresponding value in wild-type mice, although it still makes minor contribution to the nonrenal clearance, 20% at most. This increase is partly explained by a reduction in basolateral efflux in the liver due to the loss of Mrp3, as observed for biliary excretion. The possibility of adaptive regulation of cytochrome P-450 enzymes due to the loss of Mrp3 cannot be excluded in the present study. Caution must be paid to interpreting pharmacokinetic data in Abcc3-/- mice for drugs that are mainly eliminated from the liver by metabolism.

Comparison of the hepatic uptake clearance between wild-type and Abcc3-/- mice. Arterial plasma (A) and liver (B) concentrations of methotrexate in wild-type (closed circles) and Abcc3-/- mice (open circles) were determined at designated times after a single bolus intravenous administration of methotrexate (1.1 μmol/kg). Kpliver at time t were plotted against AUCp,0-t divided by the plasma concentration at time t (C). The slope of the plot represents CLuptake,p. Solid lines represent the linear regression line for wild-type and Abcc3-/- mice. Each plot represents the mean ± S.D. (n = 4).
Unlike Abcc3-/- mice, Abcc4-/- mice did not show any change in the plasma concentration-time profile after oral administration (Fig. 1B). Absence of an effect of functional impairment of Mrp4 may suggest its limited contribution to the disposition of methotrexate. However, we previously reported minimal change in the systemic elimination of Mrp4 substrates, such as ceftizoxime and cefazolin, but a significant increase in the kidney accumulation in Abcc4-/- mice (Ci et al., 2007). Functional impairment of Mrp4 may affect tissue concentrations, and further studies are required to reach a concrete conclusion about the role of Mrp4 in the disposition of methotrexate.
Greater hepatic clearance based on plasma concentration accounts partly for the lower bioavailability of methotrexate in Abcc3-/- mice compared with wild-type mice (Table 1). Furthermore, the lower FaFg in Abcc3-/- mice (Table 1) suggests that the intestinal absorption of methotrexate is decreased in Abcc3-/- mice. To confirm this possibility, intestinal transport was examined in vitro using everted sacs from the intestine. PSnet was saturable in the duodenal everted sacs, and PSnet in other segments was similar to PSnet in the duodenum at a high methotrexate concentration (Table 2). Therefore, a saturable mechanism accounts for the large mucosal-to-serosal transport of methotrexate in the duodenum at a low concentration. PSnet at a low methotrexate concentration (22 μM) was significantly decreased in everted sacs from the duodenum of Abcc3-/- mice compared with wild-type mice, whereas the corresponding parameter determined at a high concentration was similar (Table 2). The fact that tissue concentration of methotrexate was significantly greater in the duodenal everted sacs of Abcc3-/- mice compared with wild-type mice suggests that the lower PSnet in Abcc3-/- mice is not due to the reduction in uptake ability but to the reduction in efflux from inside of the enterocytes to the serosal side. Indeed, the intrinsic serosal efflux of methotrexate was significantly decreased in the duodenum of Abcc3-/- mice compared with that in wild-type mice. Because Mrp3 is expressed in the basolateral membrane of enterocytes (Rost et al., 2002; Shoji et al., 2004; Zelcer et al., 2006), these data suggest that Mrp3 mediates the basolateral efflux of methotrexate in the duodenum. Unlike the duodenum, there were no significant differences in the PSnet of methotrexate in other segments. It should be noted that Mrp3 protein is expressed in whole segments of the intestine (Zelcer et al., 2006). Therefore, a discrepancy exists between the distribution of Mrp3 along the intestine and the effect of the loss of Mrp3 on the PSnet of methotrexate. There are two possibilities for this result. First, the basolateral efflux is mediated by other transporters in the mid to distal segments. Second, uptake transporters for methotrexate exhibit heterogeneous distribution along the intestine because uptake transporters are involved in the mucosal-to-serosal transport of methotrexate in conjunction with Mrp3. mRNA quantification by real-time PCR showed abundant expression of the mRNA of uptake transporters of methotrexate such as Pcft and Rfc-1 in the duodenum (Fig. 5). This supports the second possibility, whereas the first possibility remains to be examined. Considering that MRP3 exhibits broad substrate specificity to xenobiotic compounds (Suzuki and Sugiyama, 2000; Borst et al., 2007), it is possible that MRP3 also plays a significant role in the disposition of other drugs.

Mucosal-to-serosal transport of methotrexate in the everted sac from the intestine. Everted segments of the duodenum, jejunum, ileum, and colon of wild-type and Abcc3-/- mice were incubated with methotrexate, and serosal side was perfused with drug-free buffer. The total volume of serosal perfusate was collected every 5 min. Methotrexate concentrations in the serosal perfusate of the everted sacs from duodenum (A and B), jejunum (C), ileum (D), and colon (E) of wild-type (closed circles) and Abcc3-/- (open circles) mice were determined. The mucosal concentration of methotrexate was 22 (A and C–E) or 2200 μM (B). Each point represents the mean ± S.D. (duodenum: n = 6 or 7; jejunum: n = 7; ileum: n = 4; colon: n = 3 or 4). *, P < 0.05, significantly different from wild-type mice.
It has also been shown that the pharmacokinetic characteristics of methotrexate and its hepatobiliary transport exhibit great interspecies differences, even between mice and rats. CLuptake,p in mice is 3.6-fold greater than that in rats (43.1 and 11.9 ml/min/kg in mice and rats, respectively), although the CLbile,p exhibits only a 1.6-fold difference between the two species (14.9 and 9.43 ml/min/kg, respectively). This is ascribed to the difference in the rate-limiting process. CLuptake,p was 3-fold greater than CLbile,p in mice, whereas CLuptake,p and CLbile,p were similar in rats (Ueda et al., 2001). That is, uptake is the rate-limiting process in overall hepatobiliary excretion in rats but not in mice. When the canalicular efflux clearance (CLbile,liver) is greater than the basolateral efflux clearance (CLeff), the uptake process becomes rate limiting for overall elimination. Therefore, the relationship between the canalicular efflux clearance (CLbile,liver) and basolateral efflux clearance (CLeff) should show an interspecies difference between rats and mice. Indeed, the intrinsic biliary clearance based on the liver concentration (CLbile,liver) was almost 8-fold smaller in mice (1.65 ml/min/kg) than the corresponding value in rats (13.1 ml/min/kg). Although the basolateral efflux clearance was not directly determined in both rats and mice, it is possible that the basolateral efflux clearance (CLeff) is smaller in rats than in mice because Mrp3 is barely expressed in rat liver (Hirohashi et al., 1998). In humans, unlike rodents, the recovery of methotrexate in the bile using the duodenal aspiration technique is less than 10% of the dose after intravenous administration (Shen and Azarnoff, 1978), although human hepatic transporters such as OATP1B1 (Abe et al., 2001), MRP2 (Hooijberg et al., 1999), and BCRP (Chen et al., 2003) accept methotrexate as a substrate.
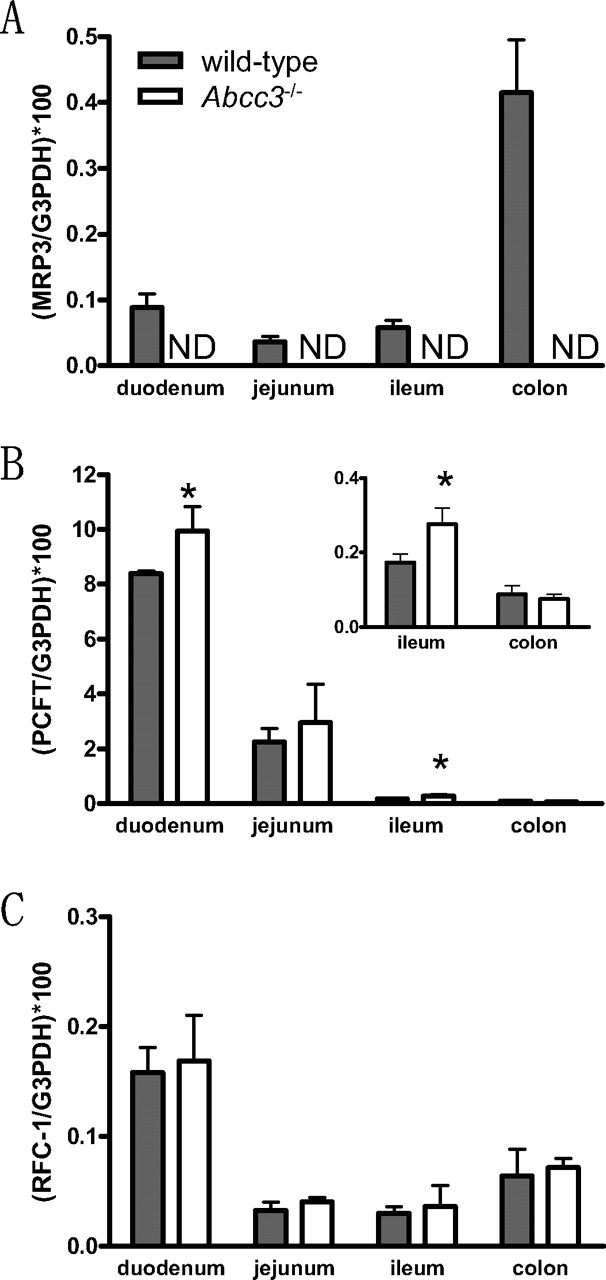
mRNA expression of Mrp3, Pcft, and Rfc-1 along the small intestine, and colon from wild-type and Abcc3-/- mice. mRNA expression of Mrp3 (A), Pcft (B), and Rfc-1 (C) were quantified by real-time PCR, and they were corrected by those of G3pdh. Each column represents the mean ± S.D. (n = 3). *, P < 0.05, significantly different from wild-type mice. ND, not detected.

Mrp3 protein expression in mouse kidney and liver by Western blot analysis. The crude membrane fractions (50 μg of protein) of the kidney and liver from wild-type (WT) and Abcc3-/- mice were subjected to Western blot analysis. Mrp3 protein was detected in the kidney and liver of wild-type mice, but no corresponding band was detected in those of Abcc3-/- mice.
For renal transport, the renal clearance of methotrexate was similar in wild-type and Abcc3-/- mice; however, the kidney-to-plasma concentration ratio (Kpkidney) was significantly increased in Abcc3-/- mice. Although Western blot analysis detected expression of Mrp3 protein in the kidney (Fig. 6), the exact membrane localization of Mrp3 in mouse kidney remains unclear and is needed to understand the role of Mrp3 in the kidney. Because Mrp3 is localized to the renal tubules in rat kidney (Kuroda et al., 2004), and to the basolateral membrane of distal convoluted tubules, but not to the proximal tubules in human kidney (Scheffer et al., 2002), it is possible that Mrp3 is involved in the basolateral efflux of methotrexate in the mouse kidney.
In conclusion, this study showed that basolateral efflux in the liver mediated by Mrp3 increases the exposure of methotrexate in the systemic circulation, and facilitates absorption in the duodenum in conjunction with uptake transporters, whereas Mrp4 probably plays only a limited role in systemic exposure.
Acknowledgments
We thank Dr. Piet Borst (The Netherlands Cancer Institute, Amsterdam, The Netherlands) for supplying Abcc3-/- mice and anti-mouse Mrp3 polyclonal antiserum (A66) and Drs. Koen van de Wetering (The Netherlands Cancer Institute, Amsterdam, The Netherlands) and Noam Zelcer (Department of Pathology and Lab-oratory Medicine, University of California, Los Angeles, CA) for fruitful suggestions.
Footnotes
-
This study was supported by a grant-in-aid for Scientific Research (A) from Japan Society for the Promotion of Science (JSPS) (KAKENHI 17209005; to Y.S.) and a grant-in-aid for Scientific Research (B) from JSPS (KAKENHI 18390046; to H.K.).
-
Y.K. and M.H. contributed equally to this study.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.108.140475.
-
ABBREVIATIONS: MRP/Mrp, multidrug resistance-associated protein; ABC/Abc, ATP-binding cassette; PCFT/Pcft, proton-coupled folate transporter; RFC/Rfc, reduced folate carrier; OAT/Oat, organic anion transporter; BCRP/Bcrp, breast cancer resistance protein; G3pdh, glyceraldehyde-3-phosphate dehydrogenase; PCR, polymerase chain reaction; OATP, organic anion transporting polypeptide.
- Received April 28, 2008.
- Accepted July 14, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}