


Visual Overview

Abstract
Glyburide is frequently used to treat gestational diabetes owing to its low fetal accumulation resulting from placental efflux by the breast cancer resistance protein (BCRP)/ABCG2 transporter. Here we sought to determine how exposure to the dietary phytoestrogen genistein and expression of a loss-of-function polymorphism in the ABCG2 gene (C421A) impacted the transport of glyburide by BCRP using stably transfected human embryonic kidney 293 (HEK) cells, human placental choriocarcinoma BeWo cells, and human placental explants. Genistein competitively inhibited the BCRP-mediated transport of 3H-glyburide in both wild-type (WT) and C421A-BCRP HEK-expressing cells, with greater accumulation of 3H-glyburide in cells expressing the C421A variant. In BeWo cells, exposure to genistein for 60 minutes increased the accumulation of 3H-glyburide 30%–70% at concentrations relevant to dietary exposure (IC50 ∼180 nM). Continuous exposure of BeWo cells to genistein for 48 hours reduced the expression of BCRP mRNA and protein by up to 40%, which impaired BCRP transport activity. Pharmacologic antagonism of the estrogen receptor attenuated the genistein-mediated downregulation of BCRP expression, suggesting that phytoestrogens may reduce BCRP levels through this hormone receptor pathway in BeWo cells. Interestingly, genistein treatment for 48 hours did not alter BCRP protein expression in explants dissected from healthy term placentas. These data suggest that whereas genistein can act as a competitive inhibitor of BCRP-mediated transport, its ability to downregulate placental BCRP expression may only occur in choriocarcinoma cells. Overall, this research provides important mechanistic data regarding how the environment (dietary genistein) and a frequent genetic variant (ABCG2, C421A) may alter the maternal-fetal disposition of glyburide.
Introduction
Gestational diabetes (GD) is on the rise worldwide (Dabelea et al., 2005; Ferrara, 2007) and affects 5%–10% of pregnant women in the United States (DeSisto et al., 2014). Glyburide (INN: glibenclamide) is a second-generation sulfonylurea drug used in the treatment of type 2 diabetes and, more recently, GD. In 2000, Langer and colleagues performed a prospective randomized clinical trial that demonstrated the efficacy and safety of glyburide in the treatment of GD compared with insulin (Langer et al., 2000). Furthermore, glyburide was undetectable in cord serum (limit of detection <10 ng/ml), suggesting limited fetal exposure. These findings launched additional investigations (Bertini et al., 2005; Jacobson et al., 2005; Anjalakshi et al., 2007; Ogunyemi et al., 2007) and ultimately resulted in a major shift in the management of GD. By 2011, more than 50% of U.S. obstetricians were prescribing glyburide as first-line pharmacotherapy for GD (Camelo Castillo et al., 2014).
Kraemer et al. (2006) were the first to demonstrate that glyburide is actively removed from the fetal to the maternal circulation, suggesting that transport plays a major role in the transplacental disposition of glyburide (Kraemer et al., 2006). Multiple transporters in the placenta interact with glyburide (Gedeon et al., 2006, 2008a, 2008b; Hemauer et al., 2010), significant evidence points to the breast cancer resistance protein (BCRP/ABCG2) as the major transporter responsible for the active extrusion of glyburide from the placenta. This has been demonstrated using BCRP-overexpressing cells, Bcrp knockout mice, pregnant rats, human placentas, and human placental vesicles (Pollex et al., 2008, 2010; Zhou et al., 2008; Feinshtein et al., 2013).
BCRP is a transmembrane protein that moves xenobiotics and endogenous chemicals out of cells. In the intestine, liver, and kidney, BCRP is expressed on the apical membrane of epithelial cells and promotes the excretion of drugs and chemicals from the body (Maliepaard et al., 2001). In the placenta, BCRP is localized primarily to the apical membrane of syncytiotrophoblasts, where it transports substrates back to the maternal circulation such that fetal exposure to various chemicals including glyburide is restricted (Maliepaard et al., 2001). Although placental BCRP plays a critical role in protecting the fetus from exposure to glyburide, there is limited understanding of the potential consequences of reduced placental BCRP function in pregnant women prescribed glyburide for GD. Infants born to this population of women may be at an elevated risk of neonatal hypoglycemia as a result of fetal glyburide exposure, potentially leading to low birth weight, increased morbidity, and impaired neurologic development (reviewed in Williams, 1997). BCRP function may be compromised by genetic single nucleotide polymorphisms, as well as by environmental factors, including dietary constituents.
A single nucleotide polymorphism occurring at nucleotide 421 (C > A) in the ABCG2 gene leads to an amino acid change from glutamine to lysine (Q141K). In vitro, the C421A genotype is associated with reduced BCRP function (Kondo et al., 2004; Pollex et al., 2010). Importantly, the C421A genotype occurs frequently in Asian (C/A: 30%, A/A: 10%) and Caucasian (C/A: 15%, A/A: 1%) populations (Imai et al., 2002; Zamber et al., 2003; Kobayashi et al., 2005).
Genistein is a soy isoflavone that is found naturally in plants of the Leguminosae family and occurs abundantly in soybeans (U.S. Department of Agriculture, http://www.ars.usda.gov/nutrientdata/isoflav;
United Soybean Board, 2014, http://www.soyconnection.com/sites/default/files/ConsumerAttitudes_Med_062714.pdf).
Considering the increasing consumption of soy-containing products (i.e., tofu, soy formula, dietary supplements) over the last 10 years (United Soybean Board, 2014, http://www.soyconnection.com/sites/default/files/ConsumerAttitudes_Med_062714.pdf), it is important to recognize that components of soy, including genistein, may interfere with the disposition of prescribed drugs. Genistein alone and in combination with other isoflavones has been shown to directly interfere with the BCRP-mediated disposition of drugs in vivo, including BCRP substrates enrofloxacin and nitrofurantoin (Pulido et al., 2006; Merino et al., 2010). Genistein also interacts with proteins that may regulate placental BCRP expression, including the estrogen receptor and the epidermal growth factor receptor (Kuiper et al., 1998; Traxler et al., 1999). Considering this evidence, genistein may reduce placental BCRP function in two distinct manners by: 1) direct inhibition of BCRP activity and 2) altered transcriptional regulation of BCRP.
Because of the increased use of glyburide in GD treatment and the sensitivity of the developing fetus, it is critical to consider separately and together the influence of genetic and environmental factors on the placental BCRP-mediated disposition of glyburide. The purpose of this study was to use complementary in vitro and ex vivo model systems to characterize the molecular mechanisms by which dietary-relevant concentrations of genistein impair the transport of glyburide by BCRP in placental trophoblasts.
Materials and Methods
Chemicals.
Unless otherwise specified, all chemicals were obtained from Sigma-Aldrich (St. Louis, MO).
Cell Culture.
Human embryonic kidney-293 (HEK) cells were stably transfected with an empty vector (EV, pcDNA 3.1), the human wild-type (WT) BCRP/ABCG2 gene, or the human C421A BCRP/ABCG2 variant (kindly provided by Dr. Robert Robey, National Cancer Institute, National Institutes of Health, Bethesda, MD) (Morisaki et al., 2005). Cells were maintained in Dulbecco’s modified Eagle medium (Life Technologies, Carlsbad, CA) with 10% fetal bovine serum (Atlanta Biologicals, Norcross, GA), 1% penicillin-streptomycin (Life Technologies), and 0.2 mg/ml geneticin (Life Technologies) to select for transfected cells. Human placental choriocarcinoma BeWo cells were purchased from American Type Culture Collection (ATCC, Manassas, VA) and grown in phenol-red free Dulbecco’s modified Eagle’s medium: F-12 (Life Technologies) with 10% charcoal-stripped and dextran-treated fetal bovine serum (Atlanta Biologicals) and 1% penicillin-streptomycin (Life Technologies). All cells were maintained at 37°C with 5% CO2 and used in experiments at 80%–90% confluence. For the regulation studies, BeWo cells were incubated with genistein (10 μM) or ICI 182,780 (0–1 μM) for 48 hours, after which they were processed for mRNA, protein, or functional analysis. All chemicals were dissolved in dimethyl sulfoxide (DMSO) such that the final percentage (% v/v) of DMSO did not exceed 0.1% of the treatment media.
Flow Cytometry.
HEK cells overexpressing the EV, WT-BCRP, or C421A-BCRP gene were suspended in cold isotonic PBS with 0.5% bovine serum albumin (BSA), centrifuged (500g, 5 minutes), and washed three times in PBS with 0.5% BSA. Cells in suspension were blocked with human IgG (2 μg/500,000 cells in PBS/0.5% BSA) for 15 minutes at room temperature. Cells were then incubated for 45 minutes at 4°C with the monoclonal phycoerthyrin-labeled anti-BCRP antibody (5D3) or the phycoerthyrin-labeled negative control IgG antibody (R&D Systems, Minneapolis, MN) according to the manufacturer’s protocol. Cells were washed three times (5 minutes with PBS/0.5% BSA) and resuspended in 2% paraformaldehyde/PBS for flow cytometry analysis using a Gallios/FC500 Cytometer with 488-nm wavelength laser excitation (Beckman Coulter, Indianapolis, IN) in the Flow Cytometry/Cell Sorting Core Facility at Rutgers University.
Hoechst 33342 Accumulation Assay.
The BCRP-specific fluorescent substrate, Hoechst 33342, was used to quantify BCRP function in HEK and BeWo cells as previously described (Bircsak et al., 2013). Briefly, HEK and BeWo cells were trypsinized and added to a 96-well plate. After centrifugation (500g, 5 minutes, 5°C) and removal of the media, cells were loaded with Hoechst 33342 (7–15 µM) in the presence or absence of the BCRP-specific inhibitor, Ko143 (1–3000 nM), or the test compound, genistein (0.1–100 µM, for 30 minutes at 37°C and 5% CO2, uptake phase). Cells were washed, centrifuged, and resuspended in substrate-free media with or without inhibitor for 1 hour (efflux phase). During all phases, inhibitor compounds were initially dissolved in DMSO and did not exceed 1% (v/v) of the final treatment media. After the efflux phase, cells were centrifuged, washed, and resuspended in cold PBS for quantification of intracellular fluorescence using the Cellometer Vision automated cell counter (Nexcelom Bioscience, Lawrence, MA). Cell suspension (20 µl) was added to the cell counting chamber, and each sample was analyzed using bright-field images for cell size and cell number. A VB-450-302 filter (excitation/emission: 375/450) allowed for intracellular fluorescence detection of Hoechst 33342. The total number of cells analyzed for each sample ranged from 200 to 2000. In BeWo cells, raw fluorescence intensity for each cell was normalized to cell size.
3H-Glyburide Accumulation Assay.
HEK and BeWo cells were trypsinized and added to a 96-well plate. After centrifugation (500g, 5 minutes, 5°C) and removal of the media, cells were loaded with 3H-glyburide (0.1–10 µM; Specific Activity: 40 µCi; PerkinElmer, Waltham, MA) in the presence and absence of the BCRP-specific inhibitor, Ko143 (1–3000 nM) or the test compound, genistein (0.01–20 µM) for 1 hour at 37°C and 5% CO2. Inhibitors were dissolved in DMSO such that the final concentration of DMSO did not exceed 1% (v/v) in the final treatment media. After the 1-hour incubation, cells were centrifuged, washed, and lysed using 1 M NaOH. Lysates were then neutralized using 1 M HCl and added to 4 ml ScintiSafe Econo 1 liquid scintillation fluid (Fisher Scientific, Waltham, MA) in 7-ml glass liquid scintillation vials (Perkin Elmer). Radioactivity was detected using a TriCarb 2100TR Liquid Scintillation Analyzer (PerkinElmer-Packard, Waltham, MA). A standard curve was used to extrapolate glyburide concentrations. Accumulation of glyburide was normalized to the total protein concentration of cell lysates from representative wells using the bicinchoninic acid (BCA) assay (Pierce Biotechnology, Rockford, IL).
Western Blot.
HEK and BeWo cells were lysed in lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 5 mM EDTA, 1% Triton X-10, and 1% protease inhibitor cocktail). Placenta membrane fractions were prepared as previously described (Memon et al., 2014). Protein concentrations were determined by the BCA assay, and 5–10 μg of total protein was added to SDS-PAGE 4%–12% Bis-Tris gels (Life Technologies), which were resolved by electrophoresis. Transfer of proteins onto polyvinylidene fluoride membranes was completed using an overnight transfer apparatus (BioRad Criterion Blotter; BioRad, Hercules, CA). Membranes were blocked using 5% nonfat milk in 0.5% Tween-20–PBS and then incubated with primary antibodies in 2% nonfat milk in 0.5% Tween-20-PBS for 2 hours. Primary antibodies detected proteins, including BCRP (BXP-53, 1:5000; Enzo Life Sciences, Farmingdale, NY), β-actin (Ab8227, 1:2000, Abcam, Cambridge, MA), and Na+/K+ ATPase (Ab76020, 1:20,000, Abcam). The membranes were washed and species-specific HRP-conjugated secondary antibodies were added to the blots for 1 hour. Supersignal West Dura Extended Duration Substrate (Pierce Biotechnology) was used for chemiluminescent detection of proteins with a Fluorchem Imager (ProteinSimple, Santa Clara, CA). Semiquantitation of bands was performed using the AlphaView Software (ProteinSimple). β-Actin or Na+/K+ ATPase were used as loading controls where appropriate.
RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction.
BeWo cells were collected in buffer RLT provided in the RNeasy Mini Kit (Qiagen, Germantown, MD), plus 1% β-mercaptoethanol. Cells were lysed using QIAshredder columns (Qiagen), and total RNA was isolated with the RNeasy Mini Kit (Qiagen) according to the manufacturer’s protocol. Total RNA concentration and purity (260/280) were determined using a Nanodrop spectrophotometer (Fisher Scientific). Complementary DNA (cDNA) was generated from total RNA (500 ng) using the High-Capacity cDNA Reverse Transcription Kit (Life Technologies) and a MultiGene OptiMax Thermal Cycler (Labnet International Inc., Edison, NJ). Quantitative PCR was performed with specific forward and reverse primers (Integrated DNA Technologies, Inc., Coralville, IA), cDNA, Sybr Green dye (Life Technologies), and a ViiA7 RT-PCR System (Life Technologies) in the Bionomics Research and Technology Center at Rutgers University. Ct values were converted to delta delta Ct values by comparison with ribosomal protein 13A (RPL13A) as a reference gene and the DMSO-treated control cells. Primer sequences for BCRP and RPL13A are provided in (Supplemental Table 1).
Patient Selection.
Written informed consent was obtained and placentas were collected from five healthy women with uncomplicated pregnancies after term delivery by scheduled cesarean section. Inclusion criteria were healthy women, aged 18–40, term gestation (≥36 weeks), and scheduled cesarean section without labor. Exclusion criteria included chronic medical conditions (e.g., hypertension, diabetes, autoimmune disorders); pregnancy-induced medical conditions (e.g., pregnancy-induced hypertension, pre-eclampsia, gestational diabetes); maternal infection; clinical chorioamnionitis; medication use (with the exception of prenatal vitamins); maternal smoking, alcohol, or drug abuse; and known fetal chromosomal abnormalities. All placentas expressed two wild-type alleles (C/C) at the 421 nucleotide position in the ABCG2 gene as determined by a Fluidigm Biomark Genetic Analysis system in the Bionomics Research and Technology Center at Rutgers University. Other patient demographic information is provided in (Supplemental Table 2). The Institutional Review Boards of Robert Wood Johnson Medical School (Protocol 0220100258) and Rutgers University (Protocol E12-024) approved this study. Whereas the range of maternal ages in this study was 34–39 years, no effect of maternal age on placental BCRP expression has been documented.
Placental Explants.
All placentas were obtained within 10 minutes of delivery and processed within 2 hours. Placentas were carefully inspected for any visible abnormalities and location of umbilical cord. Only placentas with central or eccentric cord insertion were used. The maternal decidua and the chorionic plate along the overlying membranes were removed, and sections of villous tissue were washed in PBS three times to remove maternal blood before dissection into 2 × 2 × 2-mm (8 mm3) pieces of tissue. In a 24-well dish, two pieces of 8-mm3 villous tissue were cultured in DMEM:F12 media without phenol red (Life Technologies) and with 10% charcoal-stripped and dextran-treated fetal bovine serum (Atlanta Biologicals) and 1% penicillin-streptomycin (Life Technologies). To allow for complete degeneration and regeneration of the syncytium as described by other laboratories (Siman et al., 2001), explants were maintained in culture for 5 days, with the media changed and collected every 24 hours. On day 5, genistein (0–10 μM) treatment began for 48 hours (media not changed during 48 hours of treatment). Genistein was dissolved in DMSO, and the final DMSO percentage was 0.1% (v/v) in the treatment media. Following the 48-hour treatment, explants were washed in PBS and frozen in liquid nitrogen before homogenization in sucrose (250 mM)-Tris (10 mM) buffer (pH 7.4) with protease inhibitors (1%, v/v), using the TissueLyser LT (Qiagen) according to the manufacturer’s protocol.
hCG ELISA and LDH Assay.
Each day of explant culture, media were collected for assessment of syncytiotrophoblast function and overall tissue integrity by quantification of human chorionic gonadotropin (hCG) and lactate dehydrogenase (LDH), respectively. The hCG ELISA was completed using the manufacturer’s protocol (Calbiotech, Spring Valley, CA). Activity of LDH in the media was determined using the manufacturer’s protocol.
Statistical Analysis.
Data are presented as mean ± S.D. of multiple independent experiments (n = 3–5) and analyzed using Graphpad Prism 5.0 (Graphpad Software Inc., La Jolla, CA). Two-way analysis of variance with Bonferroni post-test, One-way analysis of variance with Newman-Keuls post-test, or a two-tailed student’s t test were used to assess statistical significance (P < 0.05) according to the number of comparisons and variables. Nonlinear regression analysis (dose-response: [log] inhibitor versus response—variable slope (four parameters) or Michaelis-Menten) was used to determine kinetic parameters (IC50, Km, Jmax).
Results
Functional Inhibition of BCRP by Genistein
WT- and C421A-BCRP Overexpressing HEK Cells.
Whole-cell and cell-surface protein expression of BCRP were assessed by Western blot and flow cytometry, respectively. Three independent experiments revealed that BCRP protein expression in whole-cell lysates was reduced by about 30% in C421A-BCRP expressing cells compared with WT-BCRP expressing cells (P = 0.172) (Fig. 1A). At the cell surface, BCRP protein expression in the C421A-BCRP cells was significantly decreased by 50% compared with the WT cells (Fig. 1B). Both methods confirmed the absence of BCRP protein in HEK cells expressing the EV (data not shown).

Characterization of BCRP protein in WT- and C421A-BCRP–overexpressing HEK cells. (A) BCRP protein expression in HEK whole-cell lysates was determined by Western blot (10 µg protein homogenate/lane). Na+/K+ ATPase was used as a loading control. Western blot data are presented as a representative Western blot from one experiment. The bar graph shows the semiquantitation of band densities and represents the mean ± S.D. from three independent experiments. (B) BCRP protein expression on the cell surface of HEK cells was determined by flow cytometry. Cells were incubated for 45 minutes with the phycoerthyrin-labeled anti-BCRP antibody (5D3) or the phycoerthyrin-labeled negative control IgG antibody (green: WT-BCRP 5D3 stained cells, blue: C421A-BCRP 5D3 stained cells, red: WT-BCRP IgG control). Flow cytometry histogram data are presented as one representative experiment. Bar graph represents mean ± S.D. fluorescence intensity of individual cells from three independent experiments. †Statistically significant differences (P < 0.05) compared with WT-BCRP.
Two substrate accumulation assays (Hoechst 33342 and 3H-glyburide) were used to examine modulation of BCRP transport activity by pharmacologic inhibition and genetic variation. Initial experiments determined the optimal substrate concentrations for use in the two accumulation assays [Hoechst 33342, 7 µM and 10 µM unlabeled glyburide, and 0.1 µM 3H-glyburide; (Supplemental Fig. 1)]. In the absence of inhibitor, there was a trend for the cells overexpressing the variant BCRP (C421A) to accumulate 50%–100% more Hoechst 33342 or 3H-glyburide than the respective WT-BCRP cells (Fig. 2). The BCRP-specific inhibitor Ko143 increased the accumulation of Hoechst 33342 and 3H-glyburide in a concentration-dependent manner in both BCRP genotypes (Fig. 2, A and C), demonstrating the ability of the assays to detect inhibition of BCRP activity. Similar to Ko143, genistein increased the accumulation of Hoechst 33342 and 3H-glyburide in WT- and C421A-BCRP expressing cells (Fig. 2, B and D). Confirming the involvement of the BCRP transporter in the genistein-induced substrate accumulation, EV cells lacking BCRP protein accumulated both substrates, which was not altered by genistein (data not shown). It should be noted that Ko143 significantly increased the accumulation of both BCRP substrates in the C421A-BCRP cells at lower concentrations than in WT-BCRP cells. Further, the half-maximal inhibitory concentration (IC50) value for the inhibition of 3H-glyburide transport in the C421A-BCRP (10.1 ± 3.11 nM) cells by Ko143 was 50% lower than the WT-BCRP cells (21.5 ± 1.81 nM) (Table 1). For genistein, the IC50 values for the inhibition of 3H-glyburide transport in both BCRP genotypes were comparable (WT: 4.65 ± 0.27, C421A: 3.92 ± 1.26) (Table 1).

Inhibition of BCRP transport in WT- and C421A-BCRP overexpressing HEK cells. BCRP function was assessed by measuring the cellular accumulation of (A, B) Hoechst 33342 (7 µM) or (C, D) 3H-glyburide (10 µM unlabeled glyburide, 0.1 μM 3H-glyburide) in the presence of increasing concentrations of the BCRP-specific inhibitor, Ko143 or genistein. Intracellular fluorescence and radioactivity were quantified by a Cellometer Vision or a liquid scintillation counter, respectively. Bar graphs represent mean ± S.D. (n = 3 independent experiments). *Statistically significant differences (P < 0.05) compared with 0 µM genotype control. †Statistically significant differences (P < 0.05) compared with WT-BCRP control.
Inhibition of 3H-glyburide transport in BCRP-overexpressing cells and placental BeWo cells
To determine the mechanism by which genistein inhibited the BCRP-mediated transport of glyburide, accumulation experiments were performed using varying concentrations of both glyburide and genistein. Genistein significantly decreased the Km value for glyburide transport by 80% without changing the Jmax in either BCRP genotype (Fig. 3; Table 2), suggesting that genistein competitively inhibited the BCRP-mediated efflux of glyburide. In addition, the mean rate of 3H-glyburide accumulation in the genistein-treated cells (2 and 10 μM) was significantly greater than the mean rate of 3H-glyburide accumulation in the control cells in both genotypes (0 μM genistein) (Fig. 3).

Characterization of the genistein-glyburide interaction mediated by BCRP. BCRP function was assessed in WT- and C421A-BCRP HEK-expressing cells by measuring the cellular accumulation of 3H-glyburide (0–500 μM unlabeled glyburide, 0.1 μM 3H-glyburide) in the presence of increasing concentrations of genistein (0–10 µM) and was quantified using a liquid scintillation counter. Data represent mean ± S.D. (n = 3 independent experiments). Nonlinear regression analysis (Michaelis-Menten) was used for curve-fitting analysis. *Statistically significant differences (P < 0.05) in the mean rate of 3H-glyburide accumulation compared with the 0 µM control.
Kinetic parameters of glyburide transport in HEK cells overexpressing BCRP protein (WT or C421A)a
Placental BeWo Cells.
Expression of endogenous BCRP protein in placental BeWo cells was confirmed by Western blot along with BCRP protein expression in plasma membranes isolated from three individual placentas (Fig. 4A). BCRP function in BeWo cells was assessed using two substrate accumulation assays (Hoechst 33342 and 3H-glyburide) (Supplemental Fig. 2). Ko143 and genistein significantly increased the accumulation of both Hoechst 33342 and 3H-glyburide by 30%–100% in placental BeWo cells (Fig. 4, B–E). Notably, concentrations as low as 0.01 µM genistein inhibited 3H-glyburide transport (IC50 = 0.18 ± 0.22 μM; Table 1). Prototypical inhibitors of other efflux transporters (MDR1: PSC833: 2 µM; MRPs: 25 µM MK571) did not alter the accumulation of 3H-glyburide in the BeWo cells (data not shown), supporting the specific involvement of BCRP in the inhibition of glyburide transport by genistein.
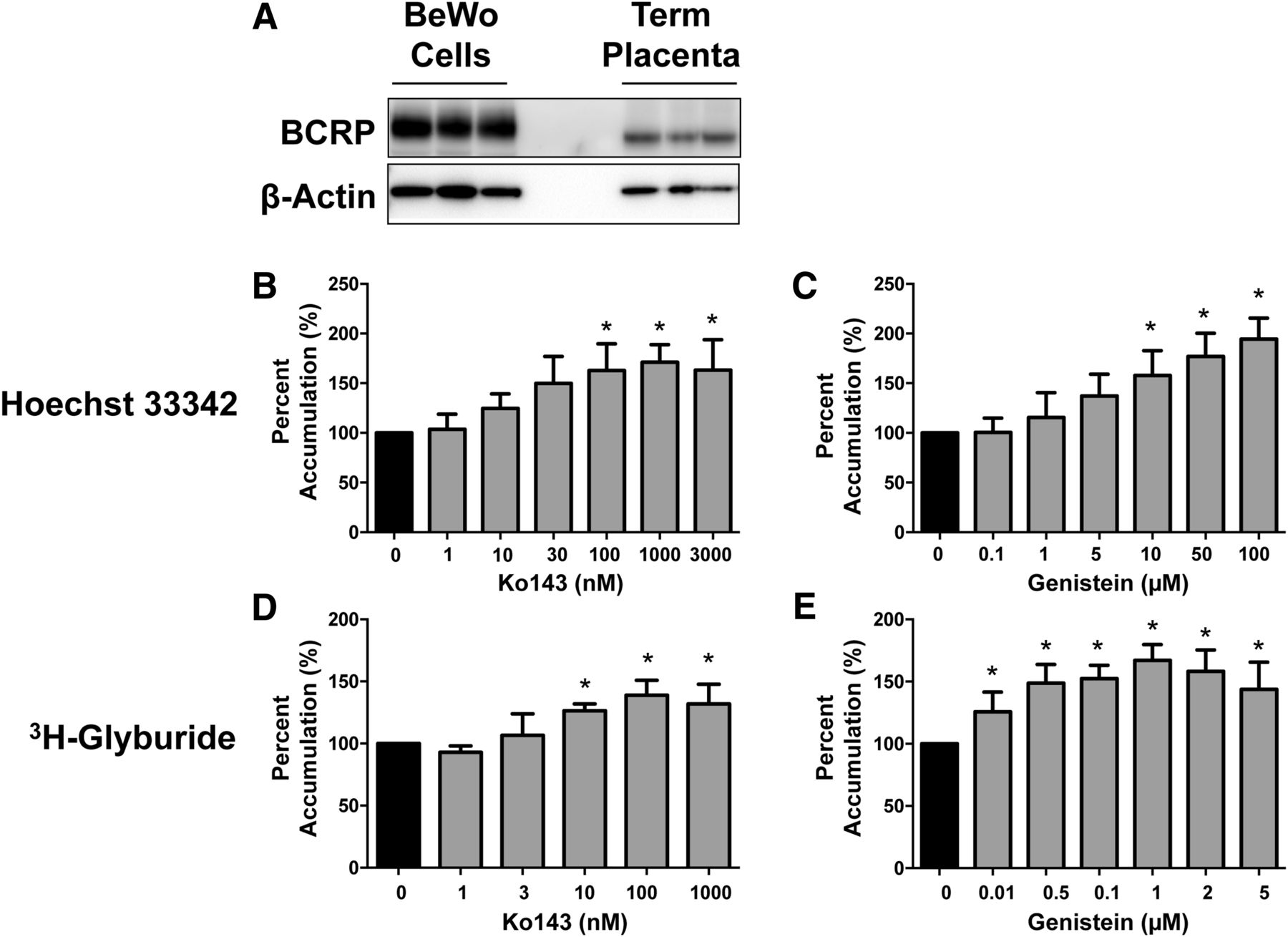
Inhibition of BCRP transport in placental BeWo cells. (A) BCRP protein expression in placental BeWo whole-cell lysates compared with human placenta plasma membrane fractions was determined by Western blot (9 µg of protein homogenate/lane). β-Actin was used as a loading control. (B–E) BCRP function was assessed in the presence of increasing concentrations of Ko143 (B, D) or genistein (C, E), by the accumulation of (B and C) Hoechst 33342 (15 µM), or (D, E) 3H-glyburide (0.1 μM). Intracellular fluorescence or radioactivity was quantified using a Nexcelom Cellometer Vision or a liquid scintillation counter, respectively. Bar graphs represent mean ± S.D. (n = 3 or 4 independent experiments). *Statistically significant differences (P < 0.05) compared with the 0 µM control.
Regulation of Placental BCRP Expression by Genistein
BeWo Cells.
To determine whether genistein could alter the transcriptional regulation of BCRP expression, placental BeWo cells were incubated with genistein for 48 hours. Compared with control cells, genistein significantly decreased BCRP mRNA and protein expression up to 40% (Fig. 5, A and B). Furthermore, 48-hour exposure to genistein (5 μM) increased the accumulation of 3H-glyburide by 30% in placental BeWo cells, which was comparable to the pharmacologic inhibition of glyburide transport by Ko143 (100 nM) (Fig. 5C). Notably, genistein did not affect mRNA and protein expression of other efflux transporters, including the multidrug resistance-associated protein 1 (MRP1) (data not shown). Additionally, no difference was found in cell morphology or the syncytialization marker, syncytin mRNA expression, suggesting that genistein did not alter BeWo cell differentiation (data not shown). Because genistein is a phytoestrogen, the mechanism by which it downregulated BCRP mRNA and protein expression in BeWo cells was investigated using the estrogen receptor α antagonist ICI 182,780. Exposure to ICI 182,780 (1 µM) for 48 hours did not alter BCRP protein expression; however, the combination of genistein and ICI 182,780 together abolished the downregulation of BCRP protein expression caused by genistein alone (Fig. 6).
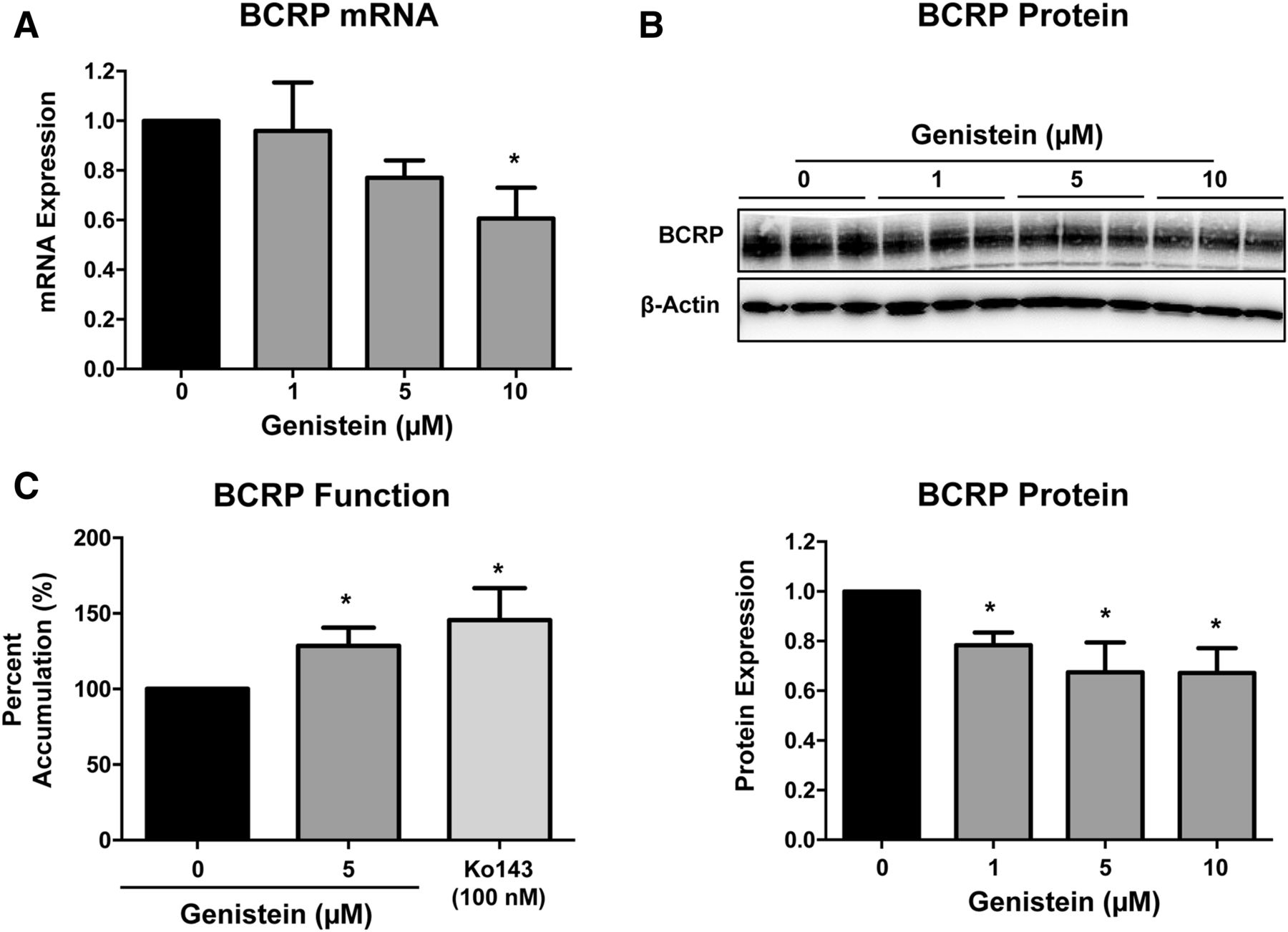
Regulation of BCRP transporter expression and function in placental BeWo cells. After a 48-hour exposure to genistein (0–10 µM), (A) qPCR was used to quantify BCRP and housekeeping gene, RPL13A, mRNA expression in placental BeWo cells. (B) BCRP protein expression in placental BeWo whole-cell lysates was determined by Western blot (10 µg of protein homogenate/lane). β-Actin was used as a loading control. Western blot data are presented as a representative Western blot from one experiment. (C) BCRP function was assessed by the cellular accumulation of 3H-glyburide (0.1 μM), which was quantified using a liquid scintillation counter. All bar graphs represent mean ± S.D. (n = 3 or 4 independent experiments). *Statistically significant differences (P < 0.05) compared with the 0 µM genistein control.

Estrogen receptor-mediated regulation of the BCRP transporter in placental BeWo cells. BCRP protein expression in placental BeWo whole-cell lysates was determined by Western blot (10 µg of protein homogenate/lane) after a 48-hour exposure to the estrogen receptor antagonist, ICI 182,780 in the presence and absence of genistein. β-Actin was used as a loading control. All Western blot data are presented as a representative Western blot from one experiment. The bar graphs are the semiquantitation of band density and represent the mean ± S.D. from three independent experiments. *Statistically significant differences (P < 0.05) compared with the 0 µM control.
Placental Explants.
Additional experiments aimed to determine whether genistein-mediated downregulation of BCRP expression could occur in native placental tissue. Explants were obtained from healthy human term placentas and allowed to undergo shedding and syncytialization for 5 days before treatment with genistein on days 5–7. Viability of the explants was confirmed by a decrease in LDH activity in the media over seven days (Supplemental Fig. 3A; Siman et al., 2001). The degree of syncytialization was verified by detection of hCG in the media and by histologic analysis (Supplemental Fig. 3B and 3C) (Siman et al., 2001)). Genistein treatment did not affect LDH activity or hCG secretion. Furthermore, BCRP protein expression was not altered by genistein (1–10 µM, 48 hours) in human term placental explants (Fig. 7).

Regulation of BCRP transporter expression in human term placental explants. Five healthy human term placentas were collected and processed within 2 hour of delivery. Explants were cultured for 5 days with the media replaced every day, before treatment with genistein (1-10 µM) on day 5 for 48 hour. BCRP protein expression in total explant lysates was determined by western blot (5 µg protein homogenate/lane). β-Actin was used as a loading control. Western blot data are presented as a representative western blot from one placenta. The scatter dot plot is the semiquantitation of band density and represents five individual placentas with mean ± S.D.
Discussion
The present study demonstrated the influence of genetic and dietary factors on the BCRP-mediated transport of glyburide using complementary in vitro and ex vivo placenta model systems. The C421A-BCRP genotype alone reduced the transport of 3H-glyburide compared with WT-BCRP control cells, whereas genistein competitively inhibited 3H-glyburide transport by BCRP to a similar extent in both genotypes. Confirming the potential for a placenta-specific glyburide accumulation, the BCRP-mediated transport of 3H-glyburide was inhibited by a short-term exposure (1 hour) to genistein in placental BeWo cells. Moreover, BCRP mRNA, protein, and function were reduced in BeWo cells exposed to genistein for 48 hours; however, the same genistein exposure did not alter BCRP protein expression in normal placental explant tissues. Results from the present study support future investigations that consider genetic and environmental (dietary genistein) factors when optimizing glyburide treatment in GD patients.
In this study, we confirmed the reduced capability of the C421A-BCRP protein to transport 3H-glyburide in HEK cells that overexpress the mutated BCRP protein (Q141K) compared with WT-BCRP overexpressing cells (Fig. 2; Table 2). Whereas Pollex et al. (2010) described BCRP protein cell-surface expression to be equal between genotypes, we observed less total (30%) and cell-surface (50%, P < 0.05) BCRP protein expression in the C421A-BCRP HEK cells (Fig. 1). Considering these findings, altered BCRP function in our experiments may be due to reduced BCRP protein trafficking to the cell surface, in line with other studies reporting decreased BCRP protein expression and function in C421A-BCRP–overexpressing cells (Imai et al., 2002; Kondo et al., 2004; Tamura et al., 2007, Furukawa et al., 2009; Woodward et al., 2013). In human tissue, the heterozygous variant genotype (421C/A) did not change BCRP protein expression in the intestine or placenta, but there was a significant decrease in BCRP protein expression in placentas of individuals homozygous for the SNP (421A/A) (Zamber et al., 2003; Kobayashi et al., 2005; Urquhart et al., 2008). Taken together, there may be differences in BCRP protein expression in cells or tissue based on the copy number of the C421A gene. Furthermore, there may be a cell-type and/or -tissue–specific effect of C421A on BCRP protein expression. Most importantly, patients expressing one or two variant alleles exhibit increased blood concentrations (sulfasalazine, diflomotecan, rosuvastatin) and enhanced side effects of drugs (gefitinib) that are substrates for BCRP (Sparreboom et al., 2004; Cusatis et al., 2006; Urquhart et al., 2008; Keskitalo et al., 2009). Because of these clinical findings and the frequent occurrence of the SNP, the International Transporter Consortium named the C421A BCRP genetic variant as a clinically relevant transporter polymorphism that should be evaluated in drug development studies (Giacomini et al., 2013).
Over the past 10 years, the sale and consumption of soy in the United States have dramatically increased, likely owing to the growing number of reports suggesting that a soy diet provides health benefits (Strom et al., 1999; Chen et al., 2003; Hussain et al., 2003; Constantinou et al., 2005; Bitto et al., 2008; Clarkson et al., 2011; Squadrito et al., 2013). Taken together with the increase in the prescribing of glyburide for GD (Camelo Castillo et al., 2014), there is a need to investigate the impact of soy on glyburide transport. Genistein and its conjugated metabolites (genistein-sulfate and genistein-glucuronide) are substrates for BCRP, suggesting that they competitively inhibit BCRP transport (Imai et al., 2004; Enokizono et al., 2007; Alvarez et al., 2011; Mease et al., 2012). Others, however, have proposed that genistein can noncompetitively inhibit BCRP transport via disruption of ATP hydrolysis as well (Randak et al., 1999; Di Pietro et al., 2002). To assess the mechanism of genistein inhibition, we used the 3H-glyburide accumulation assay (Pollex et al., 2010) and Michaelis-Menten nonlinear regression analysis to calculate kinetic parameters (Km and Jmax), which represent substrate affinity and rate of accumulation, respectively. It should be noted that because the assay measured 3H-glyburide accumulation, rather than rate of 3H-glyburide efflux, the kinetic parameters trended in the opposite direction of what is normally observed (i.e., decrease in Km indicates reduced affinity of 3H-glyburide for BCRP rather than increased affinity). In cells overexpressing both the WT-BCRP and the C421A-BCRP cells, genistein competitively inhibited the BCRP-mediated transport of 3H-glyburide to similar degrees (Figs. 2D and 3; Table 2). The Km and Jmax values derived from these experiments, however, were calculated for direct comparison with each other and cannot be directly extrapolated to an in vivo situation. Determining Km and Vmax in a study that measures 3H-glyburide efflux rather than accumulation may better correlate to an in vivo glyburide-genistein interaction. Although the various studies have demonstrated genistein to interfere with the transport of other BCRP substrates (i.e., mitoxantrone, enrofloxacin) (Zhang et al., 2004; Pulido et al., 2006), this is the first report implicating genistein in the inhibition of 3H-glyburide transport by BCRP. It is important to note that Ko143 significantly reduced 3H-glyburide transport in the C421A-BCRP overexpressing cells at a lower concentration than the WT-BCRP overexpressing cells, whereas genistein equally inhibited 3H-glyburide transport between cell types (Table 1). This finding suggests that individuals with the C421A-BCRP genotype may be at a greater risk for chemically induced BCRP inhibition depending on the inhibitor compound.
BeWo cells are commonly used as a model of placental transport because they endogenously express functional BCRP protein and produce the placenta-specific hormone, hCG (Takeuchi et al., 1990). In placental BeWo cells, genistein increased the accumulation of Hoechst 33342 and 3H-glyburide (Fig. 4). Interestingly, the concentrations of genistein that inhibited 3H-glyburide transport out of the BeWo cells were quite low and within the range of those observed in the plasma or serum of people consuming a soy diet (0.01–3 µM) (Arai et al., 2000; Uehar et al., 2000; Frankenfeld et al., 2003; Gardner et al., 2009). These data provide plausibility to the assertion that genistein may be able to inhibit BCRP transport of substrates in vivo. The BCRP-specific inhibitor, Ko143, was used as a positive control to confirm the involvement of BCRP in the genistein-mediated cellular accumulation of Hoechst 33342 and 3H-glyburide.
Placental BCRP function may also be affected by chemical-mediated changes in mRNA and/or protein expression. After 48-hour exposure to genistein, BCRP mRNA and protein expression was decreased in BeWo cells compared with the vehicle treated cells (Fig. 5, A and B). In turn, this caused significant accumulation of 3H-glyburide in cells that were exposed to genistein (5 μM, 48 hours) (Fig. 5C). Only one other study has reported a significant decrease in ABCG2 mRNA expression after an in vitro genistein exposure (15 µM, 24 hours, gastric cancer cells, MGC803) (Huang et al., 2014), whereas many others detected no change in BCRP mRNA and/or protein expression in other cell types (Imai et al., 2004; Ebert et al., 2007; Arias et al., 2014).
In the 1960s, genistein was recognized as a phytoestrogen because of its ability to alter reproductive function in sheep (Barrett et al., 1965) and agonize both ERα (IC50= 145 nM) and β (IC50= 8.4 nM) with a greater affinity for ERβ (Martin et al., 1978; Kuiper et al., 1998). This is unlike the endogenous ligand, 17β-estradiol, which agonizes both ERα (IC50= 0.93 nM) and ERβ (IC50= 1.06 nM) with similar affinities (Kuiper et al., 1998). Importantly, an estrogen response element has been identified in the promoter region of the ABCG2 gene (Ee et al., 2004b). Furthermore, 17β-estradiol has been shown to downregulate BCRP protein expression in various cells and tissues, including placental BeWo cells (Imai et al., 2005; Wang et al., 2006; Mahringer and Fricker, 2010), which express both ERα and ERβ (Gambino et al., 2012). It should be noted, however, that other studies have observed an upregulation of BCRP protein by 17β-estradiol in BeWo cells (Ee et al., 2004a; Prouillac et al., 2009) and cytotrophoblasts (Evseenko, et al., 2006). Nonetheless, we aimed to explore the involvement of the ER signaling pathway in the genistein-mediated downregulation of BCRP expression in placental BeWo cells, the ERα antagonist ICI 182,780 (1 μM) (Van Den Bemd et al., 1999; Peekhaus, et al., 2004) alone did not alter BCRP expression but was able to prevent the downregulation of BCRP by genistein (Fig. 6). These findings suggest that ER signaling participates in the repression of BCRP expression by genistein in BeWo cells.
Placental explants are an ex vivo model of the human term placenta as they retain the morphology of the human placental villi, contain multiple cell types, and are frequently used in examining transporter regulation in normal human placenta (Atkinson et al., 2006; Javam et al., 2014). Genistein did not alter BCRP protein expression in human term placental explants (Fig. 7). Although these findings contrast with those observed in placental BeWo cells, it is important to note that extrapolation of gene regulation findings observed in choriocarcinoma cells to normal placenta must be made with caution. DNA methylation patterns vary between normal primary trophoblasts and choriocarcinoma cells, contributing to the overall differential gene expression and regulation profiles between the two types of cells (Novakovic et al., 2011). Additionally, whereas explants are a model of the term placenta, it is important to note that BeWo cells are generally representative of first trimester trophoblasts (Wice et al., 1990), and therefore differences in the regulation of protein expression between the two models is conceivable. Furthermore, explants offer a more complex system than trophoblasts alone. Capillary endothelial cells also express BCRP (Maliepaard et al., 2001) and may confound the results from the explant studies as the Western blot results represent BCRP expression in the whole explant homogenate and not trophoblasts alone. Future research should address the regulation of BCRP expression in explants using immunohistochemistry to localize BCRP expression or by using isolated primary trophoblasts. Together, results from this study indicate that the direct pharmacologic inhibition of BCRP by genistein may have greater implications for BCRP-mediated transport of glyburide than modulation of transcription and/or translation.
The prescription of glyburide for the management of GD has reached an all-time high as most initial reports found no differences in neonatal hypoglycemia rates between insulin- and glyburide-managed GD (Langer et al., 2000; Jacobson et al., 2005; Anjalakshi et al., 2007). More recently, Schartz and colleagues described a weak, but significant, negative correlation between neonatal blood glucose concentration and umbilical cord blood glyburide concentration (Schwartz et al., 2015). Interindividual differences have been found in the umbilical cord blood to maternal blood concentration ratios of glyburide for which the authors named genetic differences in the BCRP/ABCG2 gene as a likely contributor. Differences in diets, including those that are rich in soy, may also contribute to this variability.
In summary, our data demonstrate that genistein inhibits the BCRP-mediated efflux of glyburide in vitro by direct inhibition and reduced protein expression in placental BeWo cells but not placental explants. To better optimize the individualized prescribing of glyburide for GD, future research should address the contribution of the C421A-BCRP genotype and genistein in the diet to the transplacental passage of glyburide.
Acknowledgments
The authors thank Dr. Robert Robey, National Institutes of Health National Cancer Institute, for the empty vector, wild-type, and C421A-BCRP overexpressing HEK cells.
Authorship Contributions
Participated in research design: Bircsak, Aleksunes.
Conducted experiments: Bircsak, Gupta, Gorczyca.
Provided facilities and expertise in placenta collection: Yuen, Vetrano, Weinberger.
Wrote or contributed to the writing of the manuscript: Bircsak, Aleksunes, Gupta, Yuen, Vetrano, Weinberger.
Footnotes
- Received October 30, 2015.
- Accepted February 2, 2016.
This work was supported by the National Institutes of Health National Institutes of Environmental Health Sciences [Grants ES020522, ES005022, ES007148, ES020721]. Kristin Bircsak was supported by predoctoral fellowships from the American Foundation for Pharmaceutical Education and Pharmaceutical Research and Manufacturers of America.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- ABC
- ATP-binding cassette
- BCA
- bicinchoninic acid
- BCRP
- breast cancer resistance protein
- BSA
- bovine serum albumin
- cDNA
- complementary DNA
- DMSO
- dimethyl sulfoxide
- ER
- estrogen receptor
- EV
- empty vector
- GD
- gestational diabetes
- hCG
- human chorionic gonadotropin
- HEK
- human embryonic kidney 293
- IC50
- half maximal inhibitory concentration
- INN
- international nonproprietary name
- Jmax
- maximum accumulation rate
- Km
- substrate concentration of half the maximum accumulation rate
- LDH
- lactate dehydrogenase
- MDR1
- multidrug resistance protein 1
- MRP
- multidrug resistance-associated protein
- PCR
- polymerase chain reaction
- RPL13A
- ribosomal protein L13A
- WT
- wild-type
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}