


Abstract
For more than 20 years it has been known that neonatal exposure to phenobarbital results in a delayed, but permanent overexpression of drug-metabolizing enzymes in adult male and female rats. Accordingly, to identify the specific isoform(s) of P450 responsible for the imprinted overexpression of hepatic monooxygenases, we have monitored the developmental profile of some dozen hepatic P450 isoforms in 4- to 150-day-old male and female rats neonatally treated with the barbiturate. Some of the cytochrome P450s (CYP), i.e., CYP2A1, 2A2, 2C6, 3A1, and 3A2, exhibit the typical transient response in which isoform levels (mRNA, protein, and/or specific catalytic activity) rise precipitously at the time of phenobarbital administration and rapidly decline to preinduction levels after withdrawal of the barbiturate. Other isoforms, i.e., CYP1A1, 1A2, 2C7, 2C11, 2C12, and 2C13, were neither constitutively expressed nor phenobarbital inducible in the neonate. Only one of these isoforms, female predominant (M:F,∼1:2) CYP2C7, exhibited a barbiturate-induced delayed, but persistent ∼30 to 50% overexpression from puberty through adulthood. We propose that at the time of exposure, neonatally administered phenobarbital produces a “silent” programming defect resulting in a delayed, but persistent overexpression of the isoform, contributing, at least in part, to a permanent elevation of hepatic drug-metabolizing enzyme activities.
Perinatal exposure of laboratory animals to subteratogenic doses (i.e., not producing malformations) of phenobarbital can result in a multitude of long-term reproductive, growth, hepatic, and neural dysfunctions. These defects include reduced sexual behavior (Clemens et al., 1979), decreased serum gonadotropin levels and infertility (Gupta et al., 1980, 1982), abnormal circulating testosterone secretory profiles (Wani et al., 1996), subnormal growth rates (Gupta et al., 1980; Agrawal et al., 1995a), decreased hepatic monoamine oxidase levels (Soliman and Richardson, 1983), altered seizure susceptibility (Sobrian and Nandedkar, 1986), and elevated brain concentrations of neurotransmitters and locomotor and learning deficits (Middaugh et al., 1981; Vorhees, 1983, 1985; Sobrian and Nandedkar, 1986). Of particular relevance to this study are reports demonstrating that neonatal administration of phenobarbital can induce a delayed but permanent elevation in the activities of several hepatic monooxygenases, e.g., total cytochrome P450 (CYP), ethoxycoumarin O-deethylase, ethylmorphine N-demethylase, and hexobarbital hydroxylase (Yanai, 1979; Bagley and Hayes, 1985; Agrawal et al., 1995a). As expected, neonatal administration of the barbiturate induces an almost immediate increase in the activities of the hepatic monooxygenases that declines to noninduction levels when treatment ceases. Contrary to the well known transient effects of phenobarbital, at approximately the time of sexual maturity when gender-dependent differences in hepatic monooxygenases appear (males > females), a second “round” of enzyme induction occurs that persists throughout adulthood.
The mechanism(s) by which phenobarbital produces these latently expressed defects is unknown. Because the half-life of the barbiturate in adult rats is measured in hours (Valerino et al., 1974) and is no more than days in newborns (Ishizaki et al., 1981), it is unlikely, for example, that the delayed postpubertal elevation in hepatic monooxygenase activities can be explained by the persistence of the neonatally administered phenobarbital.
In this study, we have administered phenobarbital to newborn rats at a therapeutic-like dose (Albright and Burnham, 1980; Carl and Smith, 1988), which is considerably below the threshold for inducing morphological and behavioral defects (Vorhees, 1983, 1985), to identify the specific isoform(s) responsible for the persistent elevation in hepatic drug-metabolizing enzyme activities. In this regard, we have examined the expression (mRNA, protein, and/or specific catalytic activity) of some dozen constituent, inducible, sex-dependent and -independent hepatic P450 isoforms from the time of phenobarbital administration in the first week of life through adulthood.
Materials and Methods
Animals were housed in the University of Pennsylvania Laboratory Animal Resources facility under the supervision of certified laboratory animal medicine veterinarians and were treated according to a research protocol approved by the University's Institutional Animal Care and Use Committee. At all times, animals were housed on hardwood bedding in plastic cages, with water and commercial rat diet supplied ad libitum. The animal quarters were air conditioned (20–23°C) and maintained on a 12-h light/dark cycle (lights on at 8:00 AM). After a 2- to 3-week acclimation period in our facilities, the animals were bred by randomly housing two adult female Sprague-Dawley rats [Crl:CD(SD)BR] with an individual adult male of the same strain. On the day of parturition, all litters were mixed and randomly assigned to the dams at 10 pups per litter, with a sex ratio of 1:1 or as close to this ratio as possible. Starting within 24 h after birth and continuing through the first 7 days of life, all of the pups in a litter were injected s.c. with either 40 mg/kg b.wt. phenobarbital sodium (Sigma Chemical Co., St. Louis, MO) or an equivalent sodium concentration of NaCl diluent, pH 9.l, administered as 5 μl/g b.wt. The pups were weaned at 24 days of age.
Male and female offspring were sacrificed by decapitation at 4, 8, 25, 45, 65, and 150 days of age. Livers were quickly removed and perfused with ice-cold saline. Each liver was quickly minced; a portion reserved for mRNA determination was plunged into liquid nitrogen and subsequently stored at −70°C. The remaining minced liver was used for microsome preparation. (Livers from only the 4-day-old pups were pooled, i.e., three livers per sample.)
Total RNA was isolated from ∼0.5 g of individual rat liver by a single-step guanidinium-thiocyanate method (Chomczynski and Sacchi, 1987). RNA samples from individual livers were fractionated by electrophoresis under denaturing conditions in 1.2% agarose gels containing 1× 3-(N-morpholino)-propanesulfonic acid buffer and 1.28% formaldehyde. The RNA was transferred to GeneScreen nylon membranes (DuPont/NEN, Boston, MA) by capillary transfer in 10× standard saline citrate and then fixed to the filters by UV cross-linking. Prehybridizations and hybridizations in Rapid-hyb buffer (Amersham, Arlington Heights, IL) with32P-labeled oligonucleotide probes were performed with high-stringency washings. The washed blots were wrapped in clear plastic and exposed to X-ray films with two intensifying screens at −70°C for 1 to 3 days. The nucleotide sequence of oligonucleotide probes for CYP1A1, CYP1A2, CYP2C6, CYP2C7, CYP2C11, CYP2C12, CYP2C13 (Waxman, 1991), CYP3A1 (Gemzik et al., 1992), and CYP3A2 (Ram and Waxman, 1991) have been reported.
Evidence that RNA was equally loaded and transferred was obtained by equivalent intensity of ethidium bromide staining of 18S and 28S rRNA bands (Schuetz et al., 1990). Furthermore, the rat 18S rRNA oligonucleotide probe was used as a control to verify the consistency and integrity of RNA loading (Ramsden et al., 1993). Quantitation of the mRNA by laser densitometry of the X-ray films was kept within the linear range as established by slot blot hybridizations and normalized to the 18S rRNA signals in each lane as well as to two control samples repeatedly run on every blot.
Hepatic microsomes were prepared from individual rat livers (Shapiro and Szczotka, 1984) and then assayed for individual P450s by Western blotting and/or by measurement of their selective catalytic activities (Agrawal et al., 1995b; Agrawal and Shapiro, 1996b). Briefly, 10 μg of microsomal protein was electrophoresed on 0.75-mm-thick SDS-polyacrylamide (7.5%) gels and electroblotted onto nitrocellulose filters. The blots were probed with monoclonal anti-rat CYP2C11; anti-rat CYP1A1/2 (Oxford Biomedical Research, Oxford, MI); anti-rat CYP2C12/13 (kindly provided by Dr. Marika Rönnholm, Huddinge University Hospital, Huddinge, Sweden) mouse IgG; polyclonal anti-rat CYP2C7 (kindly provided by Dr. Stelvio M. Bandiera, The University of British Columbia, Canada); and anti-rat CYP3A1/2 (Human Biologics, Phoenix, AZ) rabbit IgG and detected with an enhanced chemiluminescence kit (Amersham; Pampori et al., 1995). Quantitation of the relative protein levels was by laser densitometry of the X-ray films and by normalizing protein signals to two control samples repeatedly run on all blots.
Testosterone metabolites, including 2α- and 16α-, 7α-, 15α-, and 6β-hydroxylases, reflective of the activity levels of CYP2C11, CYP2A1, CYP2A2 and CYP3A2 proteins, respectively (Waxman, 1991;Schenkman, 1992), and female-specific3testosterone 5α-reductase (coincidental with CYP2C12) were assayed according to our methods as described previously (Pampori et al., 1991;Agrawal et al., 1995b). Male-predominant, multi-P450-dependent microsomal hexobarbital hydroxylase was measured as reported in Shapiro and Szczotka (1984).
All data were subjected to ANOVA. Significant differences were determined with t statistics and the Bonferroni procedure for multiple comparisons.
Results
In agreement with our previous report (Agrawal et al., 1995a), we observed a >10-fold transient increase in hepatic microsomal hexobarbital hydroxylase in 8-day-old male and female pups treated with phenobarbital during the first week of life. Shortly thereafter, the enzyme activity declined to normal, but was again “re-induced” above control levels when measured at 50 to 60 days of age. Although neonatal exposure to the barbiturate had no effect on the sexually dimorphic expression of the enzyme (concentrations being ∼5 to 6 times greater in males), we observed a statistically significant ∼25% higher hexobarbital hydroxylase activity in exposed males and females at 150 days of age (data not shown). Although far from novel (Yanai, 1979; Bagley and Hayes, 1985; Agrawal et al., 1995a), our finding of a latent overexpression of the enzyme had to be confirmed to demonstrate the suitability of the animals for our present study to identify the specific P450 isoform(s) responsible for the elevated hexobarbital hydroxylase activities.
Neonatal administration of phenobarbital induced a transient 4- to 5-fold increase in both male and female concentrations of CYP3A1 and 3A2 mRNA at 4 and 8 days of age that had declined to normal when next measured at 25 days of age (Fig. 1). In contrast, CYP2C7, 2C11, 2C12, and 2C13 mRNAs were neither expressed constitutively in the pups, nor were they responsive to barbiturate induction at this time. In another variation, although CYP2C6 mRNA was undetectable in the livers of 4- and 8-day-old untreated male and female pups, neonatal exposure to the barbiturate produced a transient, but dramatic expression of the isoform that equaled or exceeded adult levels. Transcript levels, however, had returned to normal when next determined at 25 days of age.

Hepatic CYP mRNAs from 4- to 150-day-old male and female rats neonatally treated with either 40 mg/kg b.wt. of phenobarbital (PB) or an equivalent sodium concentration of NaCl (control) for the first 7 days of life. Procedural details are described in Materials and Methods. Each data column is a mean ± S.D. with an n ≥ 6 and expressed as a percentage of the mean value of the group with the highest concentration (designated 100%) of the isoform.†P < .01 compared with same age males receiving the same neonatal treatment. *P < .01 compared with control animals of the same age and gender.
Sexually dimorphic expression levels of the isoform mRNAs in young (65-day-old) and mature (150-day-old) rats were in agreement with previous reports (Pampori and Shapiro, 1996). That is, CYP2C6 and 2C7 were female predominant (M:F, 1:2); CYP2C11, 2C13, and 3A2 were male specific; CYP2C12 was female specific; and CYP3A1 was gender independent (Fig. 1).
Neonatal administration of phenobarbital appeared to have no long-term consequences on the hepatic expression of CYP2C6, 2C12, 3A1, and 3A2 mRNAs in either adult male or female rats. In contrast, although early exposure to the barbiturate was ineffective in inducing CYP2C7, 2C11, and 2C13 mRNAs in the pups, all three isoforms were overexpressed in adulthood (Fig. 1). Overexpression of male-specific CYP2C11 and 2C13 mRNAs was limited to males, whereas female-predominant CYP2C7 transcript levels were overexpressed ∼30 to 50% above normal in both sexes. [Irrespective of age, gender, or treatment, CYP1A1 and 1A2 mRNAs were undetectable in all samples (data not shown).]
In general, protein levels for the individual CYP isoforms reflected mRNA levels (Fig. 2). That is, in agreement with transcript levels, CYP2C7, 2C11, 2C12, and 2C13 proteins were neither expressed constitutively in the pups nor were they responsive to phenobarbital induction at this time. Results with anti-rat CYP3A1/2 indicated that like the mRNA findings, a barbiturate-induced transient 3- to 4-fold increase in both male and female concentrations of the proteins at 4 and 8 days of age had declined to normal when next measured at 25 days of age.

Hepatic CYP proteins from 4- to 150-day-old male and female rats neonatally treated with either 40 mg/kg b.wt. of phenobarbital (PB) or an equivalent sodium concentration of NaCl (control) for the first 7 days of life. Procedural details are described in Materials and Methods. Each data column is a mean ± S.D. with an n ≥ 6 and expressed as a percentage of the mean value of the group with the highest concentration (designated 100%) of the isoform.†P < .01 compared with same age males receiving the same neonatal treatment. *P < .01 compared with control animals of the same age and gender.
The gender differences for the individual P450 proteins were the same as found for the mRNAs (Figs. 1 and 2). [Because our antibody could not discriminate between CYP3A1 and 3A2, and CYP3A1 is basically a sex-independent inducible form constituently expressed at very low levels (Gemzik et al., 1992; Pampori and Shapiro, 1996), our findings with anti-CYP3A1/2 indicate a male dependence with concentrations in adult females reflecting only CYP3A1 protein.]
In agreement with transcript levels (Fig. 1), neonatal administration of phenobarbital produced a latently expressed ∼30 to 50% above normal overinduction of CYP2C13 and 2C7 proteins in adult males and in adults of both sexes, respectively. In contrast, the latent overexpression observed in CYP2C11 mRNA was not translated into above-normal protein levels (Fig. 2). CYP1A1/2 proteins, like their respective mRNAs, were undetectable in all groups (data not shown).
The specific catalytic activities of several P450 isoforms can be determined by measuring the levels of selective testosterone-metabolizing enzymes (Waxman, 1991; Schenkman, 1992). In this regard, the effects of age, gender, and neonatal phenobarbital treatment on CYP3A1/2-dependent testosterone 6β-hydroxylase and CYP2C12-associated testosterone 5α-reductase were the same as observed at the mRNA and protein levels (Fig.3). In general, CYP2C11-dependent testosterone 2α-hydroxylase activities were reflective of CYP2C11 mRNA and protein with the exception that the barbiturate-induced delayed overexpression in mRNA was represented at neither the catalytic nor protein levels. In agreement, testosterone 16α-hydroxylase, another CYP2C11-dependent catalytic activity, exhibited no overinduction in adult rats (data not shown). Both CYP2A1-dependent testosterone 7α-hydroxylase and CYP2A2-dependent testosterone 15α-hydroxylase exhibited their characteristic sexual dimorphisms (female- and male-dependent, respectively) and a transient induction by neonatal phenobarbital. Testosterone 15α-hydroxylase, however, responded to the neonatally administered barbiturate with a latently overexpressed induction in adult males and females.
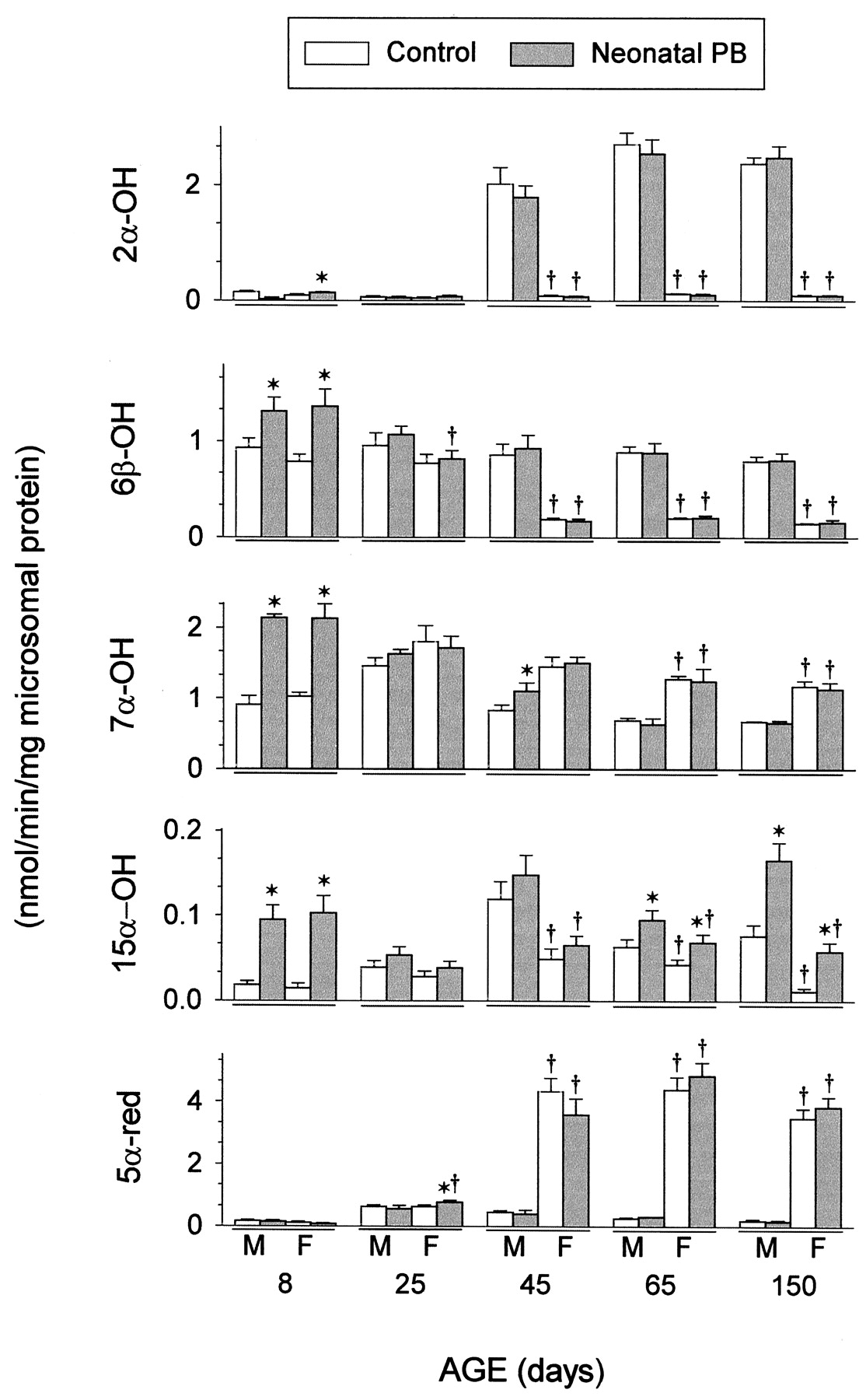
Catalytic levels of hepatic microsomal CYP2C11-dependent testosterone 2α-hydroxylase (2α-OH), CYP3A1/2-dependent testosterone 6β-hydroxylase (6β-OH), CYP2A1-dependent testosterone 7α-hydroxylase (7α-OH), CYP2A2-dependent testosterone 15α-hydroxylase (15α-OH), and CYP2C12-associated testosterone 5α-reductase (5α-red) from 8- to 150-day-old male and female rats neonatally treated with either 40 mg/kg b.wt. of phenobarbital (PB) or an equivalent sodium concentration of NaCl (control) for the first 7 days of life. Procedural details are described in Materials and Methods. Each data column is a mean ± S.D. with an n ≥ 6 and expressed as a nanomole of 2α-hydroxytestosterone (2α-OH), 6β-hydroxytestosterone (6β-OH), 7α-hydroxytestosterone (7α-OH), 15α-hydroxytestosterone (15α-OH), or 5α-dihydrotestosterone (5α-red) produced per minute per milligram of microsomal protein.†P < .01 compared with same age males receiving the same neonatal treatment. *P < .01 compared with control animals of the same age and gender.
Discussion
The sexually dimorphic profiles of the hepatic P450 isoforms found herein are in agreement with previous reports, recently summarized byPampori and Shapiro (1996). That is, CYP2C12 is female specific, whereas CYP2C11, 2C13 and 3A2 are male specific. Hepatic CYP2A1, 2C6, and 2C7 are female predominant, and CYP3A1 is gender independent. Moreover, the observed developmental pattern of expression for each isoform was similar to earlier reports. CYP2C7 (Bandiera et al., 1986;Gonzalez et al., 1986), 2C11 (Morgan et al., 1985; Cresteil et al., 1986), 2C12 (Cresteil et al., 1986), and 2C13 (Bandiera et al., 1986) remain undetectable until puberty at ∼5 to 7 weeks of age, at which time they express their characteristic sexual dimorphisms. Similar to the sex-specific isoforms, hepatic CYP2C6 is undetectable during the neonatal period, but expression increases to near adult levels shortly before puberty, thereafter exhibiting the usual male:female ratio of 1:2 in adulthood (Waxman et al., 1985; Omiecinski et al., 1990). CYP2A1 (as measured at the catalytic testosterone 7α-hydroxylase level) and 3A2 are expressed during the neonatal period at near adult levels with no gender differences until puberty when CYP2A1 levels in the male liver decline by 50% (Gonzalez et al., 1986; Nagata et al., 1987) and CYP3A2 is no longer observed in female liver (Waxman et al., 1985;Shimada et al., 1989b). Although we were able to detect concentrations, albeit very low, of CYP3A1 mRNA and calculate percentage of comparisons at the various neonatal, pubertal, and adult time points, this P450 is an inducible form only poorly, if at all, expressed constitutively (Cresteil et al., 1986; Gemzik et al., 1992).
Generally, the preponderance of studies examining the inductive effects of phenobarbital on individual P450 isoforms have been conducted in adult rats. Only a very few studies have investigated the inductive responsiveness in the neonate (Cresteil et al., 1986; Omiecinski et al., 1990). In agreement with an earlier report (Cresteil et al., 1986), we found that CYP2C11 and 2C12 were not phenobarbital inducible during the neonatal period. In addition, we observed that both hepatic CYP2C7 and 2C13 were similarly unresponsive to the barbiturate during the first week of life. Because these isoforms are not expressed before puberty and are not responsive, or only slightly so, to the barbiturate in adulthood (Waxman et al., 1985; Cresteil et al., 1986), the refractory reaction in the neonate is understandable.
Similar to CYP2C7, 2C11, 2C12, and 2C13, CYP2C6 is unexpressed in the neonate and poorly responsive to the inductive effects of phenobarbital in adulthood (Waxman et al., 1985; Bandiera et al., 1986; Cresteil et al., 1986). Yet CYP2C6 is the only isoform of these five that was inducible during the first week of life and as previously reported (Omiecinski et al., 1990), during the last few days of gestation. Perhaps the somewhat earlier constitutive expression of CYP2C6 (i.e., prepubertal) compared with CYP2C7, 2C11, 2C12, and 2C13 (i.e., pubertal or postpubertal) indicates that the inductive mechanism responding to the barbiturate also may have developed earlier in life (i.e., perinatally).
We found that CYP3A1 and 3A2 as well as CYP2A1-dependent testosterone 7α-hydroxylase and CYP2A2-dependent testosterone 15α-hydroxylase were all expressed during the neonatal period and also were responsive to phenobarbital induction at this time. Accordingly, the cellular mechanisms regulating constitutive expression and phenobarbital inducibility appear to have developed concurrently.
By definition, phenobarbital induction of P450s is both a rapid and reversible reaction. That is, phenobarbital administration initiates an immediate (i.e., within hours) elevation in responsive hepatic P450s that decline to preinduction levels when the barbiturate is withdrawn (as observed in our neonates). Induction of the isoforms is dependent on the presence of phenobarbital. In theory, there is no limit to the number of times the process can be repeated. However, the reversibility of the inductive mechanism, so characteristic of the process, is compromised when the barbiturate is administered to the developing animal where the differentiating hepatic P450 system is vulnerable to irreversible imprinting defects.
Reports that neonatal exposure to phenobarbital results in a delayed, but permanent elevation in several drug-metabolizing enzymes is a contradiction to the expected rapid and transient response normally observed. Accordingly, neonatal administration of therapeutic-like doses of phenobarbital have been found to produce a postpubertal and permanent overexpression of such hepatic monooxygenases as aminopyrine demethylase and hexobarbital hydroxylase (Bagley and Hayes, 1985;Agrawal et al., 1995a). Previously, in an attempt to identify the P450 isoform(s) responsible for the permanent stimulation in hepatic drug-metabolizing enzyme activities, we examined the long-term effects of neonatal phenobarbital administration on expression levels of CYP2B1 and 2B2, the prototypical and most responsive isoforms to barbiturate induction (Agrawal and Shapiro, 1996a). After neonatal exposure to the barbiturate, we observed a rapid and profound elevation in transcript levels for both isoforms that subsequently declined but remained at concentrations 2-fold higher than controls through adulthood. Although the immediate neonatal inductive response was accompanied by a commensurate increase in CYP2B1 and 2B2 proteins and catalytic activities, the permanent 2-fold elevation in mRNAs was not translated into above-normal protein and activity levels, excluding CYP2B1 and 2B2 as the isoforms responsible for the phenobarbital imprinted overinduction of hepatic monooxygenases. In this regard, depending on several factors, such as dose, age, or gender, phenobarbital induction of CYP2B1 and 2B2 in mature rats can stimulate a significant elevation in transcript levels without an associated increase in proteins or catalytic activities (Agrawal and Shapiro, 1996b). This “uncoupling” of transcription and translation of CYP2B1 and 2B2 by both phenobarbital induction and imprinting is not unique to the CYP2B isoforms. Phenobarbital treatment to adult rats produces a substantial increase in CYP2C11 mRNA but a surprising decrease in protein levels (Shimada et al., 1989a). Similarly, we have found that CYP2C11 imprinting by the barbiturate results in a long-lasting overexpression of the transcript with no change in protein or CYP2C11-dependent testosterone 2α- or 16α-hydroxylases. Accordingly, the P450 isoform(s) responsible for the phenobarbital-imprinted overexpression of hepatic drug-metabolizing enzymes in both male and female rats should exhibit a persistent, postpubertal elevation in its mRNA, protein, and/or activity in both genders. Our results indicate that these requisites are fulfilled by CYP2C74, which unfortunately does not express a specific testosterone-metabolizing enzyme activity (Schenkman, 1992). Although CYP2C7 in both sexes is neither constitutively expressed nor phenobarbital inducible until the peripubertal period (Waxman et al., 1985; Cresteil et al., 1986), neonatal exposure to the barbiturate produced a “silent” programming defect resulting in a delayed but permanent postpubertal ∼30 to 50% overexpression in CYP2C7 mRNA and protein concentrations. Because CYP2C7 represents ∼20% of the total hepatic P450 in female rats and ∼6% in male rats (Bandiera and Dworschak, 1992), the ∼30 to 50% imprinted overexpression of the isoform could account, in part, for the observed 20 to 30% overexpression of hepatic hexobarbital hydroxylase (Agrawal et al., 1995a).
Unlike the other major P450 isoforms found in male and female rat liver, CYP2C7 has a unique requirement for retinoids (Westin et al., 1997) as well as physiological concentrations of thyroid (Ram and Waxman, 1990) and growth hormones (Pampori and Shapiro, 1999; Agrawal and Shapiro, 2000) for its full expression. In this regard, neonatally administered phenobarbital suppresses normal circulating concentrations of thyroxine (Fishman et al., 1982) and growth hormone (Kaneko et al., 1987; Agrawal and Shapiro, 1995a), the latter required for retinoid metabolism (Westin et al., 1997), which by some unknown mechanism could imprint the developing liver to express a permanent “rebound” overexpression of CYP2C7 in adulthood.
Although any clinical significance of our findings is uncertain, it has been considered (Shapiro, 1992), and we do know that in humans, peripartum exposure to barbiturates can have long-lasting, adverse effects on intelligence (Farwell et al., 1990, Reinisch et al., 1995), growth, and reproduction (Yaffe and Dorn, 1990; Dessens et al., 1994) as well as an enhanced susceptibility to cancers (Gold et al., 1978). Considering that millions of people have been exposed to barbiturates during development (Reinisch and Sanders, 1982), it is possible that any permanent elevation, albeit a modest ∼30 to 50% increase in the activity of a selective P450 isoform(s) over a life time, could compromise the effectiveness of drug therapies and alter the normal metabolism of endogenous substrates and environmental chemicals, consequently impacting, perhaps subtly, on the long-term health of an individual.
Acknowledgments
We appreciate the generosity of Drs. Marika Rönnholm, Agneta Mode, and Jan-Åke Gustafsson in supplying the antibody to rat CYP2C12 and Dr. Stelvio M. Bandiera in supplying the antibody to rat CYP2C7. Materials used to assay rat growth hormone were obtained through the National Hormone and Pituitary Program and Dr. A. F. Parlow. We also thank Alka Agrawal for excellent technical assistance.
Footnotes
-
Send reprint requests to: Bernard H. Shapiro, Laboratories of Biochemistry, School of Veterinary Medicine, University of Pennsylvania, 3800 Spruce St., Philadelphia, PA 19104-6048. E-mail:shapirob{at}vet.upenn.edu
-
↵1 This study was supported by National Institutes of Health Grants HD16358 and GM45758.
-
↵2 Current address: Department of Drug Metabolism, Merck Research Laboratories, P.O. Box 2000, RY80-A9, Rahway, NJ 07065-0900.
-
↵3 The terms sex-dependent, sex-predominant or -dominant, and sex-specific are often used indiscriminately. We use sex- or gender-dependent to imply that expression levels are dependent on the existence of gender; sex- or gender-predominant indicates expression levels, regardless of magnitude, are consistently greater in one gender; and sex- or gender-specific implies that expression is basically restricted to only one gender.
-
↵4 Although we have observed a persistent elevation of CYP2A2-dependent testosterone 15α-hydroxylase in male and female rats neonatally treated with phenobarbital, we have discounted the importance of any increased CYP2A2 in contributing significantly to the broadly overexpressed drug-metabolizing enzyme activities in the affected animals. In agreement with our finding of comparatively very low CYP2A2-testosterone 15α-hydroxylase activity in adult (150-day-old) male rat liver and virtually no activity in adult female liver (Fig. 3), CYP2A2 protein has been calculated to represent ∼1 to 2% of the total P450 in the male and basically nothing in the female (Thummel et al., 1988). Thus, even a permanent severalfold increase in the concentration of the isoform would make an almost irrelevant contribution, especially in the female, to the animals' hepatic monooxygenase activities.
- Abbreviation:
- CYP
- cytochrome P450
- Received December 28, 1999.
- Accepted February 17, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}