


Abstract
The purpose of our study was to establish the localization of the anion transporter Npt1 in liver and the relevance of Npt1 to carrier-mediated hepatic transport of β-lactam antibiotics. Immunocytochemical examination of mouse liver with antiserum forNpt1 showed basolateral (sinusoidal) membrane localization. Function of Npt1 was characterized in Xenopus laevis oocytes. Injection of in vitro-transcribed cRNA into oocytes resulted in an increased uptake of [14C]benzylpenicillin (PCG). The Npt1-mediated uptake was saturable with a Michaelis constant (Km ) of 0.46 ± 0.18 mM and a maximum rate (Vmax) of 46.6 ± 8.5 pmol/60 min/oocyte, and the uptake of [14C]PCG was independent of Na+ and pH, but dependent on chloride ion. Npt1-mediated [14C]PCG uptake was inhibited by several β-lactam antibiotics and probenecid. Oocytes injected with Npt1-cRNA demonstrated significantly enhanced transport activity for other anionic compounds such as [14C]faropenem, [14C]foscarnet and [3H]mevalonic acid, as well as [14C]PCG, compared with water-injected oocytes. In conclusion, Npt1 is suggested to participate in hepatic sinusoidal membrane transport of organic anions such as β-lactam antibiotics as well as inorganic anions for the efflux from hepatocyte-to-blood direction.
Most β-lactam antibiotics are eliminated into urine; however, some derivatives are exclusively excreted into bile (Kind et al., 1970; Barza et al., 1975; Matsui et al., 1982). The influences of lipophilicity (Ryrfeldt, 1971; Ryrfeldt et al., 1973; Forti et al., 1975) and molecular weight (Hirom et al., 1972; Wright and Line, 1980) on the biliary excretion of β-lactam antibiotics have been examined, but the critical factors determining the excretion route have not been fully established. In the process of biliary excretion, the antibiotics must cross both the hepatic sinusoidal and canalicular membranes, so membrane transport processes are expected to be important. We have previously investigated the hepatic uptake mechanism of β-lactam antibiotics through the sinusoidal membrane by the use of freshly isolated rat hepatocytes (Tamai et al., 1985; Tsuji et al., 1986; Terasaki et al., 1986; Tamai and Tsuji, 1987) and the liver uptake index method (Tsuji et al., 1990). We established that most β-lactam antibiotics are taken up via a carrier-mediated process that is common to organic anions, such as probenecid. However, molecular identification of the transport system for the antibiotics has not been achieved.
cDNAs for two bile salt-transporting polypeptides in the sinusoidal plasma membrane have been cloned. The Ntcp have an Na+-dependent bile salt uptake function in mammalian hepatocytes (Hagenbuch et al., 1991; Hagenbuch and Meier, 1994). Further, the cloned oatp mediates Na+-independent transport of bile salts, sulfobromophthalein, estrogen conjugate and a variety of amphipathic compounds (Jacquemin et al., 1994;Shi et al., 1994; Kullak-Ublick et al., 1994;Bossuyt et al., 1993; Yamazaki et al., 1996). However, little is known about the β-lactam antibiotics transport activity of these transporters.
Recently, several mammalian renal Na+-dependent transport systems for Pi have been identified. At the molecular level, there are two distinct types (I and II) (Biber et al., 1996). The type I Na+/Pi cotransporter was originally isolated from rabbit kidney as an NaPi-1 (Werner et al., 1991) and subsequently human NPT1 (Chong et al., 1993) and mouseNpt1 (Chong et al., 1995) were obtained from kidney cortex. Type I transporters share approximately 65% amino acid identity among family members. When expressed in Xenopusoocytes, the type II Na+-coupled Pi transporter shows the characteristics of renal brush border Na+/Pi cotransport, including sigmoidal Na+ dependence, pH dependence, high affinity for Pi (Km of 0.1 mM), and possible regulation by protein kinase C (Magagin et al., 1993; Buschet al., 1994, 1995; Hayes et al., 1995). In contrast to type II, the type I transporter induces a relatively weak Pi-transport activity with a low Pi affinity in Xenopusoocytes (Werner et al., 1991). Interestingly, rabbit NaPi-1 has transport activity for both inorganic (chloride and phosphate ions) and organic anions including PCG, phenol red and probenecid (Buschet al., 1996). Rabbit NaPi-1 and mouse Npt1 were identified in kidney and liver by Northern blot analysis (Werneret al., 1991; Chong et al., 1995), whereas humanNPT1 in liver has not been examined yet. NaPi-1 protein was located by immunohistochemical analysis in the apical membrane of renal proximal tubule cells (Biber et al., 1993). Accordingly, it is suggested that NaPi-1 plays an important role in the excretion of anionic xenobiotics in kidney. However, the role of the type I transporter in the liver is still unclear, because the localization of NaPi-1 has not been established and characterization of organic anion transport activity via type I transporter is still in its early stages.
Our purpose was to investigate the localization of mouse Npt1 in the liver by immunohistochemical study and the activity ofNpt1-mediated uptake of several anionic compounds byXenopus oocytes expression system. We compared the functional properties of β-lactam antibiotic transport mediated byNpt1 with the characteristics of the previously proposed β-lactam antibiotics transport mechanism in the sinusoidal membrane of liver, and concluded that Npt1 may contribute to the efflux of β-lactam antibiotics from hepatocyte to blood not for the uptake into hepatocytes of them.
Materials and Methods
Materials.
[14C]PCG (56 mCi/mmol), [36Cl]NaCl (0.11 GBq/g) and [14C]glycylsarcosine (60 mCi/mmol) were purchased from Amersham International, Ltd. (Buckinghamshire, England). [14C]Tetraethylammonium bromide (1.5 mCi/mmol), NaH32PO4 (1 Ci/mmol) and [14C]taurocholic acid (2 Ci/mmol) were purchased from New England Nuclear (Boston, MA). [3H]Methotrexate (30 Ci/mmol) and [14C]foscarnet (52 mCi/mmol) were purchased from Moravek Biochemicals, Inc. (Brea, CA). [14C]Faropenem (52 mCi/mmol) was kindly supplied by Suntory Co. (Osaka, Japan). [3H]Mevalonolactone (15 Ci/mmol) was purchased from ARC Inc. (St. Louis, MO). Mevalonic acid was prepared by alkaline hydrolysis of the mevalonolactone according to the method reported previously (Kim et al., 1992). Mevalonolactone was treated with 0.05 N NaOH and the resultant hydrolyzed solution was adjusted to pH 7.0 with 0.1 N HCl, and stored at 4°C until use. 2-Ketoglutaric acid and probenecid were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). β-Lactam antibiotics used in this work were kindly supplied as follows: ampicillin anhydrate and cyclacillin from Takeda Chemical Industries (Osaka, Japan); apalcillin and Cefpiramide from Sumitomo Chemical and Industrial Co., Ltd. (Osaka, Japan); benzylpenicillin from Banyu Pharmaceutical Co., Ltd. (Tokyo, Japan); cefixime and ceftizoxime from Fujisawa Pharmaceutical Co. (Osaka, Japan); cefoperazone from Toyama Chemical Co., Ltd. (Toyama, Japan); cephalexin, cephaloridine and cephalotin from Shionogi & Co. (Osaka, Japan); cloxacillin and dicloxacillin from Meiji Seika Kaisha, Ltd. (Tokyo, Japan); nafcillin from Wyeth Japan Co. (Tokyo, Japan); and cephradine from Sankyo Co. (Tokyo, Japan).
Cloning of mouse Npt1 cDNA.
Total RNA was extracted from mouse kidney and polyadenylated [poly(A)+]RNA was purified by affinity chromatography with the use of an oligo(dT)-primed cDNA synthesis kit (Gibco BRL, Gaithersburg, MD). Plaques were screened by hybridization under high-stringency conditions with32P-labeled human NPT1 cDNA (Chong et al., 1993). Six positive clones were isolated, the largest clone was subcloned into the NotI site of pBluscript II SK+ and completely sequenced from both strands (T7 sequencing kit, Pharmacia, Milwaukee, WI) (Mizusawa et al., 1986). This clone was yielding an insert length of 1880 bp excluding the 30-nucleotide poly(A)+ tail. An open-reading frame of 1395 bp was detected, encoding a 465-amino acid polypeptide prior to the TGA termination codon. This clone was identical to mouse Npt1 cDNA reported previously (Chong et al., 1993).
Immunohistochemistry.
For production of an antiserum againstNpt1, a peptide corresponding to a sequence (Glu-Ile-Gln-Asp-Trp-Ala-Lys-Glu-Ile-Lys-Thr-Thr-Arg-Leu) (aa 452-465) within the cDNA-deduced primary structure (Chong et al., 1995) was synthesized and conjugated with keyhole limpet hemocyanin (Sigma, St. Louis, MO). This peptide corresponds to a putative C-terminal intracellular domain of Npt1 (Chong et al., 1995). The specific antibodies were affinity-purified on Cellulofine AM (Seikagaku Kogyo, Tokyo, Japan). For immunostaining ofNpt1, unfixed cryostat sections were used. After microwave irradiation (for 10 min in 10 mM citrate buffer; pH 6.0) and hydrogen peroxide treatment, the sections were incubated overnight in anti-Npt1-specific antibody (2–4 μg/ml) at 4°C.Npt1 proteins were visualized with avidin-biotin-peroxidase complex. To verify the specificity of the immunoreaction, we confirmed that the immunostaining was blocked by the antigen peptide (50 μg/ml) (Hisano et al., 1996).
Transport experiments in Xenopus laevis oocytes.
Oocytes from Xenopus laevis were manually dissected in medium A (96 mM NaCl, 2 mM KCl, 1 mM MgCl2 and 5 mM HEPES adjusted to pH 7.6 with NaOH) and defolliculated in modified Barth’s solution (88 mM NaCl, 1 mM KCl, 2.4 mM NaHCO3, 0.82 mM MgSO4, 0.33 mM Ca(NO3)2, 0.41 mM CaCl2, 100 U/ml penicillin G, 100 mg/ml streptomycin and 10 mM HEPES adjusted to pH 7.5 with NaOH) as described previously (Tamaiet al., 1997).
The plasmids were then linearized with HindIII and used forin vitro transcription after capping. cRNA was dissolved in water at a concentration of 0.2 μg/μl and injected (50 nl) into oocytes, which were assayed for transport activity after 3 to 4 days.
To measure uptake of inorganic and organic anions, oocytes (5–10/individual time point or condition) were incubated in 250 μl of transport buffer (100 mM Na-gluconate, 2 mM K-gluconate, 1 mM Ca-gluconate, 1 mM Mg-gluconate, 10 mM HEPES/NaOH, pH 7.5) containing a radiolabeled compound at 25°C. The uptake was terminated by washing the oocytes three times with 15 ml of ice-cold transport buffer. Washed oocytes were transferred to vials containing 1 ml of 5% sodium dodecyl sulfate to be solubilized and the associated radioactivity was measured with a liquid scintillation counter. Usually uptake values were expressed as the cell-to-medium ratio or uptake coefficient obtained by dividing uptake amount or uptake rate by the concentration of the substrates in the medium.
Results
Localization of Npt1 in mouse liver.
The cellular expression and membrane localization of Npt1 in the liver were investigated by immunohistochemistry (fig.1). In the kidney cortex, immunoreactivity for Npt1 was localized at the luminal membranes of the proximal tubules (data not shown). This distribution is similar to that in rabbit kidney (Biber et al., 1993). In the liver, Npt1 is present on the basolateral (sinusoidal) membrane of hepatocytes. This localized expression in liver and kidney is the same as that of oatp protein in rats (Shi et al., 1994).

Immunohistochemical localization of Npt1 in mouse liver. Frozen sections of liver (A and B) were stained with anti-Npt1-specific antibody (2–4 μg/ml) in the absence (A) or presence (B) of 50 μg/ml peptide antigen, and immunohistochemical analysis was performed as described in “Materials and Methods.” Immunoreactivity shown by arrows indicates sinusoidal membrane sides. Bars represent 100 μm.
Expression of PCG uptake activity in Xenopus laevis oocytes injected with mouse Npt1.
To confirm the expression of functional inorganic anion transport activity of mouse Npt1, we measured the uptake of 36Cl− and H32PO42− in an Npt1-expressingXenopus laevis oocyte heterologous expression system. Uptake values of 36Cl− in Npt1 cRNA- and water-injected Xenopus oocytes were 0.25 ± 0.016 and 0.15 ± 0.020 μl/oocyte, and those of H32PO42− were 0.11 ± 0.011 and 0.08 ± 0.007 μl/oocyte (mean of five determinations ± S.E.), respectively, at 60 min. Because although the expressed activity was not so high, uptake values of phosphate and chloride ions were statistically different (P < .05) between Npt1 cRNA- and water-injected oocytes, mouse-Npt1 appears to have transport activity for phosphate and chloride ions.
Time course and concentration dependence of the uptake of [14C]PCG.
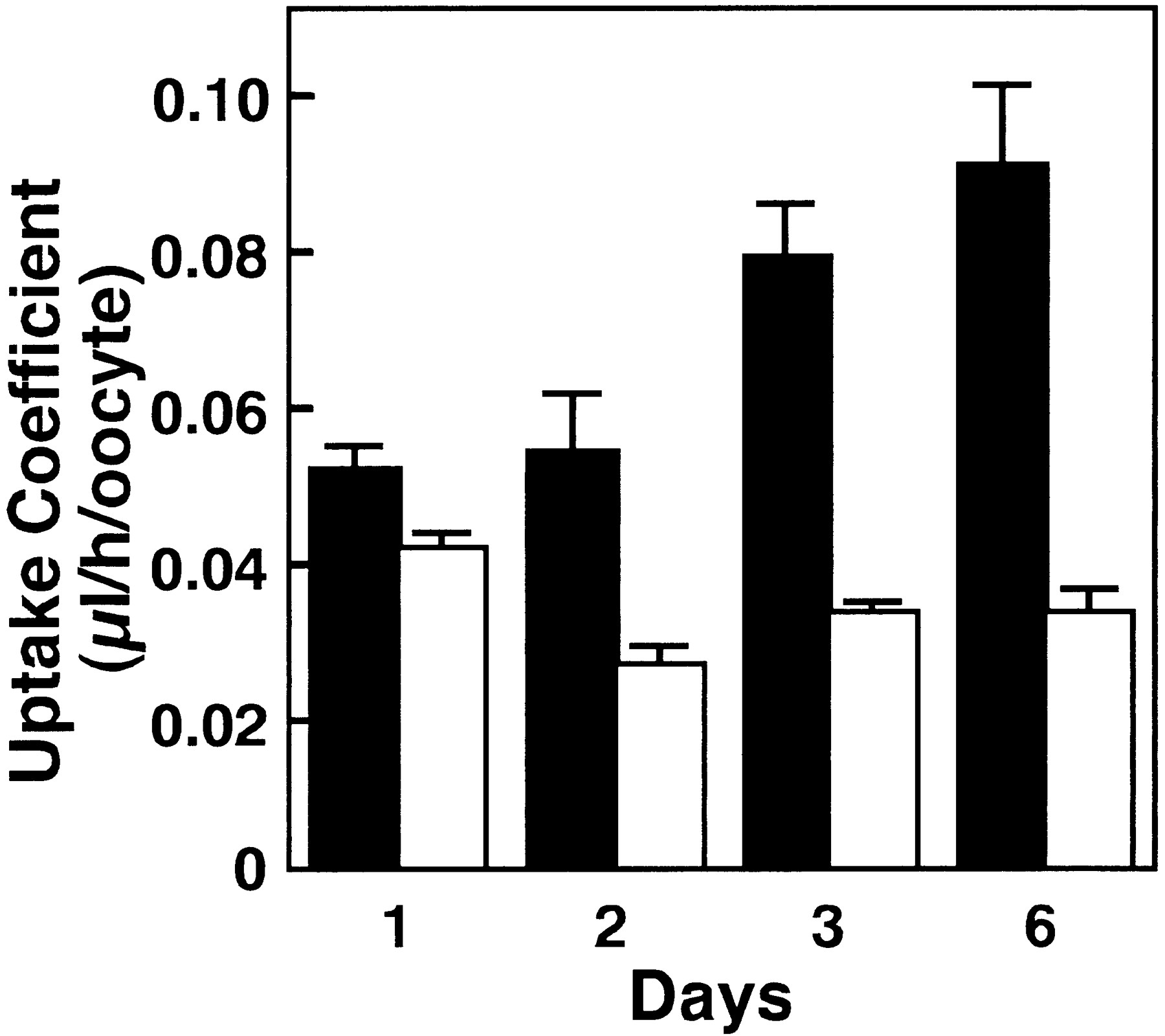
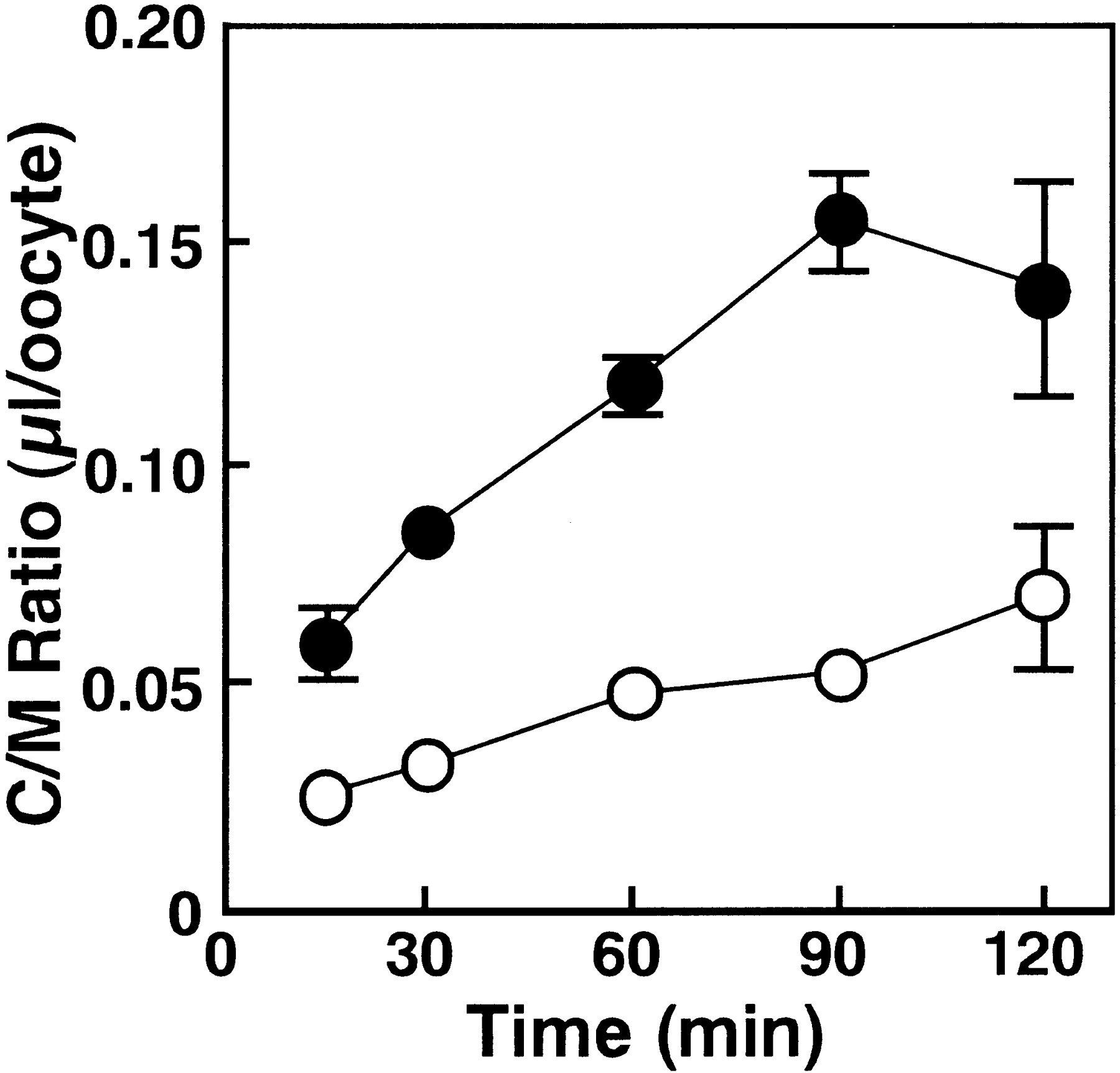
To clarify whether or not mouseNpt1 has organic anion transport activity, we investigated the transport of β-lactam antibiotics in cRNA-injectedXenopus oocytes. In this study, the anionic β-lactam antibiotic, PCG was selected as the substrate, because we have previously investigated the hepatic uptake mechanism of PCG through the sinusoidal membrane in hepatocytes (Tamai et al., 1985;Tsuji et al., 1986; Terasaki et al., 1986). The uptake of [14C]PCG in Npt1 cRNA-injected oocytes was significantly higher than that in water-injected oocytes on day 1 and was further enhanced on days 2, 3 and 6 after injection of the cRNA (fig. 2). Subsequent uptake studies were carried out more than 3 days after cRNA injection. Figure3 shows the time course of [14C]PCG uptake by Npt1 cRNA- and water-injectedXenopus oocytes. The uptake by Npt1 cRNA-injected oocytes increased linearly for 90 min, although no significant uptake was observed in the water-injected Xenopus oocytes. Accordingly, all subsequent initial uptake studies were performed at 60 min.

Functional expression of Npt1 in X. laevis oocytes. The uptakes of [14C]PCG (20 μM) inNpt1 cRNA-(10 ng) injected (filled columns) and water-(50 nl) injected (open columns) oocytes were measured for 60 min at 25°C on days 1, 2, 3 and 6 after injection in transport buffer (pH 7.5). The results are shown as means ± S.E. of 5 to 10 oocytes.

Time course of [14C]PCG uptake inNpt1 cRNA-injected oocytes. The uptakes of [14C]PCG (20 μM) in Npt1 cRNA-(10 ng) injected (filled circles) and water-(50 nl) injected (open circles) oocytes were measured at 25°C in chloride-free 10 mM HEPES-NaOH buffer (pH 7.5). The results are shown as means ± S.E. of 5 to 10 oocytes.
The concentration dependency of the initial uptake of [14C]PCG was examined. To evaluateNpt1-derived uptake, the result was expressed after subtraction of the uptake by water-injected oocytes from that by cRNA-injected oocytes (fig. 4). Eadie-Hofstee plot (shown in fig. 4 inset) suggested that a single saturable process was involved in Npt1-mediated [14C]PCG transport. Nonlinear least-squares analysis yielded an apparent Km of 0.46 ± 0.18 mM and Vmax of 46.6 ± 8.5 pmol/60 min/oocyte.

Concentration dependence of PCG uptake inNpt1 cRNA-injected oocytes. Uptake at various concentrations of PCG was measured for 60 min at 25°C in transport buffer (pH 7.5). Each solution contained an appropriate concentration of Na-gluconate to be isotonic. The data were obtained by subtraction of the uptake by water-injected oocytes from that by Npt1 cRNA-injected oocytes. Inset, Eadie-Hofstee plot for the saturable process of concentration-dependent PCG uptake. The results are shown as means ± S.E. of 5 to 10 oocytes.
Effect of extracellular pH, Na+ and Cl− on [14C]PCG uptake.
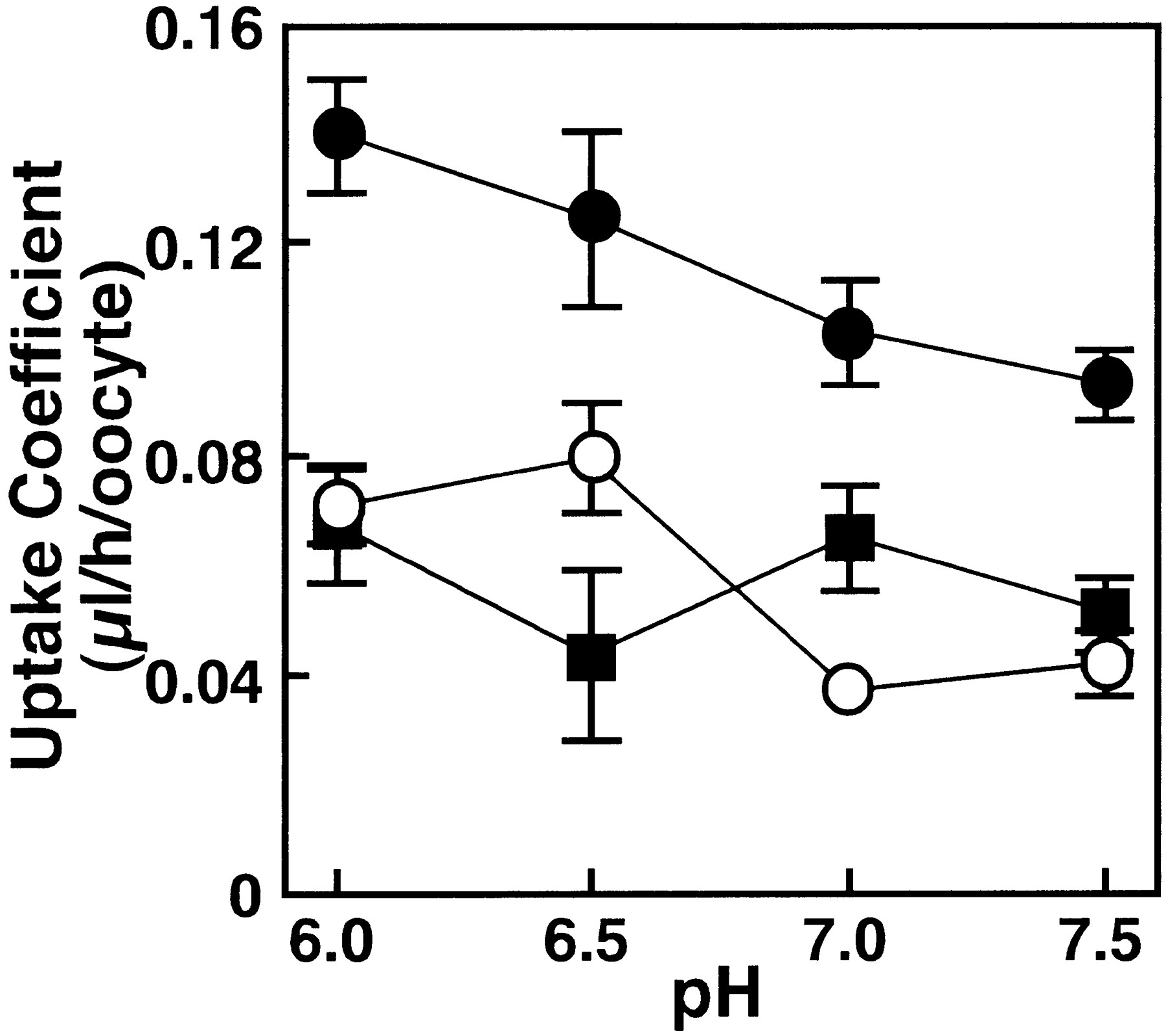
The pH, Na+ and Cl− dependences of [14C]PCG transport mediated by cRNA- and water-injected Xenopus oocytes are shown in figures 5, 6 and 7. Figure 5shows the effect of pH on [14C]PCG uptake byNpt1. When the pH of the medium was varied from 7.5 to 6.0, PCG uptake by the water-injected oocytes increased slightly. After subtracting PCG uptake by control oocytes, there was no significant difference in the Npt1-mediated uptake values at each pH. Figure 6 shows that the uptake of [14C]PCG is not significantly altered upon replacement of Na+ with K+. Figure7 shows the effect of chloride ions on uptake of [14C]PCG. Uptake of [14C]PCG was significantly decreased with increase of Cl− concentration in the transport medium.
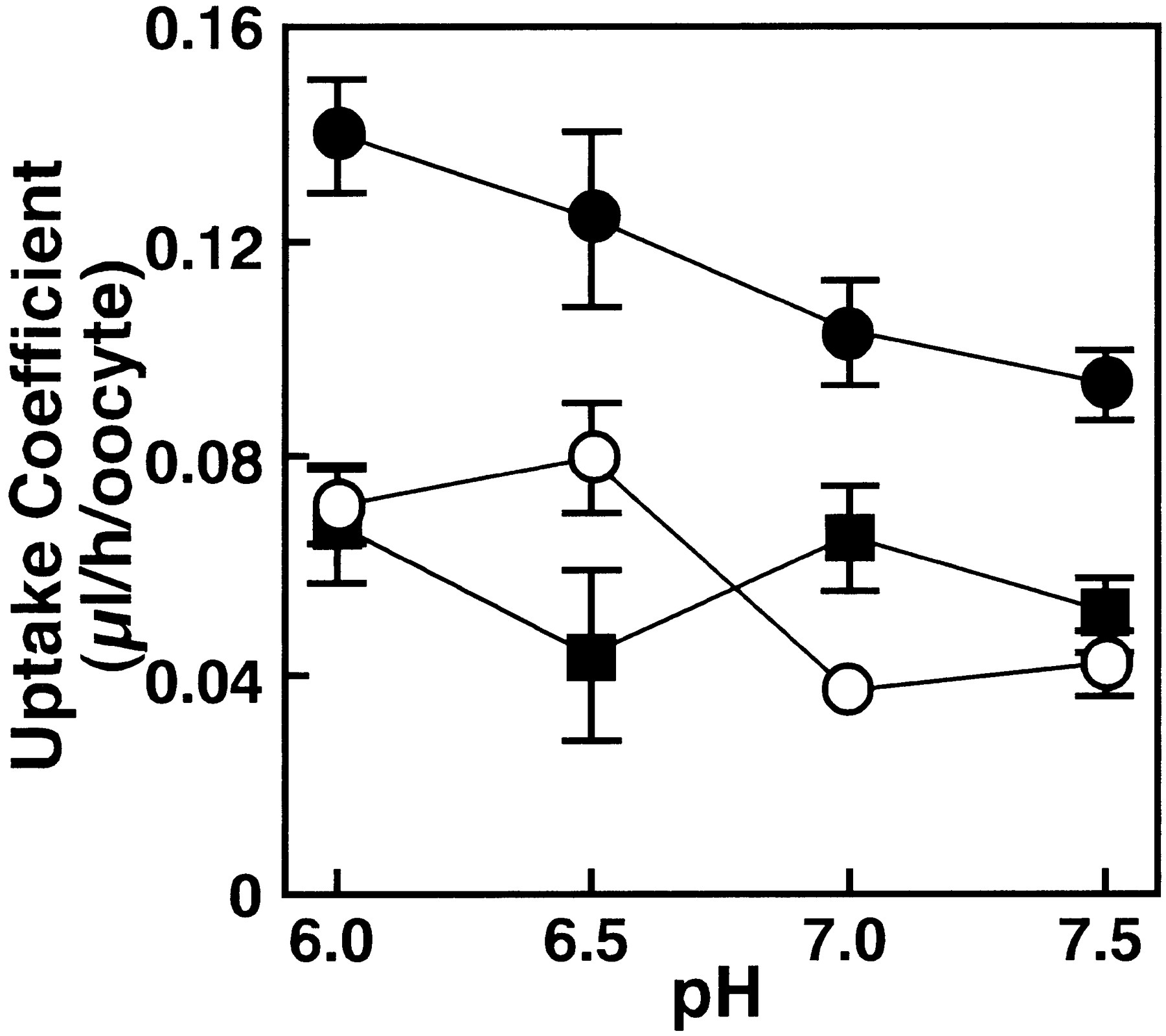
Effect of extracellular pH on PCG uptake inNpt1 cRNA-injected oocytes. The uptake of [14C]PCG (20 μM) in Npt1 cRNA (closed circles) and water (open circles) injected oocytes were measured for 60 min at 25°C in transport buffer. pH of transport buffer was adjusted by Mes-NaOH for pH 6.0 and by HEPES-NaOH for pH 6.5, 7.0 and 7.5. Each solution contained an appropriate concentration of Na-gluconate to be isotonic. The uptake shown by closed squares is theNpt1-dependent uptake obtained as the difference between the uptakes in Npt1 cRNA-injected and water-injected oocytes. The results are shown as means ± S.E. of 5 to 10 oocytes.

Effect of extracellular Na+ on PCG uptake in Npt1 cRNA-injected oocytes. The uptakes of [14C]PCG (20 μM) in Npt1 cRNA-(10 ng) injected (filled columns) and water-(50 nl) injected (open columns) oocytes were measured for 60 min at 25°C in the presence or absence of Na+ in transport buffer (pH 7.5). The results are shown as means ± S.E. of 5 to 10 oocytes.

Effect of extracellular Cl− on PCG uptake in Npt1 cRNA-injected oocytes. The uptakes of [14C]PCG (20 μM) in Npt1 cRNA-(10 ng) injected (filled circles) and water-(50 nl) injected (open circles) oocytes were measured for 60 min at 25°C in 10 mM HEPES-NaOH buffer (pH 7.5) containing various concentration of Cl− (0, 5, 25 and 100 mM). Each solution contained an appropriate concentration of Na-gluconate to be isotonic. The results are shown as means ± S.E. of 5 to 10 oocytes.
Inhibition of [14C]PCG uptake by various β-lactam antibiotics and organic anionic compounds.
To examine the range of β-lactam antibiotics that can be taken up by Npt1, we examined the inhibitory effect of several β-lactam antibiotics on theNpt1-mediated [14C]PCG uptake (table1). The concentrations of [14C]PCG and the β-lactam antibiotics were 20 μM and 5 mM, respectively. All β-lactam antibiotics examined, including both anionic and zwitterionic derivatives had significant inhibitory effects (P < .05). Anionic derivatives tended to have more potent inhibitory effects than zwitterionic derivatives. One mM probenecid significantly inhibited the uptake of 20 μM [14C]PCG. This is similar to the result obtained in our hepatic sinusoidal membrane transport studies (Terasaki et al., 1986). A dicarboxylic acid, 2-ketoglutaric acid, did not inhibit the uptake of [14C]PCG. This result indicates that the substrate selectivity of Npt1 is distinct from that of a recently cloned OAT1 which transports β-lactam antibiotics across the renal epithelial basolateral membrane (Sekine et al., 1997).
Inhibitory effect of β-lactam antibiotics and organic anions on the uptake of [14C]PCG in Npt1 cRNA-injected oocytes
Transport of various organic anionic compounds.
Substrate specificity of Npt1 was assessed by measuring the transport of several organic anionic compounds. Uptakes of [14C]PCG, [14C]faropenem, [14C]foscarnet and [3H]mevalonic acid by cRNA-injected oocytes were significantly increased compared with those by water-injected oocytes (table 2), whereas the uptakes of [3H]taurocholate, [14C]glycylsarcocine, [3H]methotrexate and [14C]tetraethylammonium were not increased byNpt1-cRNA expression. These results show thatNpt1 basically has transport activity for anionic compounds and is distinct in substrate specificity from previously cloned organic anion transporters, Ntcp (Hagenbuch et al., 1991), oatp (Jacquemin et al., 1994) and OAT-K1 (Saito et al., 1996), as well as the oligopeptide transporter, PepT1 (Tamaiet al., 1997).
Substrate selectivity of Npt1
Discussion
The mechanism of uptake of β-lactam antibiotics into hepatocytes is of biochemical and pharmacological interest, because these antibiotics can be classified into two groups, i.e., urinary excretion type and biliary secretion type, based on the characteristic elimination pathway. It is possible that the hepatic transporters involved in the uptake and/or secretion play a crucial role in determining the pathway (renal or biliary) of elimination of β-lactam antibiotics. It has been shown that ampicillin, ceftriaxone and cefodizime are probably secreted via cMOAT across the bile canalicular membrane (Oude Elferink, 1989; Verkade et al., 1990;Sathirakul et al., 1993, 1994), whereas the relevant transport system in the sinusoidal membrane has not yet been characterized. In addition, specific transporter should be involved in the hepatic efflux to blood, because hepatic uptake process is not necessarily rate limiting for biliary excretion and certain amount of drugs taken up by hepatocytes are thought to be back-fluxed into blood (Tamai et al., 1985; Tsuji et al., 1986; Tamai and Tsuji, 1987).
Previous studies in this laboratory have revealed that β-lactam antibiotics such as benzylpenicillin, cefpiramide, cefazolin and cephalexin are taken up into freshly isolated rat hepatocytes, which are considered to be a model of the hepatic sinusoidal membrane, via a carrier-mediated transport process (Tsuji et al., 1986). To examine the molecular mechanism of this transport system on the sinusoidal membrane side, we investigated the role of mouse phosphate transporter, Npt1, in β-lactam antibiotic uptake into hepatocytes, because the rabbit phosphate transporter, NaPi-1, exhibited transport activity for organic anions, including benzylpenicillin (Busch et al., 1996).
Polyclonal antibody raised against Npt1 enabled the study of subcellular distribution. Although Npt1 has been identified in mouse kidney and liver by Northern blot analysis, its cellular localization in the liver is not known. Our study shows thatNpt1 is detectable by immunostaining in the sinusoidal (basolateral) membrane. This localization is consistent with a role ofNpt1 in the transport of β-lactam antibiotics between blood and hepatocytes.
In this study, Npt1 cRNA-injected oocytes showed significantly enhanced transport of [14C]PCG as compared with water-injected oocytes. The Km forNpt1-mediated PCG uptake was 0.46 mM. Our previous study showed that Km for PCG uptake is 0.47 mM in freshly isolated rat hepatocytes (Tsuji et al., 1986). Although there is a species difference, these Km values are very similar. PCG transport activities via Npt1 and in freshly prepared rat hepatocytes were both independent of Na+ and pH, and were inhibited by addition of probenecid. The result obtained in the uptakes by isolated hepatocytes previously was that zwitterionic derivatives exhibited lower affinity than anionic ones (Tamai et al., 1985). This pattern of the inhibitory effect is similar toNpt1 but is not identical as obtained in the less or comparative effect of anionic derivatives such as cephalothin and ceftizoxime with zwitterionic derivatives. These results indicate thatNpt1 possesses the similar characteristics as the predicted transport system for β-lactam antibiotics in the hepatic sinusoidal membrane. However, it was proved to be incorrect to compare directly the results obtained in this study with those in our previous study in freshly isolated rat hepatocytes, because PCG uptake by the Npt1 cRNA-injected oocytes was measured in a medium in which chloride was replaced with gluconate.
Our results suggest that PCG transport activity via Npt1 is affected by chloride ion (fig. 7). At an early stage of this study we predicted that Npt1 works as an organic anion/Cl−antiporter. However, because our preliminary result did not support the idea (data not shown), the mechanism by which Npt1 mediates organic anion transport was not established. Considering the physiological concentration of chloride, there is a possibility that the chloride ion concentration in blood (∼100 mM) may not be optimal for Npt1-mediated organic anion uptake from blood into hepatocytes. Because the concentration of chloride ion in hepatocytes is approximately 15 mM and Npt1 efficiently transports PCG at this chloride concentration, Npt1 protein may facilitate secretion of organic anions from hepatocytes to blood, and specific transporter other than Npt1 is supposed to have a role in the hepatic sinusoidal uptake of β-lactam antibiotics.
Subsequent to uptake into hepatocytes via specific organic anion transport systems, intracellular compounds can be partially secreted back into the sinusoidal space again and/or be transported via canalicular membrane transporters into bile. Sinusoidal efflux has been demonstrated for compounds such as bilirubin (Wolkoff et al., 1987), DBSP (Nijssen et al., 1991) and harmol sulfate (de Vries et al., 1985). The efflux process may be mediated by the same carrier that catalyzes the uptake of these compounds, but indirect evidence exists that it may involve separate mechanisms (Nijssen et al., 1991). Although the sinusoidal efflux of β-lactam antibiotics has not been studied yet, if β-lactam antibiotics derivatives are secreted via such a mechanism,Npt1 may play a role in the back flux of β-lactam antibiotics from hepatocytes to blood.
A noteworthy feature of Npt1 is its wide substrate selectivity, covering not only β-lactam antibiotics derivatives, but also other organic anions such as penem antibiotic, faropenem, antivirus agent, foscarnet and a native weak acid, mevalonate. In this respect, Npt1 is distinct from previously cloned organic anion transporters in the liver and kidney. Further study of the transport mechanisms and substrate specificity of Npt1 protein may reveal the role of Npt1 in the liver.
In conclusion, we have demonstrated that Npt1 protein functions as a transporter of β-lactam antibiotics and other organic anions in the liver. Because Npt1 activity is reduced in the presence of high concentration of chloride ion, Npt1 is presumed to transport β-lactam antibiotics and other organic anion from hepatocyte to blood, but not for hepatic uptake physiologically. For the uptake of β-lactam antibiotics from blood to hepatocyte transporter other than Npt1 should be present in the sinusoidal membranes. These kind of study will lead to the understanding of determinant of elimination pathway, namely biliary or urinary excretion for β-lactam antibiotics.
Footnotes
-
Send reprint requests to: Prof. Akira Tsuji, Department of Pharmacobio-Dynamics, Faculty of Pharmaceutical Sciences, Kanazawa University, 13-1 Takara-machi, Kanazawa 920-0934, Japan.
-
↵1 This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science and Culture, Japan, by grants from the Japan Health Sciences Foundation, the Drug Innovation Project and the Japan Research Foundation for Clinical Pharmacology and by CREST (Core Research for Evolutional Science and Technology) of Japan Science and Technology Corporation (J.S.T.).
- Abbreviations:
- PCG
- benzylpenicillin
- HEPES
- N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid
- MES
- 2-(N-morpholino)ethanesulfonic acid
- Pi
- inorganic phosphate
- cRNA
- complementary RNA
- Ntcp
- Na+-taurocholate-cotransporting polypeptides
- oatp
- organic anion-transporting polypeptide
- Received February 27, 1998.
- Accepted May 11, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}