


Abstract
Hemoglobin vesicles (HbVs) could serve as a substitute for red blood cells (RBCs) in resuscitation from massive hemorrhage. A massive transfusion of RBCs can increase the risk of infection, which is not caused by contaminating micro-organisms in the transfused RBCs but by a breakdown of the host defense system. We previously found that complement activity was increased after resuscitation with HbVs at a putative dose in a rat model of hemorrhagic shock. It is known that complement system plays a key role in host defense in the embryonic stage. Therefore, the objective of this study was to address whether the suppression of bacterial infections in hemorrhagic shock rats was a result of increased complement activity after massive HbV transfusion. For this purpose, Escherichia coli were incubated with plasma samples obtained from a rat model of hemorrhagic shock resuscitated by HbVs or RBCs, and bacterial growth was determined under ex vivo conditions. As a result, E. coli growth was found to be suppressed by increased complement activity, mediated by the production of IgM from spleen. However, this antibacterial activity disappeared when the E. coli were treated with complement-inactivated plasma obtained from splenoctomized rats. In addition, the resuscitation of HbVs from hemorrhagic shock increased the survival rate and viable bacterial counts in blood in cecum ligation and puncture rats, a sepsis model. In conclusion, the resuscitation of HbVs in the rat model of hemorrhagic shock suppresses bacterial growth via complement activation induced by IgM.
Introduction
Hemoglobin vesicles (HbVs), which were developed for use as an artificial oxygen carrier, have a cellular structure similar to that of red blood cells (RBCs), in that they are highly concentrated Hbs encapsulated in a phospholipid bilayer membrane with polyethylene glycol (Sakai et al., 2009b). HbV has been shown to possess a number of positive characteristics: the absence of viral contamination (Sakai et al., 1993), a long-term storage period of more than 2 years at room temperature (Sakai et al., 2000b; Sou et al., 2000), low toxicity (blood compatibility, no nephrotoxicity) (Sakai et al., 2000a), and good metabolic performance (Taguchi et al., 2009b, 2010). Moreover, the pharmacological effects of HbVs have been reported to be equivalent to that of RBCs, when evaluated in a model of hemorrhagic shock in rats (Sakai et al., 2004b, 2009a). In addition, the retention of HbVs in the circulation compares favorably with other artificial oxygen carriers (Taguchi et al., 2009b), and the half-life in humans is predicted to be sufficiently long to allow autologous blood to be recovered after a massive hemorrhage (Taguchi et al., 2009a). Based on these facts, it would be predicted that HbVs would be superior to a conventional blood transfusion and have considerable promise for use as a RBC alternative in patients with massive hemorrhages.
In modern medical care, there is now little doubt that the transfusion of RBCs is the gold standard for the treatment of patients with massive hemorrhages. However, such massive infusions are associated with numerous and significant complications, which include acute hemolytic transfusion reactions, bacterial sepsis, microchimerism, and transfusion-related acute lung injuries. Among these drawbacks, bacterial infections, which are not initially present as contaminants in transfusion solutions but invade afterward, remain a serious problem in the cases of injured and postoperative patients. In fact, it was reported that an increased risk of infection was as high as 15 to 40% in critically ill patients after massive transfusions (Hill et al., 2003; Rachoin et al., 2009). Because the increased incidence of bacterial infections causes increased morbidity and mortality, total hospital costs, and the total length of hospital stay, decreased incidences of bacterial infections in massive hemorrhagic patients would contribute, not only to a reduction in medical costs, but also to the enhancement of the quality of life of the affected patients.
One of the possible mechanisms for the increased rate of infection is the loss of immune system components/cells caused by hemorrhage. In addition, it is known that the increased rate of infection after massive hemorrhage is related to both the immunosuppression and breakdown of the gut barrier. The mechanism for the gut barrier breakdown is induced by ischemia reperfusion of small intestine. The precise mechanisms for immunosuppression remain uncertain, but increased bacterial infection rate is a known consequence of the breakdown of the host defense system (Sihler and Napolitano, 2010). The host defense system is comprised of two major effector systems that act immediately: 1) the complement system and the cellular system, which is composed of macrophages and neutrophils and 2) the adaptive immune system comprised of antigen-specific T and B lymphocytes. The complement represents the first line of defense for the host defense system and is the first contact with an infectious agent during the adaptive antigen-specific response. Therefore, the complement system is an important factor in the embryonic stage of development. In previous studies, we reported that IgM was induced by the resuscitation of HbVs at a putative dose, and the highest value was found at 4 days. It is well known that IgM is related to complement activation via the classic pathway. Therefore, it is possible that a massive transfusion of HbVs could result in the enhancement of the host defense system, as the result of an increase in complement activity, which would then suppress bacterial infections.
In this study, we hypothesized that the administration of HbVs under conditions of a massive hemorrhage would suppress the bacterial growth resulting from an increase in complement activity. For this purpose, we developed a rat model of hemorrhagic shock, and it was then resuscitated by HbVs or washed RBCs (wRBCs) at a putative dose of 1400 mg Hb/kg. At 4 and 7 days after resuscitation by HbVs (HbV4day, HbV7day) and wRBCs (RBC4day, RBC7day), we then evaluated the effects of complement titer (CH50) and plasma IgM on bacterial growth in ex vivo conditions. In addition, to confirm the effects of antibacterial activity under in vivo conditions, we created a double-hit model of hemorrhagic shock by cecum ligation and a puncture rat model, which is arguably a situation that closely replicates the nature and course of clinical sepsis in patients after trauma, and investigated the survival and viable bacterial counts in blood samples.
Materials and Methods
Preparation of HbV and wRBC Solution.
HbVs were prepared under sterile conditions as reported previously (Sakai et al., 1997). The Hb solution was purified from outdated donated blood, which was provided by the Japanese Red Cross Society (Tokyo, Japan). The encapsulated Hb (38 g/dl) contained 14.7 mM pyridoxal 5′-phosphate (Sigma-Aldrich, St. Louis, MO) as an allosteric effector to regulate the P50 to 25 to 28 Torr. The lipid bilayer was a mixture of 1,2-dipalmitoyl-sn-glycero-3-phosphatidylcholine, cholesterol, and 1,5-bis-O-hexadecyl-N-succinyl-l-glutamate (Nippon Fine Chemical Co. Ltd., Osaka, Japan) at a molar ratio of 5/5/1 and 1,2-distearoyl-sn-glycero-3-phosphatidyl-ethanolamine-N-polyethylene glycol (NOF Corp., Tokyo, Japan) (0.3 mol%). The size of the HbVs was controlled at approximately 250 nm using an extrusion method. The HbVs were suspended in physiological salt solution at 10 g/dl [Hb], filter-sterilized (Dismic; Toyo-Roshi, Tokyo, Japan; pore size, 450 nm), and bubbled with N2 for storage.
To prepare wRBCs, blood samples from donated Sprague-Dawley (SD) rats (Kyudou Co., Kumamoto, Japan) were withdrawn and centrifuged at 1200g for 15 min to obtain an RBC concentrate. This sample was then washed three times to remove plasma components. The Hb concentration was determined by the cyanometHb method using a Hemoglobin B-Test-Wako kit (Wako Pure Chemical, Osaka, Japan). The wRBC samples were suspended in a physiological salt solution at 10 g/dl [Hb].
Before all of the experiments, HbVs or wRBCs were mixed with recombinant human serum albumin (Nipro Corp., Osaka, Japan) to adjust the albumin concentration of the suspension medium to 5 g/dl. Under these conditions, the colloid osmotic pressure of the suspension is maintained constant at approximately 20 mm Hg (Sakai et al., 2004b).
Preparation of Hemorrhagic Shock Model Rats.
All animal experiments were performed according to the guidelines, principles, and procedures for the care and use of laboratory animals of Kumamoto University. SD rats were maintained in a temperature-controlled room with a 12-h dark/light cycle and ad libitum access to food and water. Hemorrhagic shock model rats were prepared as described previously (Taguchi et al., 2009a). In brief, SD rats (180–210 g) were anesthetized with pentobarbital. Polyethylene catheters (PE 50 tubing, o.d. equal to 0.965 mm, and i.d. equal to 0.58 mm; Becton Dickinson and Co., Tokyo, Japan) containing saline and heparin were then introduced into the left femoral artery for infusion and blood withdrawal. Hemorrhagic shock was induced by removing 40% of the total blood volume (22.4 ml/kg). Systemic blood volume was estimated to be 56 ml/kg (Sakai et al., 2004b). After removing the blood, the hemorrhagic shock rats were resuscitated by an infusion of either isovolemic HbVs or wRBCs (1400 mg Hb/kg, 22.4 ml/kg). After resuscitation, the polyethylene catheter was removed, the femoral artery was ligated, and the skin was sutured with a stitch. The animals were housed in a temperature-controlled room with a 12-h dark/light cycle and ad libitum access to food and water. The arterial blood oxygen tension (PaO2) and pH were evaluated before hemorrhage with the left femoral artery and 4 and 7 days after resuscitation with the right femoral artery. The i-STAT system (Abbott Laboratories (Abbott Park, IL) was used for analyses of PaO2 and pH.
Sample Collection.
At 4 or 7 days after resuscitation from hemorrhagic shock by HbV and wRBC solution, blood samples were collected from tail vein, and plasma samples were obtained by centrifugation (3000g, 5 min). Antibacterial activity was immediately measured, and the remaining samples were stored at −80°C until use in measurements of CH50 and plasma IgG and IgM.
The Evaluation of Antibacterial Activity.
In vitro antibacterial activities were determined by previously reported methods with minor modification (Ishima et al., 2007). M9 medium, pH 7.4, was used during the incubation of bacteria with serum. In a typical run, Escherichia coli ATCC strains were cultured overnight in M9 medium. A 50-μl bacterial suspension in M9 medium (OD630; 0.05 ± 0.01) was placed in a 96-well plate, and 50-μl aliquots of plasma samples were added. After 6 h of incubation at 37°C, the number of bacteria exposed to various plasma samples were determined by measuring the absorbance 630 nm using a microplate reader (model 450; Bio-Rad Laboratories, Hercules, CA).
Surgery for Splenectomy.
For splenectomy, the rats were anesthetized using pentobarbital and, after shaving, an incision was made in the skin at the left flank. The peritoneal membrane was opened, and the entire spleen was removed intact after ligating the splenic vein and artery at the hilum. The peritoneal membrane and skin were separately sutured (Ishida et al., 2006). One week after surgery for splenectomy, the rats were used in experiments.
Measurement of Plasma IgG, IgM, and CH50.
The plasma samples obtained 4 and 7 days after resuscitation by the administration of HbV and wRBC solutions were stored at −80°C before analysis by a commercial clinical testing laboratory (SRL, Tokyo, Japan). CH50 was measured in hemolytic assays as described previously (Mayer, 1961).
Measurement of Phagocyte Activity.
Phagocyte activity was determined by the carbon clearance method, as described previously (Taguchi et al., 2011b). Hemorrhagic shock was induced in the rats, which were then resuscitated with HbVs and wRBCs. In a typical experiment, rats were anesthetized with pentobarbital. Polyethylene catheters (PE 50 tubing) containing saline and heparin were then introduced into the left femoral vein for the infusion of a carbon particle solution and blood collection. The carbon particle solution (Fount India Ink; Pelikan Co., Hannover, Germany) was infused at 10 ml/kg within 1 min. At 4, 10, 20, 30, 45, and 60 min later, approximately 100 μl of blood was withdrawn, and exactly a 50-μl aliquot was diluted with 5 ml of a 0.1% sodium bicarbonate solution. The absorption was measured at 675 nm by means of a spectrophotometer (U-2900; Hitachi, Tokyo, Japan). The phagocyte index (K) was calculated using the equation: K = 1/(t2 − t1) × ln(C1/C2), where C1 and C2 are the concentrations (absorbance) at time t1 and t2 (min), respectively.
Preparation of Cecum Ligation and Puncture Model and the Measurement of Viable Bacterial Counts in Blood.
Bacterial infection was induced by cecum ligation and puncture, following a previously reported method with minor modification (Hubbard et al., 2005). Hemorrhagic shock was induced in the rats, which were then resuscitated with HbVs and wRBCs. Four or 7 days after resuscitation with HbVs and wRBCs, the cecum was ligated without preventing passage of gastric contents. Ten punctures in cecum ligation were then performed using a 21-gauge needle. Survival rate was determined daily for 7 days after puncture. The blood samples from surviving rats (0.3 ml) were collected from tail vein 24 h after the puncture. After collecting blood samples, the aliquot blood (0.1 ml) was immediately placed on Luria-Bertani agar plates (9.9 ml) and the plates were incubated at 37°C for 24 h. The numbers of bacterial colonies were then counted and expressed as colony-forming units/ml.
Data Analysis.
Data are shown as the means ± S.D. for the indicated number of animals. Significant differences among each group were determined using the two-tail unpaired Student's t test. The Spearman test was used for correlation analyses. The survival rate was compared using Kaplan-Meier survival curves and the log-rank test. A probability value of p < 0.05 was considered to indicate statistical significance.
Results
Survival Rate and Blood Gasses Analysis.
All hemorrhagic shock rats resuscitated by an infusion of either isovolemic HbVs or wRBCs (1400 mg Hb/kg, 22.4 ml/kg) survived during experiments and were well tolerated with no apparent change in behavior. The PaO2 before induction of hemorrhagic shock by bleeding was 88.8 ± 5.7 Torr and increased significantly to 129.3 ± 17.4 Torr after induction of hemorrhagic shock. After resuscitation with HbVs or wRBCs, PaO2 recovered similar to PaO2 measured at baseline (76.3 ± 7.8 and 93.5 ± 16.5 Torr for 4 and 7 days after HbV resuscitation, respectively; 86.8 ± 14.8 and 87.8 ± 11.1 Torr for 4 and 7 days after wRBC resuscitation, respectively). In addition, pH before and after hemorrhage was 7.33 ± 0.07 and 7.28 ± 0.08, respectively, and it recovered by resuscitation with HbVs (7.37 ± 0.06 and 7.43 ± 0.05 for 4 and 7 days after resuscitation, respectively) or wRBCs (7.41 ± 0.03 and 7.44 ± 0.02 for 4 and 7 days after resuscitation, respectively). The data showed similar tendency as that reported previously by Sakai et al. (2009a).
The Evaluation of Antibacterial Activity.
To evaluate antibacterial activity, the collected plasma samples were incubated with E. coli (ATCC strain) for 6 h. As shown in Fig. 1A, the only plasma sample in HbV4day showed a dramatic level of antibacterial activity, whereas other plasma samples obtained from the hemorrhagic shock rats did not show any activity. Because it is possible that HbV directly suppresses E. coli growth, HbV alone was incubated with E. coli (0, 0.049, 0.098, 0.195, 0.391, and 0.781 mg Hb/ml). As a result, HbVs did not show any antibacterial activity during 6 h (data not shown). This result clearly excluded the possibility of HbV itself directly contributing to the antibacterial activity observed in plasma samples from the hemorrhagic shock rats that had been resuscitated by HbV. Consequently, other factors in plasma induced by resuscitation with HbV seem to be indirectly involved in this unique phenomenon.

A, bacterial (E. coli ATCC strains) growth rate after resuscitation with HbVs or wRBCs in a rat model of hemorrhagic shock. Blood was collected from the tail vein, and plasma was obtained. E. coli ATCC strains and each plasma sample were mixed (6 h, 37°C). Bacteria growth was determined by measuring the absorbance at 630 nm. Each bar represents the mean ± S.E.M. (n = 4). **, p < 0.01 versus normal rats. B, the measurement of CH50in normal healthy rats and hemorrhagic shock rats at 4 or 7 days after HbV or wRBC resuscitation. Plasma samples were obtained at 4 and 7 days after resuscitation with HbV and wRBC. Each bar represents the mean ± S.D. (n = 4). **, p < 0.01 versus normal rats. C, relationship between CH50 and bacterial growth. The linear regression was calculated using the least-squares method (y = 0.892x + 64.9, r = 0.47, p = 0.013).
The Relationship between CH50 and Antibacterial Activity.
In a previous study, we reported that CH50 in plasma was decreased after resuscitation with HbV in the rat model of hemorrhagic shock (Taguchi et al., 2011a). Because it is well known that complement represents the first line of the host defense system and possesses bacteriolytic effects, we examined the relationship between the antibacterial activity described above and CH50 in plasma.
As shown in the previous results, the CH50 for HbV4day was significantly smaller than that in normal rats, whereas the other samples remained unchanged (p < 0.01; Fig. 1B). As shown in Fig. 1C, CH50 was well correlated with antibacterial activity (r = 0.47, p = 0.013). These results strongly suggest that CH50 is associated with the observed antibacterial activity.
The Relationship between Plasma IgM and Antibacterial Activity.
Because Ishida and Kiwada (2008) demonstrated that the IgM produced from spleen is related to the reduction of CH50 after the administration of liposomes, it is possible that the change in plasma IgM levels after HbV administration might affect antibacterial activity via the reduction of CH50.
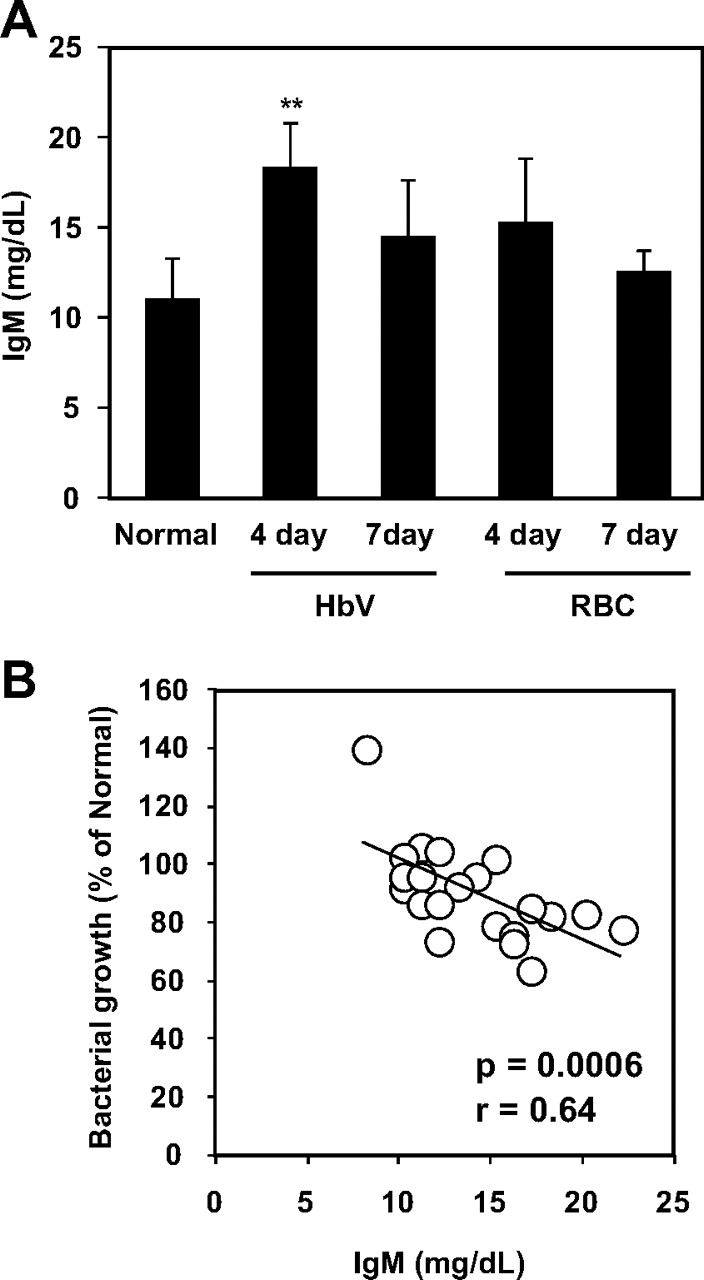
As shown in Fig. 2A, the plasma IgM level in the HbV4day was significantly larger than that in normal rats, but the other plasma samples were not significantly changed (p < 0.01; Fig. 3A). Similar to CH50 (Fig. 1B), plasma IgM levels were well correlated with the observed antibacterial activity (r = 0.64, p = 0.0006; Fig. 2B). To further demonstrate this relationship, we prepared splenectomized rats to suppress the production of IgM by the administration of HbV, and the same experiments were then performed. The level of plasma IgM and CH50 were maintained at the normal rat level by removing the spleen; even in 4 days after resuscitation with HbV (Fig. 3, A and B), no significant antibacterial activity was detected in any of the splenectomized rats (Fig. 3C). Consequently, no significant correlation was found between either plasma IgM and antibacterial activity or CH50 and antibacterial activity (r = 0.18 and 0.03, for plasma IgM and complement activity, respectively) (data not shown).

A, the measurement of plasma IgM concentrations in normal healthy rats and hemorrhagic shock rats at 4 or 7 days after HbV or wRBC resuscitation. Plasma samples were obtained at 4 and 7 days after resuscitation with HbV and wRBC solution. The plasma samples were then ultracentrifuged to remove intact HbV (50,000g, 30 min). Each bar represents the mean ± S.D. (n = 4). **, p < 0.01 versus normal rats. B, relationship between plasma IgM concentration and bacterial growth. The linear regression was calculated using the least-squares method (y = −2.77x + 129.8, r = 0.64, p = 0.0006).

A and B, the measurement of plasma IgM concentration (A) and CH50 (B) in normal healthy rats and hemorrhagic shock rats at 4 or 7 days after HbV or wRBC resuscitation in splenectomy rats. Plasma samples from splenectomy rats were obtained 4 and 7 days after resuscitation with HbV and wRBC solutions. The plasma samples were then ultracentrifuged to remove intact HbV (50,000g, 30 min). Each bar represents the mean ± S.D. (n = 4). C, the antibacterial (E. coli ATCC strains) activity of plasma obtained from splenectomy rats. E. coli ATCC strains and each plasma sample obtained from splenoctomized rats were mixed (6 h, 37°C). Bacteria growth was determined by measuring the absorbance at 630 nm. There was no significant difference among groups. Each bar represents the mean ± S.E.M. (n = 4).
Evaluation of Antibacterial Activity in the Two-Hit Model of Hemorrhagic Shock and Sepsis Rat Model Induced by Cecum Ligation and Puncture.
To examine whether the antibacterial activity caused by resuscitation with HbV were observed under conditions of bacterial infection in vivo, we prepared a double model of hemorrhagic shock and cecum ligation and puncture, as a model for sepsis, arguably one that closely replicates the nature and course of clinical sepsis in patients after trauma (Hubbard et al., 2005).
Figure 4A shows the survival rates after the induction of cecum ligation and puncture. Ninety percent of the normal rats died within 36 h after the induction of cecum ligation and puncture. However, the survival times for HbV7day, RBC4day, and RBC7day rats were prolonged, but 90% of these died within 120 h. It is noteworthy that a significant number of animals at HbV4day survived compared with normal rats, and 40% of the rats in this group survived until 168 h after the induction of cecum ligation and puncture. We also measured the amount of bacteria in blood at 24 h after the induction of cecum ligation and puncture. As shown in Fig. 4B, the amount of bacteria in blood decreased in the HbV4day and HbV7day rats compared with the normal and RBC resuscitation groups. To investigate whether the amount of bacteria in blood had an influence on survival rate, we compared the numbers of bacteria in blood between the rats that survived for more than 72 h and the rats that died within 72 h. As shown in Fig. 4C, the amount of bacteria in the blood of the rats that survived more than 72 h was less than that in the rats that died within 72 h. These results indicate that the elevated complement activity after HbV administration contributed to the removal of bacteria, even under conditions of severe sepsis, and the clearance of bacteria at an earlier time was an important factor in the improved survival rates.

A, survival rate of the two-hit model of hemorrhagic shock and sepsis. Sepsis was induced by cecum ligation and puncture. The cecum was ligated, and then 10 punctures in the cecum ligation were performed using a 21-gauge needle. The survival rate was compared using Kaplan-Meier survival curves and the log-rank test. *, p < 0.05 versus normal rats (n = 10). B, amount of bacteria in blood at 24 h after cecum ligation and puncture. C, the comparison of the amount of bacteria in blood at 24 h after cecum ligation and puncture between rats that survived over 72 h and rats that died within 72 h after the induction of sepsis. Blood samples from survival rats (0.3 ml) were collected from the tail vein 24 h after puncture. After collecting the blood samples, aliquots (0.1 ml) were immediately placed on Luria-Bertani agar plates (9.9 ml) followed by incubation at 37°C for 24 h. The numbers of bacterial colonies were then counted and expressed as colony-forming units (CFU)/ml. Each bar represents the mean ± S.D.
Phagocyte Activity.
Phagocyte activity, especially in Kupffer cells, is strongly related to the removal of bacteria in vivo. Thus, to investigate the effect of resuscitation by HbV and wRBC solutions on phagocyte activity in Kupffer cells, we estimated the in vivo carbon clearance, an indicator of phagocyte activity in Kupffer cells [Kupffer cells engulfed more than 90% of the injected carbon particles (Zweifach and Benacerraf, 1958)].
As shown in Table 1, phagocyte activity in HbV4day and HbV7day rats was approximately 1.5 and 1.9 times higher than that in normal rats. In contrast, phagocyte activity for both RBC4day and RBC7day rats was comparable with that in normal rats. These data indicate that phagocyte activity was increased after resuscitation with HbV in the rat model of hemorrhagic shock, and the enhanced phagocyte activity played a role in the clearance of bacteria in vivo.
Phagocyte activity in hemorrhagic shock rats at 4 or 7 days after HbV or wRBC resuscitation
Carbon clearance was estimated, and K was calculated from the clearance of carbon particles. Results represent the mean ± S.D. (n = 5).
Discussion
The “two-hit” theory proposes that a host primed by an initial stress such as a massive hemorrhage is likely to show an abnormal response to a second stress such as an infection (Price et al., 1999). It has been shown that bacterial translocation from the gut to mesenteric lymph nodes induced by hemorrhagic shock, in combination with the immunosuppressive effects of hemorrhage, can amplify the second hit (Turnbull et al., 1995). In fact, Rachoin et al. (2009) reported that the approximately 10% of transfused patients developed nosocomial infections, and the number of patients infected with E. coli was similar to that of methicillin-resistant Staphylococcus areus. E. coli is classed as a gram-negative bacteria, in which the outer membrane contains a lipopolysaccharide as endotoxin, which causes endotoxin shock (sepsis) and multiple organ failure (Yang and Lee, 2008). Therefore, in terms of decreasing mortality and morbidity among transfused patients with massive hemorrhage, the removal of Gram-negative bacteria, especially E. coli, at an early phase is an important issue.
The findings reported herein showed that plasma of rats resuscitated with HbVs from hemorrhagic shock showed a suppressed E. coli growth correlated to reduction of CH50 (Fig. 1). Four possible mechanisms for the reduction of CH50 after HbV resuscitation can be proposed: 1) The liver injury decreases the production of complement; however, it was reported that HbVs did not directly injure the liver and other organ. Therefore, it would affect the production of complement. 2) The direct effect of HbV, which makes contact with complement, subsequently induces the complement activation, because it was previously reported that liposomes containing cholesterol induce complement activation (Alving et al., 1977; Cunningham et al., 1979). However, in the case of HbV, the level of CH50 was equivalent to that observed for saline treatment in both in vitro studies using human serum (Abe et al., 2006) and in vivo using healthy rats (Sou and Tsuchida, 2008). In addition, E. coli incubated with HbVs did not show antibacterial activity (data not shown). Therefore, it does not seem the HbV itself directly affects the reduction of CH50. 3) Plasma IgG, which activates complement by binding to an antigen, recognizes microbial polysaccharides, and by the lipid A component of lipopolysaccharide. However, plasma IgG was hardly induced by HbV administration, and no correlation with the antibacterial activity was found (r = 0.09, data not shown). 4) Plasma IgM, which activates complement via the same pathway as plasma IgG, as mentioned above. In this study, the plasma IgM produced in spleen was well correlated with E. coli growth (Fig. 2). Furthermore, it was observed that both CH50 and antibacterial activity disappeared by suppression of IgM production using splenoctomized rats (Fig. 3). Based on these results, it seems that complement is activated by binding IgM to E. coli, and this complement activity would result in suppressed E. coli growth.
The complement system contributes to host defense against infections by two different mechanisms: C3-mediated lysis, which leads to the formation of a membrane attack complex (Frank et al., 1987), and opsonic activity, which leads to phagocytosis by macrophages and Kupffer cells (Esser, 1994). Therefore, C3 is a key factor for antibacterial activation. In fact, a clinical C3 deficiency involves the loss of major complement opsonin and failure to activate the membrane attack complex pathway (Walport, 2001). It was previously reported that when liposomes are repeatedly injected into the same animal IgM is induced by the first injected liposomes and the binding of IgM to the second injected liposomes occurs, followed by C3 activation by IgM (Ishida and Kiwada, 2008; Taguchi et al., 2009c). Therefore, C3 could also be activated by the binding of IgM to E. coli in the present study.
In addition, we used a double-hit model of hemorrhagic shock and sepsis, which closely replicates the nature and course of clinical sepsis in patients after trauma (Hubbard et al., 2005). Under these conditions, only 10% of the normal and RBC resuscitation rats survived 168 h after the induction of cecum ligation and puncture, whereas 40% of the HbV4day animals survived (Fig. 4A). Previously, Turnbull et al. (2004) reported that antimicrobial therapy with the broad-spectrum antibiotic imipenem at a dose of 25 mg/kg resulted in a significant improvement in survival rate (46%) compared with nontreatment (26%) in cecum ligation and puncture model mice. Furthermore, a imipenem-cilastatin treatment, administered 500 μg intraperitoneally every 12 h, prolonged the survival time in cecum ligation and puncture model mice, but did not prevent mortality (100% mortality; 84 and 120 h, for cecum ligation and puncture alone and imipenem-cilastatin treatment groups, respectively) (Doerschug et al., 2004). Therefore, it might be expected that complement activation by IgM after HbV resuscitation would achieve a full therapeutic effect for antibacterial activity, similar to antibiotic therapy.
In addition to the enhancement in survival rate, the amount of bacteria in blood of the HbV resuscitation groups was less than those of the other groups (Fig. 4B). In ex vivo conditions, although the HbV7day group did not show antibacterial activity, the amount of bacteria in blood was suppressed. Because the mononuclear phagocyte system, especially Kupffer cells, plays a major role in the clearance of microorganisms (Bilzer et al., 2006), the suppression of bacterial count in the HbV7day group seems to be the result of increased phagocyte activity. Actually, phagocyte activity in the HbV7day rats was approximately 1.9 times higher than that in normal rats (Table 1).
It might be a concern that the elevated complement activity induced by IgM after HbV resuscitation damaged some host organisms. However, when the plasma samples were added to human umbilical vein endothelial cells, which are commonly used as a model of vascular endothelial cells, to examine cellular injury using WST-8, no significant difference in human umbilical vein endothelial cell viability was found among the groups at 6 or 24 h (data not shown). Therefore, it is unlikely that the observed complement activation damages host organisms. In fact, it was reported that histological changes and an enhancement in organ injury markers were not observed in the hemorrhagic shock rat model after resuscitation by HbV (Sakai et al., 2009a). Moreover, complement activation is a subject of great concern as a pseudoallergic reaction (Laverman et al., 2001). In this study, no allergic-like reactions, such as anaphylactic reactions, were observed, and previous reports demonstrated that healthy rats, which were repeatedly injected a massive dose of HbV (10 ml/kg/day for 14 days), all survived and showed no toxicity (Sakai et al., 2004a). Therefore, an enhancement in complement activity after HbV injection would not be expected to induce allergic reactions or have an effect on host organisms. However, it is possible that the HbV administration might produce detrimental outcome in conditions in which complement activation increases the risk of survival, because HbV enhanced the complement activation. In the case of using HbV clinically, resuscitation should be the first priority as in hemorrhagic shock that decreased the survival rate. Having said that, the effect of complement activation via HbV administration in conditions in which complement activation increases the risk of survival deserves further investigation.
Although the complement activation induced by IgM after HbV administration showed antibacterial activity, our model has several limitations with respect to providing a complete explanation for antibacterial activity. It is well known that reactive oxygen species (ROS) and cytokines, such as interleukin-6, tumor necrosis factor-α, and C-reactive protein, are also related to antibacteria action. In this study, the effect of ROS was estimated by determining the oxidative albumin ratio, which serves as a nonspecific marker of ROS (Shimoishi et al., 2007). As a result, we found that the relationship between ROS and antibacterial activity was significantly correlated (r = 0.42, p < 0.05; data not shown). Therefore, it is possible that ROS and cytokines are also involved in the antibacteria action observed here. However, further study will be needed to clarify the contribution of these factors. Moreover, in clinics, patients frequently become infected with various species of bacteria including Gram-negative and -positive bacteria, but we used only Gram-negative bacteria, E. coli. In general, Gram-positive bacteria are less sensitive to lysis by complement than Gram-negative bacteria (Mold, 1999). However, because Gram-positive bacteria affected the opsonization by complement, antibacterial activity of Gram-positive bacteria under in vivo conditions would not be unexpected. Further study will be needed to demonstrate this fact.
Based on the present findings, resuscitation with HbV in a rat model of hemorrhagic shock suppressed the growth of E. coli via complement activation induced by IgM, compared with wRBC resuscitation. However, this possible therapeutic efficacy was limited to 4 days after resuscitation with HbV. Fortunately, in clinical settings, the timing of infections from the first transfusion is typically approximately 4 to 5 days, which is consistent with the present results (Rachoin et al., 2009). Given the above findings and ongoing progress in antibacterial therapy, HbV resuscitation may contribute to a reduction of infections in patients with a massive hemorrhage, especially in cases of inadequately resourced health service areas.
Authorship Contributions
Participated in research design: Taguchi, Watanabe, Maruyama, and Otagiri.
Conducted experiments: Taguchi and Ogaki.
Contributed new reagents or analytic tools: Sakai, Kobayashi, and Horinouchi.
Performed data analysis: Taguchi and Kadowaki.
Wrote or contributed to the writing of the manuscript: Taguchi, Sakai, Maruyama, and Otagiri.
Acknowledgments
We thank Emeritus Prof. Eishun Tsuchida for support regarding this research.
Footnotes
This work was supported, in part, by Health Sciences Research Grants from the Ministry of Health, Labor, and Welfare of Japan.
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
doi:10.1124/jpet.110.177832.
-
ABBREVIATIONS:
- HbV
- hemoglobin vesicle
- RBC
- red blood cell
- wRBC
- washed RBC
- HbV4day
- 4 days after resuscitation by HbVs
- HbV7day
- 7 days after resuscitation by HbVs
- RBC4day
- 4 days after resuscitation by RBCs
- RBC7day
- 7 days after resuscitation by RBCs
- CH50
- complement titer
- ROS
- reactive oxygen species
- SD
- Sprague-Dawley.
- Received December 6, 2010.
- Accepted January 10, 2011.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}