


Abstract
Organic anion-transporting polypeptides 1B1 and 1B3 (OATP1B1 and OATP1B3) are liver-specific transporters that mediate the uptake of a broad range of drugs into hepatocytes, including statins, antibiotics, and many anticancer drugs. Compounds that alter transport by one or both of these OATPs could potentially be used to target drugs to hepatocytes or improve the bioavailability of drugs that are cleared by the liver. In this study, we applied a bioassay-guided isolation approach to identify such compounds from the organic extract of Rollinia emarginata Schlecht (Annonaceae). Fractions of the plant extract were screened for effects on OATP1B1- and OATP1B3-mediated transport of the model substrates estradiol-17β-glucuronide and estrone-3-sulfate. We isolated three compounds, ursolic acid, oleanolic acid, and 8-trans-p-coumaroyloxy-α-terpineol, which inhibited estradiol-17β-glucuronide uptake by OATP1B1 but not OATP1B3. In addition, a rare compound, quercetin 3-O-α-l-arabinopyranosyl(1→2) α-l-rhamnopyranoside, was identified that had distinct effects on each OATP. OATP1B1 was strongly inhibited, as was OATP1B3-mediated transport of estradiol-17β-glucuronide. However, OATP1B3-mediated uptake of estrone-3-sulfate was stimulated 4- to 5-fold. Kinetic analysis of this stimulation revealed that the apparent affinity for estrone-3-sulfate was increased (decreased Km), whereas the maximal rate of transport (Vmax) was significantly reduced. These results demonstrate a mechanism through which the hepatic uptake of drug OATP substrates could be stimulated.
Introduction
Organic anion-transporting polypeptides (OATPs) are multispecific transporters that mediate the uptake of a broad range of drugs and other xenobiotics into cells. Two members of this superfamily, OATP1B1 and OATP1B3, are selectively expressed on the basolateral membrane of hepatocytes under normal physiological conditions. Among the substrates of OATP1B1 and OATP1B3 are numerous drugs used to treat hypertension, hypercholesterolemia, and diabetes (Hagenbuch and Gui, 2008).
Because of the localization of OATP1B1 and OATP1B3 at the basolateral membrane of hepatocytes, these transporters can have a significant impact on the first-pass effect of orally administered drugs. Alterations in the function of these OATPs have been shown to affect the disposition of drugs throughout the body. Polymorphisms in OATP1B1 have been associated with altered pharmacokinetics of several of the HMG-CoA reductase inhibitors (statins). Patients with OATP1B1*1b have lower plasma levels of pravastatin and pitavastatin than those with the *1a variant, whereas the *15 variant is associated with higher plasma concentrations (Nishizato et al., 2003; Chung et al., 2005; Maeda et al., 2006). OATPs have also been implicated as a cause of several known drug-drug interactions, including those between cerivastatin and gemfibrozil (Shitara et al., 2003), rosuvastatin and cyclosporine (Simonson et al., 2004), and fexofenadine and grapefruit juice (Bailey et al., 2007). This suggests that interactions at OATP1B1 or OATP1B3 may alter the disposition of drugs throughout the body.
Coadministration of a drug with a small molecule that alters transporter function can be used therapeutically to improve drug bioavailability or distribution. The bioavailability of orally administered paclitaxel and docetaxel is improved by coadministration with cyclosporine, which inhibits P-glycoprotein-mediated efflux from enterocytes (Meerum Terwogt et al., 1998; Malingré et al., 2001). Likewise, coadministration of the breast cancer resistance protein and P-glycoprotein inhibitor N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide (GF120918) with topotecan increased systemic bioavailability 2-fold when dosed orally (Kruijtzer et al., 2002). Just as inhibiting efflux from enterocytes can increase systemic bioavailability, so could inhibiting the first-pass effect of uptake into the liver. Selective inhibition of OATP1B1 or OATP1B3 could therefore be used to increase the systemic bioavailability of OATP drug substrates. Conversely, selective stimulation of OATP1B1 or OATP1B3 uptake could be used to increase drug delivery to hepatocytes.
Among the many OATP substrates are several anticancer drugs, such as docetaxel and paclitaxel (OATP1B3) (Smith et al., 2005), methotrexate (OATP1B1 and OATP1B3) (Abe et al., 2001), and the active metabolite of irinotecan, 7-ethyl-10-hydroxy-camptothecin (SN-38) (OATP1B1) (Nozawa et al., 2005). Thus, stimulation of OATP1B1 or OATP1B3 transport activity could also be a method of improving cancer treatments. OATP1B1 and OATP1B3 have also been identified in certain cancer tissues, including cancers of the breast, colon, lung, pancreas, prostate, and stomach (Abe et al., 2001; Monks et al., 2007; Muto et al., 2007; Hamada et al., 2008). Most of these studies have identified only mRNA expression, and neither membrane localization nor function has been shown. However, if OATPs are expressed and functional on the surface of these cancer cells, stimulation of OATP function could be a promising technique for increasing uptake of cytotoxic drugs into cancer cells.
So far, specific modulators of OATPs are limited. One method for identifying additional specific modulators is using high-throughput screening. A recent high-throughput screening method for detecting OATP1B3 modulators identified several compounds that preferentially modulated OATP1B1 or OATP1B3; however, this assay is not sufficiently stringent for OATP1B1 (Gui et al., 2010). Another way to identify modulators is through bioassay-guided fractionation. This is a technique frequently used to identify active compounds from plants. Plants are a potentially rich source of OATP modulators: herbal extracts used in dietary supplements have been found to affect transport by OATP1B1, OATP1B3 (Roth et al., 2011), and OATP2B1 (Fuchikami et al., 2006), and interactions between OATPs and fruit juices are well documented (Dresser et al., 2002; Bailey et al., 2007; Glaeser et al., 2007; Greenblatt, 2009).
In this study, we applied a bioassay-guided isolation approach to identify OATP modulators from Rollinia emarginata Schlecht (Annonaceae), a plant that grows in several regions of South America. This plant extract was a positive hit during an initial screening of several South American plant extracts for their effect on OATP-mediated transport. The stem barks have been used in combination with Ilex paraguayensis St Hilaire (Aquifoliaceae) (common name: hierba mate) as a migraine treatment and a relaxant. In addition, antiprotozoal and antifeedant properties have been reported (Février et al., 1999; Colom et al., 2007). Fractions of plant extract were screened for their effects on OATP1B1- and OATP1B3-mediated uptake of the two model substrates, estradiol-17β-glucuronide and estrone-3-sulfate.
Materials and Methods
Materials.
Above-ground plant material of R. emarginata was collected and identified in February 1999 in Argentina by R. Fortunato and A. Cabral (Instituto Nacional de Tecnología Agropecuaria) (collection ARP 613; latitude 25°14′0′5 south; longitude 57°57′0′0 west; RN 86, 2 km northeast of Patino, Department Primavera, Province Formosa). [3H]estrone-3-sulfate (54.3 Ci/mmol) and [3H]estradiol-17β-glucuronide (41.8 Ci/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Waltham, MA). Unlabeled estrone-3-sulfate, estradiol-17β-glucuronide, and rifampicin were purchased from Sigma-Aldrich (St. Louis, MO).
Plant Extraction and Isolation.
Dried and ground plant material (562 g) was extracted with methanol (MeOH) and dichloromethane (CH2Cl2) mixture (1:1, v/v) three times for 24-h periods at room temperature. Organic solvents were removed in vacuo at 35°C; the residue was suspended in MeOH/H2O (9:1, v/v) and partitioned with hexanes (HEX fraction). After removal of MeOH, the aqueous layer was extracted successively with dichloromethane (DCM fraction) and butanol (BUOH fraction). The HEX fraction was then subjected to silica gel column chromatography (Si-Gel CC) (32–64 μm, 36 × 460 mm) and eluted with a gradient of hexanes-ethyl acetate (EtOAc) (20:1 to 0:100, v/v) to afford 20 subfractions (A to T), which were combined according to thin-layer chromatography analysis. Subfraction HEX-G (310 mg) was submitted to Si-Gel CC (12–26 μm, 36 × 230 mm) using a gradient of hexanes and acetone (15:1 to 5:1, v/v) to obtain three subfractions (G1-G3). Subfraction HEX-G1 (205 mg) was purified using Si-Gel CC (CH2Cl2/EtOAc, 20:1, v/v) to afford compound 3 (120 mg). Subfraction HEX-G2 was purified with Si-Gel CC (12–16 μm, 20 × 460 mm) using hexanes, CH2Cl2, and methyl tert-butyl ether (20:15:1, v/v/v) as mobile phase to yield compound 4 (10.4 mg). In addition, subfraction HEX-N (284 mg) was separated using Si-Gel CC (32–64 μm, 36 × 230 mm) and CH2Cl2-EtOAc (10:1, v/v) as a solvent system to yield a mixture of compounds 1 and 2 (103 mg), which was resolved by means of semipreparative HPLC (reverse-phase C-18, 10 × 250 mm, 5 μm; solvent A, acetonitrile; solvent B, water, gradient: 80% A to 100% A in 45 min). Fraction BUOH (19.7 g) was subject to MCI-Gel CHP20P (Sigma-Aldrich) CC (65 × 350 mm) and eluted with various mixtures of water and MeOH (100:0, 25:75, 50:50,75:25, and 0:100; v/v) to afford four fractions (A-D). Subfraction BUOH-B (2.2 g) was submitted to Sephadex LH-20 (GE Healthcare Chalfont St. Giles, Buckinghamshire, UK), with MeOH as a mobile phase. A total of 180 fractions (7.5 ml each) were collected and combined into nine fractions (1–9) after thin-layer chromatography analysis. Pigments present in fraction BUOH-B7 (530 mg) were removed with a small Si-Gel plug using CH2Cl2/MeOH/H2O (4:1:0.1, v/v/v) as eluent to obtain a mixture of compounds 5 and 6 (450 mg). A portion of this mixture (40 mg) was purified using semipreparative HPLC (reverse-phase C-18, 10 × 250 mm, 5 μm; solvent A, acetonitrile; solvent B, water, isocratic 18% A) to afford compounds 5 (24 mg) and 6 (6.2 mg).
Compound Identification.
The structures of isolated compounds were established by one- and two-dimensional NMR experiments and compared with those in literature (Seebacher et al., 2003; Zhang et al., 2005; Muzitano et al., 2006; Faini et al., 2007); IR, UV, and high-resolution mass spectrometry were also in agreement with the proposed structures. NMR experiments were performed in a Bruker (Newark, DE) AVIII 500 instrument with a dual C/H cryoprobe. Standard 1H-NMR, 13C-NMR, correlation spectroscopy, heteronuclear single quantum correlation, and heteronuclear multiple-bond correlation spectroscopy experiments were recorded of each of the pure compounds. An Agilent 1200 system with a 6300 Series Ion Trap detector (Agilent Technologies, Santa Clara, CA) was used for liquid chromatography/mass spectrometry experiments. An Agilent RP-C18 (15 × 4.1 mm, 5 μm) column was used, and different gradients of acetonitrile and water were applied as a mobile phase depending on polarity of samples. High-resolution mass spectrometry was obtained with a LCT Premier (Waters, Milford, MA). The purity of each compound was determined to be more than 95% by HPLC analysis.
Cell Culture.
Chinese hamster ovary cells stably transfected with OATP1B1 or OATP1B3 were cultured as described previously (Gui et al., 2008). Cells were seeded on 24- or 96-well plates and grown to visual confluence (48–72 h). When confluent, medium was exchanged for nonselective medium containing 5 mM sodium butyrate to nonspecifically induce gene expression. Uptake experiments were performed 24 h after induction.
Transport Assays.
Transport experiments were performed essentially as described previously (Gui et al., 2008). Cells were washed three times with prewarmed uptake buffer (116.4 mM NaCl, 5.3 mM KCl, 1 mM NaH2PO4, 0.8 mM MgSO4, 5.5 mM d-glucose, and 20 mM HEPES, pH adjusted to 7.4 with Trizma base). Cells were then incubated with prewarmed uptake buffer containing the radiolabeled substrate. To stop uptake, the substrate solution was aspirated and the cells were washed four times with ice-cold uptake buffer. Cells were lysed with 1% Triton X-100 in phosphate-buffered saline; lysate was used for liquid scintillation counting and protein determination using the bicinchoninic acid assay.
Calculation and Statistics.
All calculations were performed using Prism (GraphPad Software Inc., San Diego, CA). Determination of IC50 values and kinetic parameters was performed within the initial linear period of uptake (20 s). Statistical significance was determined with two-way analysis of variance followed by Bonferroni post-test or two-tailed paired t test.
Results
Identification of Compounds with Modulating Effects on OATP1B1 and OATP1B3.
To identify components of R. emarginata that affect OATP1B1 and OATP1B3 function, the organic components were extracted with MeOH:CH2Cl2, and fractionated with various solvents (Fig. 1). Fractions were solubilized in DMSO, and functional assays were performed in triplicate on 96-well plates. Two model substrates, estradiol-17β-glucuronide (OATP1B1: Km, 5.4 μM; OATP1B3: Km, 15.8 μM) and estrone-3-sulfate (OATP1B1 high-affinity component: Km, 0.22 μM; OATP1B3: Km, 58 μM) (Gui et al., 2008; Gui and Hagenbuch, 2009) were used to identify compounds that have substrate-dependent effects on transport. Active fractions were identified by coincubating wild-type or OATP-expressing cells with uptake buffer containing 0.03 μg/ml R. emarginata extracts and 0.1 μM estradiol-17β-glucuronide or 1 μM estrone-3-sulfate for 5 min at 37°C. Results from selected fractions are shown in Fig. 2. The whole plant extract inhibited uptake of both substrates by both transporters; detannification increased the inhibitory effect, possibly because of increased availability of formerly tannin-bound compounds (data not shown). The hexane and butanol fractions both showed preferential inhibition of OATP1B1-mediated transport of estradiol-17β-glucuronide (Fig. 2A) and were further fractionated. An active subfraction of the hexane fraction (data not shown) contained four compounds, which were identified as ursolic acid (compound 1), oleanolic acid (compound 2), β-sitosterol (compound 3), and 8-trans-p-coumaroyloxy-α-terpineol (compound 4). Ursolic acid, oleanolic acid, and compound 4 inhibited OATP1B1 transport of estradiol-17β-glucuronide by more than 50% while having a minimal effect on OATP1B1 transport of estrone-3-sulfate (Fig. 2). The butanol fraction contained a subfraction (Frac B) that strongly stimulated uptake of estrone-3-sulfate by OATP1B3, while inhibiting uptake of estradiol-17β-glucuronide by both OATPs. Frac B was found to consist mainly of two structurally similar compounds: rutin (compound 5) and quercetin 3-Ο-α-l-arabinopyranosyl (1→2) α-l-rhamnopyranoside (compound 6; Fig. 3). Compound 6 stimulated uptake of estrone-3-sulfate by OATP1B3, while inhibiting uptake of estrone-3-sulfate by OATP1B1 (Fig. 2B) and uptake of estradiol-17β-glucuronide by both transporters (Fig. 2A). To further examine the effects of these compounds on OATP-mediated transport, we purified ursolic acid, oleanolic acid, compound 4, and compound 6 to more than 95% by HPLC analysis and used these purified compounds for all additional experiments.

Separation diagram of R. emarginata organic extract.
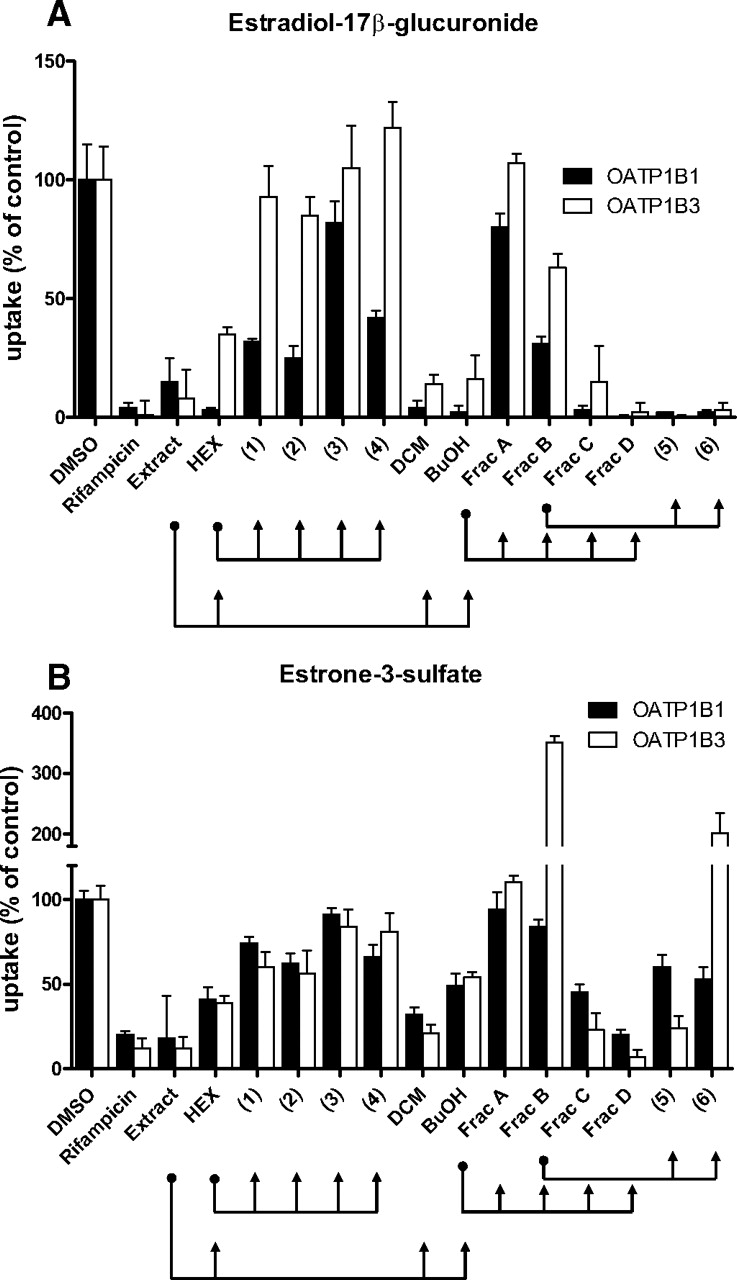
Effect of R. emarginata extract and fractions on OATP1B1- and OATP1B3-mediated uptake. Cells were coincubated with 0.1 μM [3H]estradiol-17β-glucuronide (A) or 1 μM [3H]estrone-3-sulfate (B) and 0.03 μg/ml of plant extract or fraction or 100 μM pure compound (1–6). After subtracting the values obtained in wild-type cells from those obtained by OATP1B1- or OATP1B3-expressing cells, net transporter-mediated uptake was expressed as a percentage of the vehicle control (1% DMSO). Each value is the mean ± S.D. from one experiment done in triplicate. Separation steps from initial extract to pure compounds are indicated by arrows.

Chemical structures of OATP modulators isolated from R. emarginata butanol fraction. Compounds are identified as 5 and 6.
Ursolic Acid, Oleanolic Acid, and 8-trans-p-Coumaroyloxy-α-Terpineol Selectively Inhibit OATP1B1-Mediated Transport of Estradiol-17β-Glucuronide.
To examine the selectivity of inhibition of the three compounds isolated from the hexane fraction, OATP1B1- and OATP1B3-mediated transport of 0.1 μM estradiol-17β-glucuronide or estrone-3-sulfate was measured for 20 s at 37°C in the presence of 100 μM ursolic acid, oleanolic acid, compound 4, or 1% DMSO (vehicle control). All three compounds significantly inhibited uptake of estradiol-17β-glucuronide by OATP1B1 (p < 0.001), while having no effect on uptake by OATP1B3 (Fig. 4A). Compound 4 had a similar effect on uptake of estrone-3-sulfate, inhibiting OATP1B1- but not OATP1B3-mediated transport (Fig. 4B). However, uptake of estrone-3-sulfate by both transporters was inhibited to an equal extent by ursolic acid and oleanolic acid (Fig. 4B). Inhibition of estradiol-17β-glucuronide transport by OATP1B1 was further studied with a concentration dependence. Ursolic acid and oleanolic acid inhibited uptake of estradiol-17β-glucuronide with IC50 values of 15.3 μM (Fig. 5A) and 4.2 μM (Fig. 5B). Compound 4 was the weakest inhibitor; the full plateau of inhibition could not be determined because of limited solubility (Fig. 5C).

Effect of ursolic acid, oleanolic acid, and 8-trans-p-coumaroyloxy-α-terpineol on OATP-mediated uptake. Cells were coincubated with 0.1 μM [3H]E17β (A) or [3H]E3S (B) and 100 μM ursolic acid, oleanolic acid, or compound 4 for 20 s. After subtracting the values obtained in wild-type cells from those obtained by OATP1B1- or OATP1B3-expressing cells, net transporter-mediated uptake was expressed as a percentage of the vehicle control (1% DMSO). Each value is the mean ± S.E.M. of three independent experiments, each performed in triplicate. ***, statistically significant differences (P < 0.001) from the vehicle control. †††, statistically significant differences (P < 0.001) between OATP1B1 and OATP1B3.

Concentration-dependent effect of ursolic acid, oleanolic acid, and compound 4 on OATP1B1-mediated uptake of estradiol-17β-glucuronide. Cells were coincubated with 0.1 μM [3H]E17β and increasing concentrations of ursolic acid (A), oleanolic acid (B), or compound 4 (C) for 20 s. After subtracting the values obtained in wild-type cells from those obtained by OATP1B1-expressing cells, net transporter-mediated uptake was expressed as a percentage of the vehicle control (1% DMSO). Each value shown is the mean ± S.E.M. of three independent experiments, each performed in triplicate.
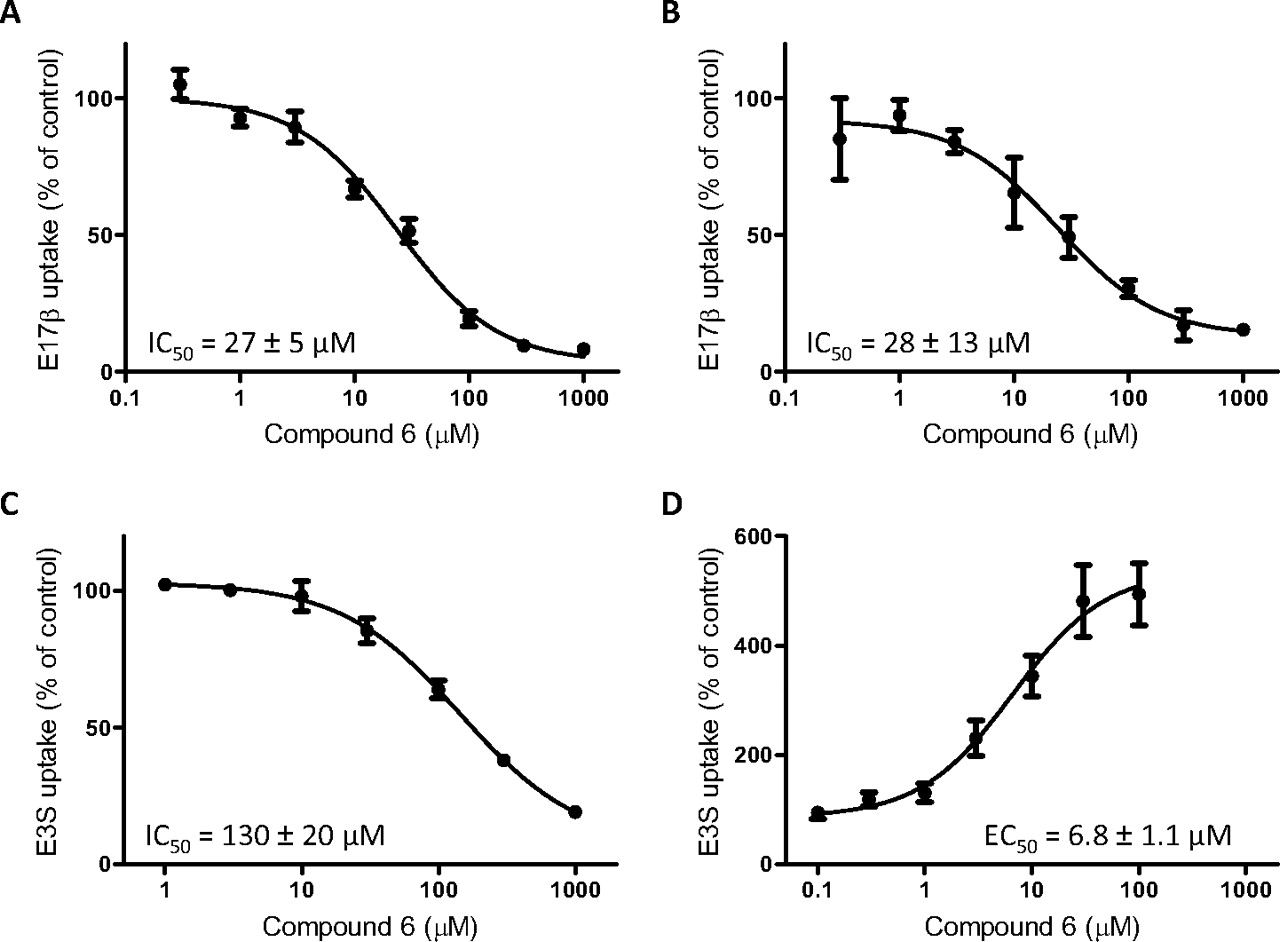
Quercetin 3-Ο-α-l-Arabinopyranosyl (1→2) α-l-Rhamnopyranoside Has Concentration-Dependent Effects on OATP1B1- and OAPT1B3-Mediated Uptake.
To further investigate the substrate-dependent effects of compound 6, uptake of 0.1 μM estradiol-17β-glucuronide or estrone-3-sulfate by OATP1B1 and OATP1B3 was measured for 20 s at 37°C in the presence of increasing concentrations of compound 6. Uptake of estradiol-17β-glucuronide was inhibited by compound 6 to a similar extent for both transporters (Fig. 6, A and B). OATP1B1-mediated uptake of estrone-3-sulfate was inhibited to a lesser extent (IC50, 130 μM; Fig. 6C). The stimulation of OATP1B3-mediated uptake of estrone-3-sulfate was also concentration-dependent, with an EC50 of 6.8 μM (Fig. 6D). At concentrations higher than 100 μM, the effect of compound 6 on estrone-3-sulfate decreased, although it remained stimulatory to the highest tested concentration of 1 mM compound 6 (data not shown).

Concentration-dependent effect of compound 6 on OATP-mediated uptake of estradiol-17β-glucuronide and estrone-3-sulfate. Cells were coincubated with 0.1 μM [3H]E17β (A and B) or [3H]E3S (C and D) and increasing concentrations of compound 6 for 20 s. After subtracting the values obtained in wild-type cells from those obtained by OATP1B1-expressing cells (A and C) or OATP1B3-expressing cells (B and D), net transporter-mediated uptake was expressed as a percentage of the vehicle control (1% DMSO). Each value shown is the mean ± S.E.M. of three independent experiments, each performed in triplicate.
Quercetin 3-Ο-α-l-Arabinopyranosyl (1→2) α-l-Rhamnopyranoside Decreases OATP1B3's Maximal Transport Rate for Estradiol-17β-Glucuronide and Estrone-3-Sulfate and Increases its Affinity for Estrone-3-Sulfate.
To characterize the mechanism by which these four compounds interact with OATP1B1 and OATP1B3, we examined their effects on the kinetic parameters of OATP1B-mediated transport. Kinetic analysis of estradiol-17β-glucuronide and estrone-3-sulfate uptake was performed in the presence of each interacting compound or the vehicle control, and results are shown in Table 1. The affinity of estradiol-17β-glucuronide for OATP1B1 was slightly decreased by each of the four substrates tested. The maximal rate of transport (Vmax) was not changed by ursolic acid or compound 4, but was somewhat decreased by both oleanolic acid and compound 6. However, none of these changes reached statistical significance. In initial experiments, we determined that ursolic acid, oleanolic acid, and compound 4 did not alter the low-affinity, high-capacity component of OATP1B1-mediated transport of estrone-3-sulfate (data not shown); therefore only the high-affinity component was studied. As for estradiol-17β-glucuronide transport, all four compounds caused small, but nonsignificant, decreases in substrate affinity, although none altered the maximal rate of transport.
Kinetics of OATP-mediated transport in the absence and presence of modulators
All inhibitors were used at 100 μM unless indicated otherwise.
At the substrate concentrations necessary to determine the kinetics of estrone-3-sulfate uptake by OATP1B3, ursolic acid, and oleanolic acid had no effect on transport. Compound 6, however, significantly altered the kinetic parameters of OATP1B3-mediated transport of both model substrates, as illustrated in Fig. 7. OATP1B3-mediated estradiol-17β-glucuronide uptake was inhibited in a noncompetitive manner (Fig. 7A). Inclusion of 25 or 75 μM compound 6 in the uptake media decreased the maximal rate of transport (Vmax) from 280 ± 45 to 188 ± 40 (not statistically significant) and 83 ± 9 pmol/mg · min (p < 0.05), respectively. Compound 6 had no effect on the apparent affinity (Km) for estradiol-17β-glucuronide (DMSO control: 16 ± 9 μM; 25 μM: 17 ± 5 μM; 75 μM: 18 ± 9 μM). Uptake of estrone-3-sulfate by OATP1B3 was measured in the presence of 50 μM compound 6 or 1% DMSO (Fig. 7B). As was the case with estradiol-17β-glucuronide, the Vmax was decreased, from 2.12 ± 0.34 to 1.07 ± 0.05 nmol/mg · min (p < 0.05). However, the Km was also decreased nearly 10-fold, from 93 ± 38 to 15 ± 3 μM (p < 0.005). This explains the stimulation of transport seen at low estrone-3-sulfate concentrations despite the decrease in Vmax.

Effect of compound 6 on OATP1B3-mediated transport. A, cells were incubated with increasing concentrations of E17β in the presence of 25 μM (■) or 75 μM (▴) compound 6 or the vehicle control (0.5% DMSO; ●) for 20 s. B, cells were incubated with increasing concentrations of E3S in the presence of 50 μM compound 6 (■) or the vehicle control (●) for 20 s After subtracting the values obtained in wild-type cells, net transporter-mediated uptake was fitted to the Michaelis-Menten equation to determine Km and Vmax values. Each value shown is the mean ± S.E.M. of at least three independent experiments, each performed in triplicate.
Discussion
Many naturally occurring plant compounds have been identified that interact with OATPs (Dresser et al., 2002; Fuchikami et al., 2006; Bailey et al., 2007; Glaeser et al., 2007; Greenblatt, 2009), making plants a potentially rich source of OATP modulators. In the present study, we used a bioassay-guided approach to isolate and identify four compounds from the organic extract of R. emarginata Schlecht (Annonaceae) that selectively modulate OATP function. This approach enabled us to quickly focus in on fractions of plant extract-containing compounds that had substrate-specific or transporter-specific effects on OATP function. As illustrated in Fig. 1, only three to four fractionation steps were required to isolate pure compounds with the desired activity from the plant extract. A downfall of this approach is the likelihood of missing OATP-modulating compounds, whose effects may be masked in the early stages of fractionation because of the large number of compounds in each fraction. However, it does allow for the early elimination of many general OATP modulators, increasing the efficiency of this approach compared with that of high-throughput screening of large compound libraries.
We demonstrated that ursolic acid, oleanolic acid, and compound 4 selectively inhibit estradiol-17β-glucuronide transport by OATP1B1 compared with OATP1B3 (Fig. 4A). Several other compounds have previously been identified as selective inhibitors of OATP1B1. Uptake of estradiol-17β-glucuronide by OATP1B1 was inhibited to a greater extent than was uptake by OATP1B3 by both indocyanine green (Cui et al., 2001) and the proposed cardiotonic agent (−)-N-{2-[(R)-3-(6,7-dimethoxy-1,2,3,4-tetrahydroisoquinoline-2-carbonyl)piperidino]ethyl}-4-fluorobenzamide (YM758) (Umehara et al., 2008). Taurocholate uptake by OATP1B1 was inhibited by BAPA-8, BAPA-6, and BAPA-3, glycocholic acid derivatives of bile acid conjugated N-(3-aminopropyl)-1,3-propanediamine, more strongly than was uptake by OATP1B3 (Vicens et al., 2007). However, we found that the selective inhibition of OATP1B1 by ursolic acid and oleanolic acid depended on the substrate being transported. Although compound 4 selectively inhibited OATP1B1 transport of both estradiol-17β-glucuronide and estrone-3-sulfate but had no significant effect on transport by OATP1B3, ursolic acid and oleanolic acid inhibited the uptake of estrone-3-sulfate by both transporters (Fig. 4B). Furthermore, in a previous study using fluorescein-methotrexate, we identified ursolic acid as a preferential OATP1B3 inhibitor (Gui et al., 2010). Ursolic acid inhibited OATP1B3-mediated fluorescein-methotrexate uptake with an IC50 of 2.3 μM, whereas the IC50 value for OATP1B1 was 12.5 μM.
Substrate-dependent modulation of OATP1B3 was also produced by compound 6. Transport of estradiol-17β-glucuronide by both OATP1B1 and OATP1B3 was inhibited to the same extent (Fig. 6, A and B). However, uptake of estrone-3-sulfate by OATP1B1 was inhibited, whereas uptake by OATP1B3 was stimulated (Fig. 6, C and D). Further investigation of this effect on OATP1B3 revealed that the maximal rate of transport (Vmax) was reduced for both substrates (Fig. 7). The apparent substrate affinity (Km) for estradiol-17β-glucuronide was unchanged, causing inhibition of transport at all concentrations studied. In contrast, the affinity for estrone-3-sulfate was increased 10-fold, leading to stimulation of transport at low substrate concentrations and inhibition of transport at high substrate concentrations.
There are many mechanisms by which transporter function may be inhibited or stimulated. Changes in the expression of transporter protein on the cell surface, whether through transcriptional regulation or post-translational modifications, would affect uptake of all substrates in the same manner, and thus cannot explain the substrate-specific changes in transporter activity produced by ursolic acid, oleanolic acid, and compound 6. Compound 4 inhibited transport of both substrates by OATP1B1 but not OATP1B3, which could be explained by rapid internalization of OATP1B1. However, the full inhibitory effect described here took place during 20 s of exposure, whereas phosphorylation-induced internalization of OATP2B1 and rat Oatp1a1 proteins seems to take at least 10 to 30 min (Köck et al., 2010; Choi et al., 2011). However, post-translational modifications such as phosphorylation may also directly alter activity of a transporter, as is seen with the glucose transporter (Berridge and Tan, 1995), and theoretically could do so in a substrate-dependent way. Alternatively, if the isolated compounds are substrates of OATP1B1 or OATP1B3, they could exert their effects through competitive inhibition. There is considerable evidence that OATPs have multiple binding sites; therefore a compound may inhibit uptake of a substrate that shares its binding site while having no effect on a substrate that has a distinct binding site. This competitive inhibition would be expected to decrease affinity for the substrate that shares a binding site, while not affecting the Vmax. Unfortunately, as relatively weak inhibitors, ursolic acid, oleanolic acid, and compound 4 did not have statistically significant effects on the kinetic parameters of OATP1B1- or OATP1B3-mediated uptake, suggesting that if they are substrates, they have very low affinity. A final explanation for these results is that the compounds may bind to a portion of OATP1B1 or OATP1B3, sterically hindering either the binding or the translocation of substrates. In the case of R002, this binding could also cause a conformational change in OATP1B3 that increases the affinity of the transporter for estrone-3-sulfate while not affecting the estradiol-17β-glucuronide binding site.
It is noteworthy that although compound 6 stimulated transport of estrone-3-sulfate, the structurally similar rutin (compound 5) did not show this effect. However, rutin was previously shown to stimulate uptake of 0.5 μM dehydroepiandrosterone sulfate by OATP1B1 (Wang et al., 2005). This demonstrates that stimulation of uptake is not a phenomenon specific to OATP1B3, but may be shared by all OATPs. This is further supported by a previous study on OATP2B1 where different steroids stimulated the uptake of either estrone-3-sulfate or dehydroepiandrosterone sulfate by OATP2B1 (Grube et al., 2006). Substrate-dependent effects of OATP modulators as shown in this study have also been seen previously. We found that epigallocatechin gallate inhibits OATP1B3-mediated uptake of Fluo-3, has no effect on uptake of estradiol-17β-glucuronide, and stimulates uptake of estrone-3-sulfate (Roth et al., 2011). We also discovered that clotrimazole stimulates OATP1B3-mediated transport of estradiol-17β-glucuronide, inhibits transport of Fluo-3, and has little effect on uptake of estrone-3-sulfate (Gui et al., 2008). Gemfibrozil inhibits OATP1B1 uptake of pravastatin, fluvastatin, simvastatin, and taurocholate, but not estrone-3-sulfate or troglitazone sulfate (Noé et al., 2007). Low concentrations of rosiglitazone (10 μM) inhibit OATP1B1 and OATP1B3 transport of bromosulfophthalein, but stimulate uptake of pravastatin by both transporters (Bachmakov et al., 2008).
Inhibition and stimulation of OATPs can clearly depend on the substrate being transported. Although there are compounds that can nonspecifically inhibit transport of all OATP substrates, as rifampicin seems to do, there are no compounds that selectively inhibit transport of all substrates by a single OATP. Using a general OATP inhibitor to reduce the hepatic first-pass effect of a drug is likely to cause a wide range of effects, because of inhibition of transport of other OATP substrates by OATPs expressed throughout the body. Therefore, to be used therapeutically, it is essential to identify compounds that selectively inhibit either OATP1B1 or OATP1B3 while having no effect on transport mediated by other OATPs. Ideally, the inhibition would also be less potent for endogenous substrates or other drugs such as statins than for the drug with the disadvantageous first-pass effect. Likewise, to increase uptake of drugs by hepatocytes or OATP-expressing cancer cells, stimulation of OATP transport should be restricted to the OATP being targeted, while having little effect on uptake of other substrates by that transporter. It is therefore essential to identify selective OATP modulators.
In conclusion, we have identified four compounds that modulate OATP function. Ursolic acid, oleanolic acid, and 8-trans-p-coumaroyloxy-α-terpineol inhibit estradiol-17β-glucuronide uptake by OATP1B1 but not by OATP1B3, whereas ursolic acid and oleanolic acid inhibit estrone-3-sulfate uptake by both transporters. Compound 6 inhibits transport by OATP1B1, but has substrate-dependent effects on OATP1B3, noncompetitively inhibiting uptake of both substrates at high substrate concentrations, but stimulating estrone-3-sulfate uptake at low substrate concentrations by increasing affinity. The results of this study show that plant materials are a good starting point for the isolation of OATP-modulating compounds, and a bioassay-guided approach can be used to efficiently identify selective OATP modulators.
Authorship Contributions
Participated in research design: Roth, Araya, Timmermann, and Hagenbuch.
Conducted experiments: Roth and Araya.
Performed data analysis: Roth, Araya, Timmermann, and Hagenbuch.
Wrote or contributed to the writing of the manuscript: Roth, Araya, Timmermann, and Hagenbuch.
Footnotes
This work was supported by the National Institutes of Health National Institute of General Medical Sciences [Grant R01GM077336]; and the National Institutes of Health National Center for Research Resources [Grant P20RR021940].
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
doi:10.1124/jpet.111.184564.
-
ABBREVIATIONS:
- OATP
- organic anion-transporting polypeptide
- HPLC
- high-performance liquid chromatography
- DMSO
- dimethyl sulfoxide
- E3S
- estrone-3-sulfate
- E17β
- estradiol-17β-glucuronide
- compound 4
- 8-trans-p-coumaroyloxy-α-terpineol
- compound 6
- quercetin 3-Ο-α-l-arabinopyranosyl (1→2) α-l-rhamnopyranoside
- MeOH
- methanol
- HEX
- hexane
- BUOH
- butanol
- CC
- column chromatography
- Si-Gel
- silica gel
- EtOAc
- hexanes-ethyl acetate
- Frac
- fraction
- GF120918
- N-(4-[2-(1,2,3,4-tetrahydro-6,7-dimethoxy-2-isoquinolinyl)ethyl]-phenyl)-9,10-dihydro-5-methoxy-9-oxo-4-acridine carboxamide
- SN-38
- 7-ethyl-10-hydroxy-camptothecin
- YM758
- (−)-N-{2-[(R)-3-(6,7-dimethoxy-1,2,3,4-tetrahydroisoquinoline-2-carbonyl)piperidino]ethyl}-4-fluorobenzamide
- BAPA
- bile acid conjugated N-(3-aminopropyl)-1,3-propanediamine.
- Received May 29, 2011.
- Accepted August 15, 2011.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}