


Abstract
Flavonoids are constituents of fruits, vegetables, and plant-derived beverages, as well as components in herbal-containing dietary supplements. The objective of this investigation was to characterize the effect of flavonoids on P-glycoprotein (P-gp)-mediated cellular efflux and to determine the molecular mechanism(s) of the flavonoid-drug interaction. Studies were conducted in the sensitive and multidrug resistant human breast cancer cell lines MCF-7 and MDA435/LCC6 and examined the effects of the flavonoids biochanin A, morin, phloretin, and silymarin on daunomycin (DNM) accumulation and doxorubicin cytotoxicity. The potential mechanism(s) involved in the interaction was evaluated by determining flavonoid effects on 1) P-gp ATPase activity, 2) [3H]azidopine photoaffinity labeling of P-gp, and 3) cellular P-gp levels. The flavonoids increased [3H]DNM accumulation in P-gp positive cells, but not P-gp negative cells, and these effects were both flavonoid concentration- and P-gp expression level-dependent. Biochanin A and silymarin potentiated doxorubicin cytotoxicity in P-gp positive cells. Biochanin A and phloretin stimulated, whereas morin and silymarin inhibited P-gp ATPase activity, confirming that these flavonoids interact with P-gp. Morin and silymarin significantly inhibited [3H]azidopine photoaffinity labeling of P-gp, suggesting a direct interaction with P-gp substrate binding. A 24-h preincubation with all flavonoids, followed by flavonoid removal, did not alter cellular P-gp level in P-gp positive cells. In conclusion, biochanin A, morin, phloretin, and silymarin all inhibited P-gp-mediated cellular efflux and the mechanism of the interaction involved, at least in part, a direct interaction. The findings of this study indicate a potential for significant flavonoid-drug interactions with P-gp substrates.
Cancer chemotherapy has been of limited success because of the intrinsic or acquired resistance of cancer cells to a broad range of chemically and functionally distinct anticancer agents, a phenomenon termed multidrug resistance (MDR). The classical form of MDR involves the overexpression of drug efflux transporters such as P-glycoprotein (P-gp) (Juliano and Ling, 1976; Kartner et al., 1983) and multidrug resistance-associated protein 1 (MRP1) (Cole et al., 1992) in the cell membrane, which pump anticancer drugs out of the cells, resulting in low intracellular drug concentrations. P-gp is a 170- to 180-kDa plasma membrane protein encoded by the human MDR1 and MDR3 genes and the murinemdr1a, mdr1b, and mdr2 genes. P-gp belongs to the ATP-binding cassette transporter superfamily and is responsible for the efflux of a broad array of hydrophobic compounds, including a number of important chemotherapeutic agents such as vinca alkaloids, anthracyclines, epipodophyllotoxins, and taxol (Germann, 1996). Structure analysis based on the P-gp sequence predicts that P-gp consists of two homologous halves, each containing six transmembrane domains and a cytoplasmic nucleotide-binding domain (Gottesman and Pastan, 1993). P-gp acts as an efflux pump by exporting its substrate from the membrane or cell cytosol to the exterior of the cells, with ATP hydrolysis as the driving force (Gottesman and Pastan, 1993). Overexpression of this efflux protein has been associated with the clinical MDR phenotype and poor prognosis for many human cancers (Nooter and Sonneveld, 1994; List, 1996). Since the report that the calcium channel blocker verapamil could reverse resistance by inhibiting P-gp-mediated drug efflux (Tsuruo et al., 1981), a wide panel of substances have been tested both in vitro and in clinical trials for their ability to reverse MDR. Although a variety of agents, including calcium channel blockers, calmodulin antagonists, cyclosporins, quinolines, and their analogs have been found to be potent P-gp inhibitors in MDR cancer cells, the clinical results have been disappointing, due to the toxicities resulting from administration of the P-gp modulators and resulting from the pharmacokinetic interactions between the modulators and cytotoxic drugs (Volm, 1998).
Besides its role in conferring MDR, P-gp has also been found in many normal tissues, including the intestinal epithelium, blood-brain barrier, hepatocytes, and renal tubular cells, suggesting its important role in drug absorption, elimination, and distribution (Yu, 1999). It has been shown that the oral absorption and brain penetration of P-gp substrates are significantly lower in normal mice compared with mdr1a (−/−) mice (Schinkel, 1998), and oral bioavailability and brain penetration of P-gp substrates can be significantly enhanced by simultaneous administration of P-gp modulators (Mayer et al., 1997;Fromm et al., 1999). Clearance of a P-gp substrate due to biliary excretion and renal secretion can be also significantly decreased in the presence of a P-gp inhibitor (Song et al., 1999; Kiso et al., 2000).
Flavonoids are the most abundant polyphenolic compounds present in the human diet in fruits, vegetables, and plant-derived beverages such as tea and red wine. More than 4000 naturally occurring flavonoids have been described (Harborne, 1986). Certain plants and spices containing flavonoids represent traditional Eastern medicines, including the widely used Ginkgo biloba, and flavonoids are present in a plethora of dietary supplements. The daily intake of total flavonoids from the average U.S. diet is estimated to be 1 g, expressed as glycosides (Kuhnau, 1976), and could be higher when dietary supplements are taken. Flavonoids have long been associated with a variety of biochemical and pharmacological properties, including antioxidant, antiviral, anticarcinogenic, and anti-inflammatory activities (Middleton et al., 2000), believed to be beneficial to human health. In a recent review, Middleton et al. (2000) concluded “the flavonoids seem to be remarkably safe nutrients with a wide range of biochemical and pharmacological activities that strongly suggest their possible role as health-promoting, disease-preventing dietary supplements”.
The objectives of this investigation are 1) to determine the effects of some naturally occurring flavonoids, namely, biochanin A, morin, phloretin, and silymarin, on P-gp-mediated drug efflux; and 2) to gain an understanding of the potential mechanism(s) underlying these interactions. These flavonoids have been shown to be able to increase the accumulation of [3H]daunomycin ([3H]DNM) in P-gp-overexpressing MCF-7/ADR cells from our initial screening of 20 naturally occurring flavonoids (Tseng et al., 2001).
Materials and Methods
Materials.
Verapamil and the flavonoids were purchased from Sigma-Aldrich (St. Louis, MO). Silymarin is comprised of silybinin, silydianin, and silychristine, and the molar concentration was calculated based on the molecular weight of silybinin because this is the major flavonoid present. [3H]DNM) (14.4 Ci/mmol) was purchased from PerkinElmer Life Sciences (Boston, MA). P-gp negative control membrane and human P-gp membrane were purchased from BD Gentest (Woburn, MA). RPMI 1640 medium, fetal bovine serum, and phosphate-buffered saline (PBS) were from Invitrogen (Buffalo, NY). MCF-7/sensitive and MCF-7/ADR cells were gifts from Dr. Ralph Bernacki (Roswell Park Cancer Institute, Buffalo, NY). MDA435/LCC6 and MDA435/LCC6MDR cells were kindly provided by Dr. Robert Clarke (Georgetown University, Washington, DC). Both MCF-7 and MDA435/LCC6 cell lines are human breast cancer cell lines; MCF-7 is estrogen-dependent and MDA435/LCC6 is estrogen-independent. [3H]Azidopine and biodegradable counting scintillate were purchased from Amersham Biosciences Inc. (Piscataway, NJ).
Cell Culture
MCF-7 and MDA435/LCC6 cells (both parent and MDR subtype) were cultured in 75-cm2flasks with RPMI 1640 culture media supplemented with 10% fetal bovine serum at 37°C in a humidified atmosphere with 10% CO2/90% air. For MCF-7 cells, culture media also contained 100 units/ml penicillin and 100 μg/ml streptomycin. MCF-7 cells with passage number of 16 to 24 and MDA435/LCC6 (both wild type and MDR subtype) cells with passage number of 16 to 28 were used for experiments. A solution of 0.05% trypsin-EDTA was used to detach the cells. For the drug accumulation studies, cells were seeded onto 35-mm2 dishes at a density of approximately 5 × 105 cells/dish. Confluence was reached after 2 days.
Western Analysis of P-gp and MRP1.
Cells grown in 100 × 15-mm culture dishes were washed with PBS and harvested using a rubber policeman. Total cell lysates were prepared by adding the lysis buffer (20 mM Tris pH 7.5, 120 mM NaCl, 100 mM NaF, 1% Nonidet P-40, 200 μM sodium orthovanadate, 50 mM β-glycerolphosphate, 10 mM sodium pyrophosphate, 4 mM phenylmethylsulfonyl fluoride, 2 mM benzamidine, 10 μg/ml leupeptin, and 10 μg/ml aprotinin) to the harvested cells. The cells were kept on ice for 30 min. The soluble extracts were obtained by centrifuging the cell lysates at 13,000g for 20 min. The protein concentrations of the soluble extracts were obtained by bicinchoninic acid protein assay (Pierce Chemical, Rockford, IL). Proteins (30 μg) were electrophoresed on 7.5% SDS-polyacrylamide gels and electroblotted onto nitrocellulose membranes (Invitrogen, Grand Island, NY). Membranes were then blocked overnight at 4°C in Tris-buffered saline containing 0.2% (v/v) Tween 20 and 5% (w/v) fat-free dry milk (Bio-Rad, Hercules, CA), and then incubated first with primary antibody and then with secondary antibody at room temperature for 2 and 1.5 h, respectively. C219 (DAKO, Carpinteria, CA) and MRPm6 (Signet Laboratories, Dedham, MA) were used as primary antibodies to detect P-gp and MRP1, respectively. Anti-mouse IgG horseradish peroxidase (Amersham Biosciences, Inc.) was used as secondary antibody. As a loading control, some of the membranes were also incubated with anti-β-actin antibody (Alltech Associates, Deerfield, IL) and anti-mouse IgG to detect β-actin. After incubation with the antibodies, membranes were washed and detected with enhanced chemiluminescence detection reagent (Amersham Biosciences Inc.). 1D Image Analysis Software (Eastman Kodak, Rochester, NY) was use to analyze the Western blot results.
[3H]DNM Accumulation Studies.
The accumulation studies were performed after the cells reached 90% confluence in 35-mm2 dishes. Cells were washed twice with assay buffer (137 mM NaCl, 54 mM KCl, 2.8 mM CaCl2, 1.2 mM MgCl2, and 10 mM Hepes, pH 7.4) and then incubated with 0.05 μM [3H]DNM containing 50 μM flavonoids or vehicle (0.2% DMSO, used as control). Verapamil (100 μM) was added as a positive control. Preliminary studies demonstrated that steady-state conditions were achieved by 2 h for [3H]DNM accumulation in MCF-7/ADR cells. The accumulation was stopped by rinsing the cells four times with ice-cold buffer (137 mM NaCl, and 14 mM Tris base, pH 7.4). Cells were then solubilized using a solution of 0.3 N NaOH and 1% SDS, and aliquots were used to determine radioactivity by liquid scintillation counting (1900 CA, Tri-Carb liquid scintillation analyzer; PerkinElmer Life Sciences) and protein content using the bicinchoninic acid protein assay. Results were normalized for the protein content of the cells in each dish, and [3H]DNM accumulation level was expressed as percentage of accumulation in control group. For each sample, three or four separate experiments were performed and each experiment had triplicate measurements.
Cytotoxicity Studies.
Cytotoxicity studies were performed in 96-well plates. Cells (5 × 103) were seeded in each well of 96-well plates. After cell attachment (24-h incubation), culture medium in each well was replaced with fresh medium containing 0 to 300 μM doxorubicin as well as 100 μM flavonoids or vehicle (0.2% DMSO). Verapamil (100 μM) was used as a positive control. After incubation for another 24 h, the drug-containing medium was aspirated and cells washed three times with PBS buffer and fresh medium (without any drug) added. The incubation was then continued for an additional 24 h and cell growth in each well was determined by sulforhodamine B (SRB) assay (Skehan et al., 1990). The absorbance values (OD570) from the SRB assay indicate the cell number in each well of the 96-well plates. Growth inhibition by doxorubicin (IC50 value) either alone or with 100 μM flavonoids was obtained by fitting the fraction of cell growth (F) by the equation: F = 1 − (Imax · Cγ)/(IC50γ + Cγ) using WinNonlin (Pharsight, Mountain View, CA). The observed F values were calculated as the ratio of the cell growth [OD570 − OD570(300)] to the maximum cell growth [OD570(0) − OD570(300)]. OD570(0), OD570(300) are the absorbance values from cells treated with 0 and 300 μM doxorubicin, respectively. C is the concentration of doxorubicin. In each experiment, quadruplicate measurements were performed for each sample. The extent of potentiation of the doxorubicin cyotoxicity is expressed as the ratio of IC50 without flavonoids to that with flavonoids. Three to five separate experiments were performed.
Initial Uptake Rate Studies.
The experiments were performed in a similar manner as the [3H]DNM accumulation studies, but after the cells were washed with assay buffer, they were incubated with 50 μM flavonoids or vehicle (0.2% DMSO) for 1.5 h. Then 0.05 μM [3H]DNM was added and the incubation was continued for 7 min. The accumulation was then stopped and samples were processed as described in [3H]DNM accumulation studies. Preliminary studies demonstrated that the accumulation of [3H]DNM in MCF-7/sensitive cells is approximately linear within 10 min, and so the initial uptake rate was calculated as [3H]DNM accumulation per minute normalized by protein content. Results were expressed as percentage of the uptake rate of the control group. For each sample, three separate experiments were performed and each experiment had triplicate measurements.
P-gp ATPase Activity Assay.
P-gp ATPase activity was measured using human P-gp membrane and P-gp negative control membrane, following the manufacturer's instructions. Briefly, human P-gp membrane or P-gp negative control membrane (30 μg) was incubated at 37°C in 60 μl of Tris-MES buffer [50 mM (2-[N-morpholino] ethanesulfonic acid) hydrate, 2 mM EGTA, 2 mM dithiothreitol, 50 mM KCl, and 5 mM NaN3, with pH adjusted to 6.8 with Tris base) containing 100 μM flavonoids, solvent control (0.2% DMSO), and/or 20 μM verapamil. The ATPase reaction was initiated by the addition of 4 mM MgATP. The reaction was stopped by addition of 30 μl of 10% SDS solution and the amount of phosphate was determined immediately by adding 200 μl of detection solution (4 parts of 10% ascorbic acid, pH 5.0 and 1 part of 35 mM ammonium molybdate in 15 mM zinc acetate, pH 5.0) and incubating at 37°C for 20 min in the dark. The amount of phosphate was then determined by measuring the absorbance at 800-nm wavelength with phosphate buffer (pH 7.4) as standards. Preliminary studies using phosphate buffer standards demonstrated that 100 μM flavonoids or 20 μM verapamil, as used in these experiments, did not interfere with the phosphate assay. Phosphate released by ATPase reaction was estimated by the difference in the phosphate level between 0-min (ATPase reaction was stopped immediately with SDS solution) and 20-min incubation periods, and the ATPase activity was expressed as the rate of phosphate release per milligram of membrane protein. The difference between ATPase activity in the absence and presence of 100 μM orthovanadate (orthovanadate-sensitive ATPase activity) (Urbatsch et al., 1995) was determined.
[3H]Azidopine Photoaffinity Labeling.
Photoaffinity labeling of P-gp by [3H]azidopine was performed as described previously (Castro and Altenberg, 1997) with certain modifications. Briefly, 50 μg of human P-gp membrane or control membrane was incubated with 100 μM flavonoids, or solvent control (0.2% DMSO) for 30 min; [3H]azidopine was then added to a final concentration of 0.5 μM and incubated in the dark for 60 min. Then the samples were irradiated with UV light at wavelength of 254 to 320 nm (Ultra-Violet Products, Inc., Upland, CA) for 20 min at a distance of 8 cm. Samples were then electrophoresed on a 7.5% SDS-polyacrylamide gel electrophoresis. Gels were fixed in 10% acetic acid/40% methanol for 1 h and then incubated with AMPLIFY (Amersham Biosciences, Inc.), and dried. Autoradiography was performed at −70°C for a week. Vinblastine (100 μM) was included as a positive control. The signal intensity was quantified using OptiQuant software (PerkinElmer Life Sciences) and expressed as percentage of intensity of the solvent control.
Preincubation Studies.
MCF-7/sensitive and MCF-7/ADR cells were grown in 35-mm2 dishes. After cells reached 80 to 90% confluence, the cell media were replaced with fresh medium containing flavonoids (50 μM) or vehicle (0.2% DMSO), and the incubation (37°C) was continued for another 24 h. Cells were then washed twice with assay buffer and the 2-h accumulation of [3H]DNM was determined as described for the [3H]DNM accumulation studies. Verapamil was used as a positive control and added during the 2-h accumulation period instead of during the preincubation period. Two or three separate experiments were performed and each experiment was performed in triplicate.
Statistical Analysis.
The differences between the mean values were analyzed for significance using one-way analysis of variance followed by Dunnett's test. p values <0.05 were considered statistically significant.
Results
P-gp and MRP1 Expression Levels.
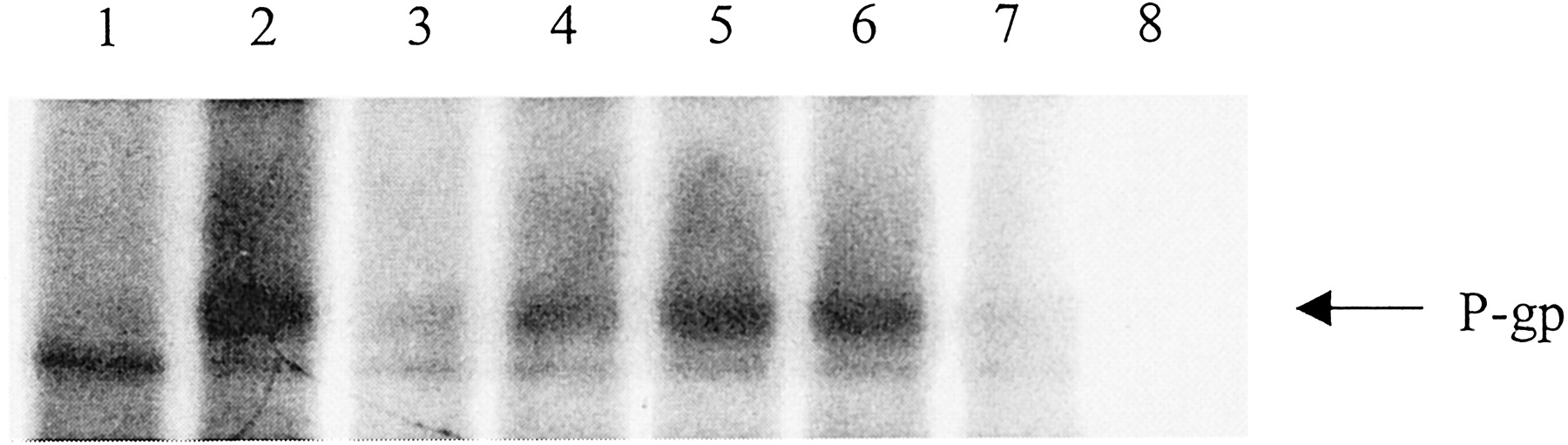
To investigate effects of flavonoids on P-gp-mediated cellular efflux, we first characterized the cellular P-gp levels in the cells used in this study using Western blot analysis. As shown in Fig. 1, the rank order of cellular P-gp level (lowest to highest) is MCF-7/sensitive, MDA435/LCC6WT, MCF-7/ADR, and MDA435/LCC6MDR1 cells. MCF-7/sensitive cells have no detectable P-gp expression, which is consistent with previous observations (Fairchild et al., 1990). Overexpression of P-gp in MCF-7/ADR and MDA435/LCC6MDR1 cells have been well established and the cells are used extensively as model MDR cells overexpressing P-gp (Fairchild et al., 1990; Leonessa et al., 1996). MDA435/LCC6WT cells have moderate cellular P-gp. No significant expression of MRP1 was found in any of the cell lines. This was determined with primary antibody MRPm6 and using H69/AR (Cole et al., 1992) and H69 cells as positive and negative controls, respectively (data not shown).

Western blot analysis of P-gp expression in MCF-7 and MDA435/LCC6 cells. Cellular P-gp level in MCF-7/sensitive (1), MCF-7/ADR (2), MDA435/LCC6 (3), and MDA435/LCC6MDR1 (4) cells were examined by Western blot analysis, as described under Materials and Methods. P-gp was detected with the primary antibody C219. β-Actin was used to confirm equal protein loading. Two separate experiments were conducted and similar results were obtained.
Effects of Flavonoids on [3H]DNM Accumulation.
After characterization of the cellular P-gp levels, the 2-h accumulation of [3H]DNM was determined in these cells in the presence or absence of flavonoids (50 μM). Verapamil (100 μM), a well characterized P-gp inhibitor, was used as a positive control. The results are shown in Fig. 2. The [3H]DNM accumulation in MCF-7/sensitive cells (Fig. 2A), which has no detectable P-gp expression, was not significantly changed by any flavonoids or verapamil. In MDA435/LCC6WT cells (Fig. 2C), which have moderate P-gp expression, and MCF-7/ADR (Fig. 2B) and MDA435/LCC6MDR1 (Fig. 2D) cells, both of which have substantial P-gp expression, [3H]DNM accumulation is significantly increased in the presence of all the tested flavonoids and verapamil (except for the [3H]DNM accumulation in MDA435/LCCMDR1 cells in the presence of phloretin, which was 277 ± 93% of the control, but not statistically significant). In addition, the increase in [3H]DNM accumulation in the presence of both flavonoids and verapamil was qualitatively correlated with the cellular P-gp levels in these cells (Fig. 2), with higher effects in cells expressing higher levels of P-gp. To further characterize the effects of flavonoids on the accumulation of [3H]DNM in P-gp positive cells, we also investigated [3H]DNM accumulation in MDA435/LCC6MDR1 cells in the presence of different concentrations (5–100 μM) of flavonoids. [3H]DNM accumulation was found to be flavonoid concentration-dependent. The enhancement of [3H]DNM accumulation increased with increasing flavonoid concentration and significant changes were observed after treatment with biochanin A (502 ± 16%, p < 0.001) and morin (604 ± 32%, p < 0.001) at 30 μM concentrations and phloretin (431 ± 28%, p< 0.001) and silymarin (445 ± 12%, p < 0.001) at 50 μM concentrations (data not shown).

[3H]DNM 2-h accumulation in cells expressing different levels of P-gp. The 2-h accumulation of [3H]DNM in MCF-7/sensitive (A), MCF-7/ADR (B), MDA435/LCC6 (C), and MDA435/LCC6MDR1 (D) cells in the presence or absence (control) of 50 μM flavonoids was investigated as described under Materials and Methods. Verapamil (100 μM) was included as a positive control. Data are expressed as mean ± S.D., n = 9 to 12. CON, control; BA, biochanin A; MO, morin; PH, phloretin; SI, silymarin; and VER, verapamil.
Effects of Flavonoids on Doxorubicin Cytotoxicity.
The effects of flavonoids on doxorubicin cytotoxicity in MDA435/LCC6MDR1 and MDA435/LCC6WT cells were investigated. Verapamil (100 μM) was used as a positive control. As shown in Fig. 3 and Table1, verapamil significantly enhanced doxorubicin cytotoxicity in both MDA435/LCC6MDR1 (p < 0.05) and MDA435/LCC6WT cells (p < 0.001) and the extent of cytotoxicity potentiation was related to the level of P-gp expression, being more pronounced in MDA435/LCC6MDR1 cells (IC50 ratio = 70.23) compared with that in MDA435/LCC6WT cells (IC50 ratio = 7.76). Biochanin A and silymarin enhanced doxorubicin cytotoxicity in MDA435/LCC6MDR1 cells (Fig. 3A; Table 1) [IC50ratio = 37.83 (p > 0.05) and 3.45 (p > 0.05), respectively], although the effects were not statistically significant when assessed by one-way analysis of variance followed by Dunnett's test (p < 0.01 andp < 0.05, respectively, if assessed by Student'st test). The trend of doxorubicin cytotoxicity enhancement by biochanin A and silymarin was consistent with the previous observation that both flavonoids can inhibit P-gp-mediated drug efflux and that biochanin A is more effective than silymarin in doing so. Morin and phloretin, however, were not able to potentiate doxorubicin cytotoxicity; instead, there was a trend toward decreased doxorubicin cytotoxicity (IC50 ratio = 0.75 and 0.53, respectively), although the effects were not statistically significant. Biochanin A significantly enhanced doxorubicin cytotoxicity in MDA435/LCC6WT cells (IC50 ratio = 10.30,p < 0.05), although the extent of cytotoxicity potentiation was less than that in MDA435/LCC6MDR1 cells (Fig. 3B; Table 1). As shown in Fig. 4, the effects of biochanin A in MDA435/LCC6MDR1 cells were concentration-dependent.

Effects of flavonoids on doxorubicin cytotoxicity in MDA435LCC6 cells. Doxorubicin cytotoxicity was investigated in MDA435/LCC6MDR1 (A) and MDA435/LCC6 (B) cells in the presence of 100 μM flavonoids or vehicle (0.2% DMSO), as described underMaterials and Methods. Verapamil (100 μM) was included as a positive control. Presented is a typical experiment and the data are expressed as mean ± S.D. Three to five separate experiments were performed and each experiment had quadruplicate measurements. ○, MDA435/LCC6; ▵, MDA435/LCC6 + 100 μM biochanin A; ▿, MDA435/LCC6 + 100 μM verapamil; ●, MDA435LCC6/MDR; ▪, MDA435/LCC6MDR1 + 100 μM phloretin; ▾, MDA435/LCC6MDR1 + 100 μM verapamil; ▴, MDA435/LCC6MDR1 + 100 μM biochanin A; ♦, MDA435/LCC6MDR1 + 100 μM morin;
 , MDA435/LCC6MDR1 + 100 μM silymarin.
, MDA435/LCC6MDR1 + 100 μM silymarin.
Effect of flavonoids (100 μM) on doxorubicin cytotoxicity in MDA435/LCC6 cells
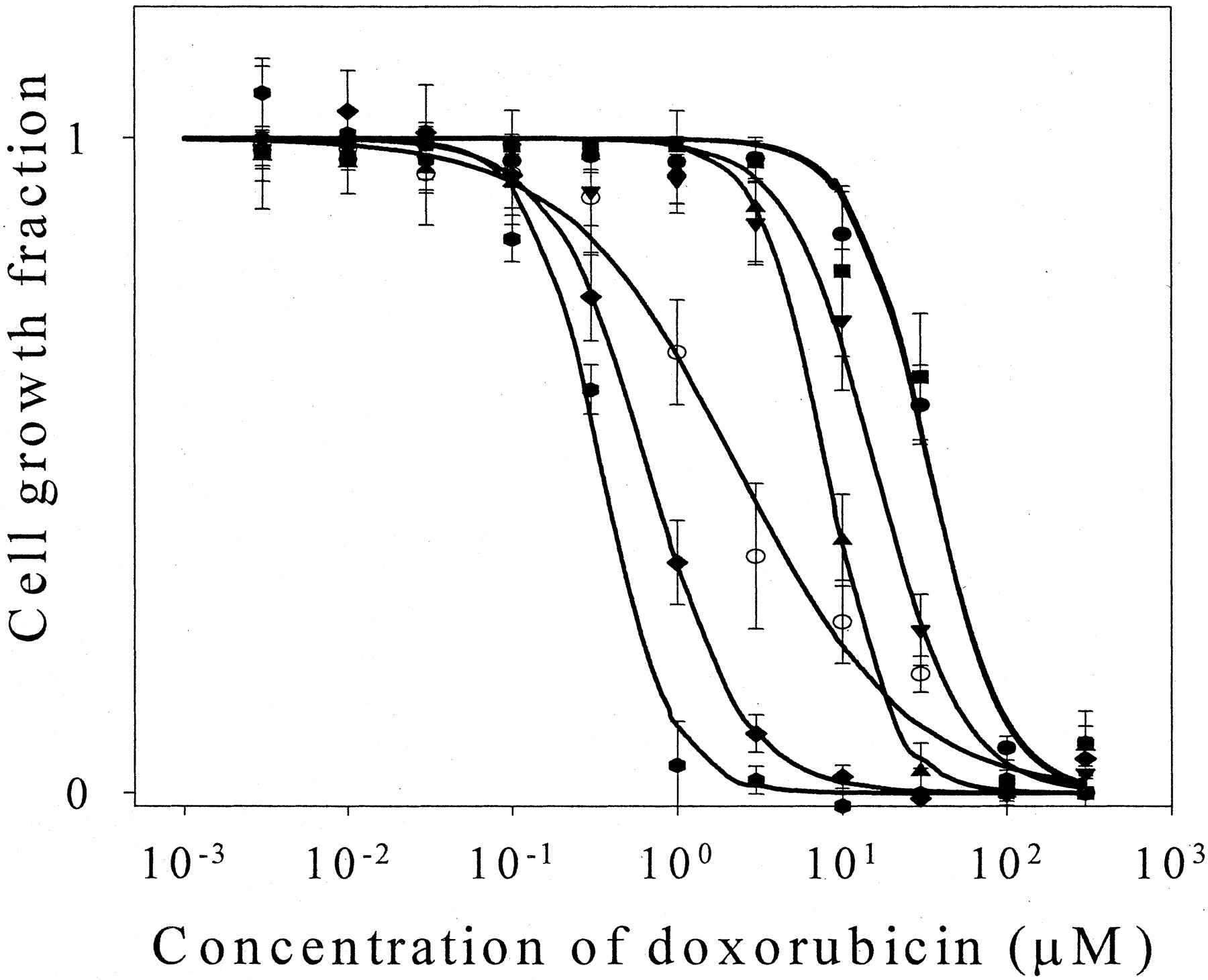
Concentration-dependent effects of biochanin A on doxorubicin cytotoxicity in MDA435/LCC6MDR1 cells. The cytotoxicity of doxorubicin in MDA435/LCC6MDR1 cells in the presence of different concentrations of biochanin A (10–100 μM) was investigated. The cytotoxicity study was performed as described under Materials and Methods. Verapamil (100 μM) was included as a positive control. Data are expressed as mean ± S.D., n= 4 wells. ○, MDA435/LCC6; ●, MDA435/LCC6MDR1; ▪, MDA435/LCC6MDR1 + 10 μM biochanin A; ▾, MDA435/LCC6MDR1 + 30 μM biochanin A; ▴, MDA435/LCC6MDR1 + 50 μM biochanin A; ♦, MDA435/LCC6MDR1 + 100 μM biochanin A; and
 , MDA435/LCC6MDR1 + 100 μM verapamil.
, MDA435/LCC6MDR1 + 100 μM verapamil.
Effects of Flavonoids on [3H]DNM Initial Uptake Rate in MCF-7/sensitive Cells.
To characterize the effects of flavonoids on passive membrane permeability, we investigated the effects of flavonoids (50 μM) on the initial uptake rate of [3H]DNM in P-gp negative cells (MCF-7/sensitive), which is a measure of DNM passive membrane permeability, assuming the uptake process in drug-sensitive cells is solely a diffusion process. None of the tested flavonoids significantly increased the initial uptake rate of [3H]DNM in MCF-7/sensitive cells, suggesting that none of these compounds increased DNM passive membrane permeability (data not shown). Instead, morin, phloretin, and silymarin slightly decreased the initial uptake rate of [3H]DNM in MCF-7/sensitive cells (82 ± 17, 82 ± 17, and 81 ± 15%, respectively, withp < 0.01 for all of these compounds).
Effects of Flavonoids on P-gp Expression after a 2-h Incubation.
The effects of flavonoids on cellular P-gp expression in MDA435/LCC6MDR1cells after a 2-h incubation period, as used in the [3H]DNM accumulation studies, were investigated. Cells grown in 35-mm2 dishes were washed and then incubated with the assay buffer used in the [3H]DNM accumulation studies containing 50 μM flavonoids or vehicle (0.2% DMSO) for 2 h. The cellular P-gp level of these cells was then examined by Western blot analysis. As shown in Fig. 5, none of the tested flavonoids significantly changed the cellular P-gp level in MDA435/LCC6MDR1 cells after a 2-h incubation, and the relative P-gp expression levels after biochanin A, morin, phloretin, and silymarin treatment were 104.5 ± 33.7, 102.7 ± 22.0, 110.4 ± 18.4, and 95.7 ± 10.2% of the control, respectively.

Effects of flavonoids on P-gp expression in MDA435/LCC6MDR1 cells after a 2-h incubation. Cells grown in 35-mm2 dishes were washed and incubated with the assay buffer used for [3H]DNM accumulation study containing 50 μM flavonoids or vehicle (0.2% DMSO). After incubation for 2 h, cells were harvested and P-gp expression level was analyzed by Western blot analysis as described under Materials and Methods. The P-gp expression level is expressed as percentage of control (vehicle treatment). Data are mean ± S.D. from four separate experiments. MCF-7/sensitive (P-gp negative control) (1); control (vehicle), 100% (2); biochanin A, 104.5 ± 33.7% (3); morin, 102.7 ± 22.0% (4); phloretin, 110.4 ± 18.4% (5); and silymarin, 95.7 ± 10.2% (6).
Effects of Flavonoids on P-gp ATPase Activity.
To further understand the interactions between flavonoids and P-gp, we examined the effects of flavonoids (100 μM) on both P-gp constitutive ATPase activity and verapamil (20 μM)-induced ATPase activity. As shown in Table 2, the presence of verapamil (20 μM) and flavonoids (100 μM) had no significant effects on orthovanadate-sensitive ATPase activity of the P-gp negative control membrane; however, the orthovanadate-sensitive ATPase activity of the P-gp membrane was significantly increased by both verapamil (a well known P-gp ATPase stimulator, p < 0.001) and biochanin A (p < 0.001) from 5.16 ± 2.37 to 31.7 ± 5.94 and 18.73 ± 3.28 nmol/min/mg, respectively, but not changed significantly by morin, phloretin, and silymarin. This observation suggested that biochanin A and verapamil can stimulate P-gp constitutive ATPase activity, but morin, phloretin, and silymarin are not effective. We also investigated the effects of flavonoids (100 μM) on the P-gp ATPase activity induced by verapamil (20 μM) by incubating the P-gp membrane in the assay system with both flavonoids (100 μM) and verapamil (20 μM). As shown in Table 2, the orthovanadate-sensitive ATPase activity of the P-gp membrane was significantly stimulated by 20 μM verapamil (from 3.40 ± 1.97 to 32.60 ± 6.83 nmol/min/mg, p < 0.001), consistent with the above-mentioned observation. In the copresence of biochanin A, orthovanadate-sensitive ATPase activity is further stimulated (from 32.60 ± 6.83 to 61.04 ± 5.30 nmol/min/mg,p < 0.01). In the copresence of phloretin, orthovanadate-sensitive ATPase activity was also significantly increased (from 32.60 ± 6.83 to 52.74 ± 8.91 nmol/min/mg,p < 0.05), although phloretin was not able to stimulate the P-gp constitutive ATPase activity (Table 2). In the copresence of silymarin, the verapamil-induced P-gp ATPase activity was significantly decreased (from 32.60 ± 6.83 to 1.97 ± 6.44 nmol/min/mg, p < 0.01). In addition, it seems that silymarin (100 μM) was able to completely inhibit verapamil-induced P-gp ATPase activity because in the copresence of verapamil and silymarin, the orthovanadate-sensitive ATPase activity (1.97 ± 6.44 nmol/min/mg) was not significantly different from the P-gp constitutive ATPase activity (3.40 ± 1.97 nmol/min/mg). Morin (100 μM) also decreased verapamil-induced P-gp ATPase activity (from 32.60 ± 6.83 to 14.72 ± 6.27 nmol/min/mg, p> 0.05), although the effect was not statistically significant.
Effects of flavonoids (100 μM) on P-gp ATPase activity
[3H]Azidopine Photoaffinity Labeling.
To understand the interaction of flavonoids with P-gp and its substrate binding, we investigated [3H]azidopine photoaffinity labeling of P-gp in the presence of flavonoids (100 μM) using human P-gp membrane and P-gp negative control membrane. Vinblastine (100 μM) was included as a positive control. As shown in Fig. 6, vinblastine inhibited [3H]azidopine photoaffinity labeling of P-gp by 60.0 ± 2.7% (p < 0.001), similar to previously reported results (Castro and Altenberg, 1997), in which vinblastine (100 μM) inhibited [3H]azidopine photoaffinity labeling of P-gp by 75 ± 4%. All the tested flavonoids inhibited [3H]azidopine photoaffinity labeling of P-gp but to different extents; the effect of biochanin A and phloretin (inhibited 23.2 ± 6.3 and 30.2 ± 11.6%, respectively) was not statistically significant. The effect of morin was moderate but statistically significant (inhibited 37.9 ± 28.4%, p < 0.05), which might be comparable with that reported for genistein, which at a higher concentration (200 μM), inhibited [3H]azidopine photoaffinity labeling of P-gp by 42 ± 15% (Castro and Altenberg, 1997). The effect of silymarin was more substantial (61.2 ± 20.7%,p < 0.001) and comparable with that of vinblastine.

[3H]Azidopine photoaffinity labeling of P-gp [3H]azidopine photoaffinity labeling of P-gp in the presence of flavonoids was performed as described underMaterials and Methods. Vinblastine (100 μM) was used as a positive control. Three separate experiments were performed and the image is from one representative experiment. P-gp negative membrane (1); P-gp membrane (2); P-gp membrane with vinblastine, biochanin A, morin, phloretin, and silymarin, respectively (3–7); and P-gp membrane, but UV irradiation was done before instead of after [3H]azidopine incubation (8). The [3H]azidopine photoaffinity labeling of P-gp in the presence of flavonoids (100 μM) and vinblastine is biochanin A, 76.8 ± 6.3%, p > 0.05; morin, 62.1 ± 28.4%, p < 0.05; phloretin, 69.8 ± 11.6%,p > 0.05; silymarin, 38.8 ± 20.7%,p < 0.001; and vinblastine, 39.9 ± 2.7% of the control, p < 0.001.
Twenty-Four-Hour Preincubation Studies.
To investigate the effects of a longer incubation with flavonoids on P-gp-mediated efflux, we investigated the 2-h accumulation of [3H]DNM in both P-gp negative (MCF-7/sensitive) and P-gp positive (MCF-7/ADR) cells after pretreatment of the cells with 50 μM flavonoids for 24 h. Verapamil was included as a positive control and was added with [3H]DNM for the accumulation study (2-h incubation period). As shown in Fig. 7A, [3H]DNM accumulation in MCF-7/sensitive cells was not changed significantly by the pretreatment with any of the flavonoids. In MCF-7/ADR cells (Fig. 7B), [3H]DNM accumulation was significantly increased by a 24-h pretreatment with 50 μM silymarin, but the change (38 ± 5%, p < 0.001) was moderate. [3H]DNM accumulation was not changed significantly by pretreatment of cells with biochanin A, morin, or phloretin. As shown in Fig. 8, none of the tested flavonoids significantly changed cellular P-gp level in MCF-7/ADR cells after a 24-h pretreatment with 50 μM of these flavonoids, and the cellular P-gp level after biochanin A, morin, phloretin, and silymarin pretreatment was 106 ± 27, 83 ± 29, 83 ± 34, and 97 ± 11%, respectively. Data are expressed as mean ± S.D. of five to six separate experiments.
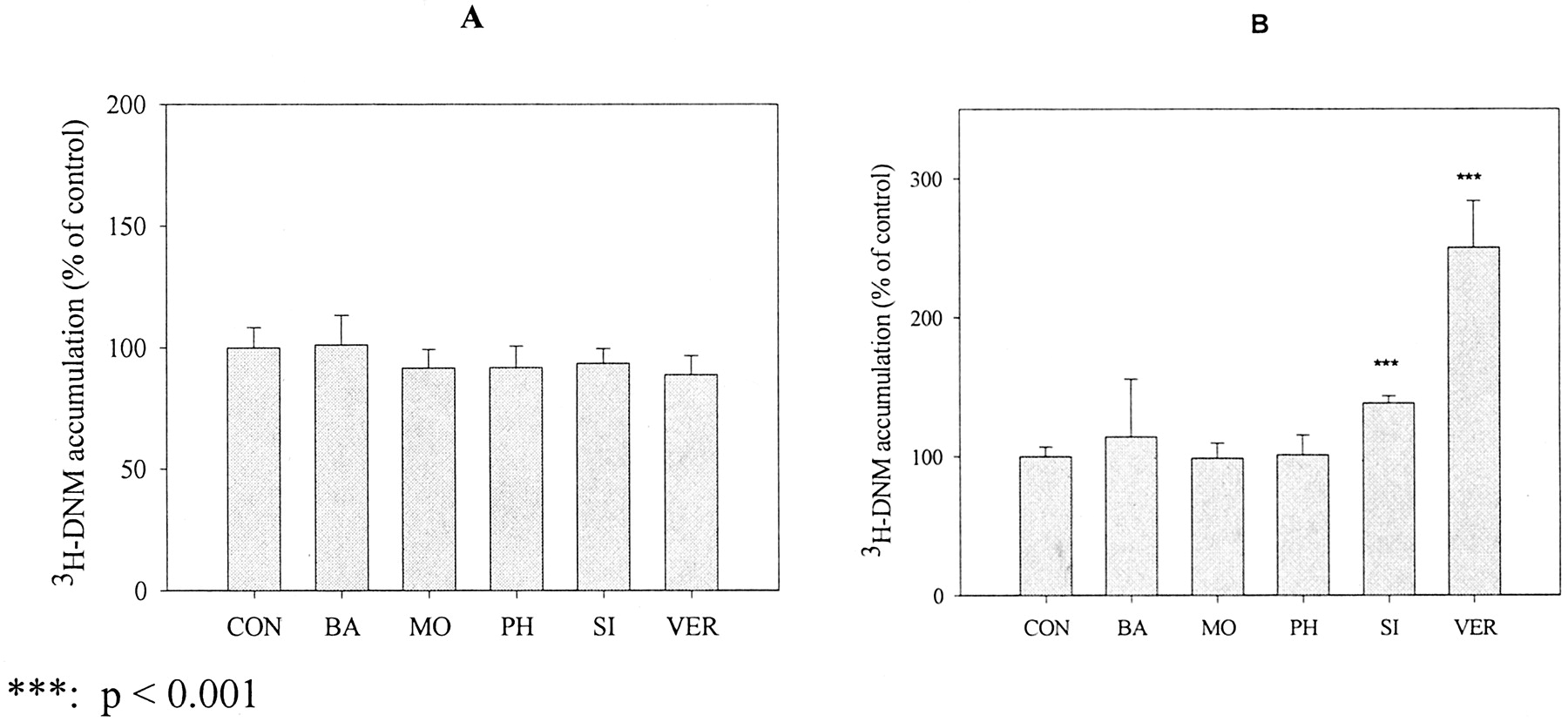
[3H]DNM accumulation in MCF-7 cells preincubated with flavonoids for 24 h. [3H]DNM accumulation was performed in MCF-7/sensitive (A) and MCF-7/ADR (B) cells after the cells were treated with 50 μM flavonoids for 24 h. The residual flavonoids were removed before the accumulation study. Verapamil was included as a positive control and was added with [3H]DNM for the accumulation study. Data are expressed as mean ± S.E., n = 6 or 9.

Western blot analysis of P-gp expression in MCF-7/ADR cells after a 24-h preincubation with flavonoids. MCF-7/ADR cells were treated with flavonoids or vehicle (0.2% DMSO) for 24 h. The P-gp expression level in these cells was then examined by Western blot analysis, as described under Materials and Methods. Presented are the results from a typical experiment and five to six separate experiments were performed. Data are expressed as mean ± S.D. MCF-7/sensitive (1); MCF-7/ADR (2); MCF-7/ADR treated with biochanin A (106 ± 27%) (3); with morin (83 ± 29%) (4); with phloretin (83 ± 34%) (5); and with silymarin (97 ± 11%) (6).
Discussion
Flavonoids are present in diet and in a variety of dietary supplements such as the widely used Ginkgo biloba, milk thistle, and soy isoflavones. With the burgeoning interest in alternative medicine, herbal products are ingested by about 10% or more of the general population and 30 to 70% of individuals with specific disease states (Duggan et al., 2001; Ni et al., 2002). Because dietary supplements are not classified as drugs and do not require Food and Drug Administration approval to be marketed, potential drug interactions are not extensively evaluated. However, there is increasing evidence indicating that use of herbal products could result in clinically important drug interactions. For example, it has been shown that St. John' wort reduced the area under the plasma concentration-time curve of indinavir by 57% (Piscitelli et al., 2000) and garlic supplements reduced the area under the plasma concentration-time curve of saquinavir by 51% (Piscitelli et al., 2002) in human subjects and the proposed mechanism(s) include induction of drug-metabolizing enzyme (CYP3A4) and/or P-gp (Durr et al., 2000).
From an initial screening of 20 naturally occurring flavonoids, we found that biochanin A, morin, phloretin, and silymarin can substantially increase the accumulation of [3H]DNM in MCF-7/ADR cells (Tseng et al., 2001). Thus, we decided to further study these flavonoids in four different breast cancer cell lines with different levels of P-gp expression. Although P-gp expression in MDA435/LCC6WT cells was not detected in a previous study (Leonessa et al., 1996) using Ab-1 polyclonal anti-gp170 antibody, we observed moderate P-gp expression in this cell line using the C219 antibody. The expression of P-gp in MDA435/LCC6WT cells was also consistent with our findings in the [3H]DNM accumulation and doxorubicin cytotoxicity studies, in which verapamil, a well known P-gp inhibitor, significantly increased [3H]DNM accumulation and potentiated doxorubicin cytotoxicity in these cells. The differences between our observation and that of Leonessa et al. (1996)may be due to the different antibodies and experimental conditions used. The flavonoids biochanin A, morin, phloretin, and silymarin were able to increase [3H]DNM accumulation in all the P-gp positive cells (MCF-7/ADR, MDA435/LCC6WT, and MDA435/LCCMDR1) but not in P-gp negative cells (MCF-7/sensitive), and the increase of DNM accumulation by these flavonoids was shown to be both flavonoid concentration-dependent and P-gp expression level-dependent, strongly suggesting that these flavonoids inhibited P-gp-mediated cellular efflux. The increase of DNM accumulation by 50 μM biochanin A or morin in all the P-gp positive cells is comparable with that with 100 μM verapamil, indicating that biochanin A and morin are fairly potent P-gp inhibitors. The possibility that the increase of DNM accumulation by these flavonoids may be due to modulation of MRP1, another drug efflux transporter, instead of P-gp, can be excluded by the following evidence. First, we were unable to detect MRP1 in all the cell lines with Western blot analysis using the MRPm6 antibody, indicating MRP1 expression in these cell lines must be very low, if present at all. In a previous study, Benderra et al. (2000) detected some expression of MRP1 in MCF-7/ADR cells by flow cytometry analysis using the same antibody; however, the expression level must be low enough to be functionally insignificant compared with P-gp because in the same study, it was shown that treatment of MCF-7/ADR cells with buthionine sulfoximine, a GSH biosynthesis inhibitor, had no effects on daunorubicin cytotoxicity in these cells; however, buthionine sulfoximine completely sensitized MCF-7/VP cells, which express relatively high levels of MRP1, to daunorubicin (Benderra et al., 2000). Second, the increase of DNM by these flavonoids has been shown to positively correlate with the cellular P-gp level of the cells. It seems likely that the cellular efflux of DNM by MRP1 in these cells is insignificant compared with that of P-gp.
In the cytotoxicity study, biochanin A and silymarin increased doxorubicin cytotoxicity in MDA435/LCC6MDR1 cells, consistent with their inhibitory effects on P-gp-mediated efflux. Biochanin A was also able to enhance doxorubicin cytotoxicity in MDA435/LCC6WT cells, but the effect was less than that in MDA435/LCC6MDR1 cells, suggesting that enhancement of doxorubicin cytotoxicity by biochanin A is mediated through P-gp inhibition. Phloretin and morin, however, were not able to potentiate doxorubicin cytotoxicity, although they were shown to inhibit P-gp-mediated efflux. A number of explanations could explain this inconsistency, although the exact reason is currently unknown. One possible explanation could be related to the stability of the various flavonoids. We found that biochanin A was stable when incubated with MDA435/LCC6MDR1 cells for 24 h (data not shown). However, the stability of morin and phloretin in culture media and in incubations with MDA435/LCC6MDR1 cells is not known. Drug accumulation studies were performed over a relatively short period of time (2 h) but the cytotoxicity studies involved 24-h drug incubations. Therefore, it is possible that phloretin and morin are degraded or metabolized by the cells during a 24-h incubation and thus had no effect on doxorubicin cytotoxicity.
To gain some understanding of the underlying mechanism(s) by which flavonoids alter P-gp-mediated cellular efflux, we examined the effects on 1) passive membrane permeability, 2) P-gp expression, 3) P-gp ATPase activity, and 4) direct binding using the photoaffinity label [3H]azidopine. An increase in passive membrane permeability could lead to an increase in DNM steady-state accumulation in P-gp positive cells but not P-gp negative cells. In P-gp negative cells, [3H]DNM accumulation at equilibrium will not be altered by the increased passive membrane permeability because the intracellular free DNM concentration will be always equal to the extracellular free DNM concentration at equilibrium, assuming the uptake of DNM in P-gp negative cells is solely a diffusion process. However, in P-gp positive cells, this is not true: the increased passive membrane permeability will result in an increased steady-state accumulation of [3H]DNM. A number of agents, including some flavonoids and P-gp modulators (Drori et al., 1995;Arora et al., 2000), have been shown to be able to change membrane lipid packing order and thus change membrane fluidity or permeability. So, it is at least theoretically possible that the observed effects of flavonoids on [3H]DNM accumulation may be due to their nonspecific interaction with the cell membrane, resulting in an increased passive membrane permeability, instead of a specific inhibition of P-gp. To exclude this possibility, we investigated the effects of flavonoids on the initial uptake rate of [3H]DNM in P-gp negative cells (MCF-7/sensitive). The uptake of anthracyclines in drug-sensitive cells, such as MCF-7/sensitive cells, can be described adequately by passive diffusion model (Frezard and Garnier-Suillerot, 1991), and so the initial uptake rate of [3H]DNM in MCF-7/sensitive cells represents a measure of passive membrane permeability. None of the tested flavonoids increased DNM initial uptake rate, indicating that the observed effects of these flavonoids are not the result of an increased passive membrane permeability.
An increase in DNM accumulation in P-gp positive cells could be due to a flavonoid-induced decrease in P-gp expression. P-gp expression was examined in MDA435/LCC6MDR1 cells after a 2-h incubation. Our data indicate that none of the tested flavonoids significantly changed the P-gp expression in these cells, suggesting that the observed enhancement of DNM accumulation by flavonoids is not due to their reduction of P-gp expression.
P-gp has been demonstrated to be an ATPase (al-Shawi and Senior, 1993), and many P-gp substrates and modulators have been shown to interact with P-gp ATPase activity (both stimulation and inhibition). For example, the P-gp inhibitor verapamil has been reported to be one of the best stimulators of P-gp ATPase (Shapiro and Ling, 1994, 1997a). Flavonoids have been reported to modulate P-gp by interacting bifunctionally with the vicinal ATP-binding site and the steroid-binding site (Conseil et al., 1998). Thus, one potential mechanism responsible for the flavonoid inhibition of P-gp-mediated efflux may be the inhibition of P-gp ATPase by binding to the ATP-binding site. In this study, we demonstrated that silymarin significantly inhibited P-gp ATPase activity induced by verapamil; however, biochanin A and phloretin stimulated P-gp ATPase activity. These findings indicate that the interactions of these flavonoids with P-gp may differ. Under our experimental conditions, we could not detect the inhibitory effects of morin and silymarin on P-gp constitutive ATPase activity, indicating that they may be not effective in modulating P-gp constitutive ATPase activity. However, it is also likely that this lack of effect on P-gp constitutive ATPase activity may be due to the limited sensitivity of the assay system, because the P-gp constitutive ATPase activity is low and further inhibition can be difficult to detect due to the experimental variability. Therefore, it is possible that the flavonoids silymarin and probably morin, which inhibit P-gp ATPase activity induced by verapamil, may bind to the ATP-binding site and vicinal steroid-binding site and thus prevent ATP binding. However, it is also possible, especially for silymarin, that these flavonoids bind to the P-gp substrate-binding site and thus prevent binding of verapamil and inhibit verapamil-induced ATPase activity. This latter hypothesis is supported by the results of our photoaffinity labeling studies, in which silymarin significantly inhibited [3H]azidopine labeling of P-gp (inhibited 61.2 ± 20.7%, p < 0.001). It is interesting that phloretin increased P-gp ATPase activity in the presence of verapamil; however, it had no effect on P-gp constitutive ATPase activity (in the absence of verapamil). The underlying mechanism is not clear at this time, but one explanation could be that binding of phloretin itself to P-gp may be not enough to affect P-gp ATPase activity, but phloretin binding could change the protein conformation and enhance the ATPase stimulatory effects upon the binding of verapamil. It has been proposed that P-gp possesses at least two substrate-binding sites and an allosteric site (Shapiro and Ling, 1997b; Shapiro et al., 1999). It is therefore possible that phloretin could bind to the allosteric site or other sites and enhance the ability of verapamil to stimulate ATPase activity. Biochanin A effects differed from those of phloretin in that it stimulated P-gp ATPase activity both in the presence and in the absence of verapamil. In addition, the ATPase activity in the presence of both biochanin A and verapamil was shown not to be significantly different from the sum of the ATPase activities in the presence of biochanin A alone and verapamil alone. These data suggested that biochanin A itself can stimulate P-gp ATPase, and the stimulatory effects of verapamil and biochanin A may be additive. The ATPase stimulatory effects indicate that biochanin A, as well as phloretin, probably do not bind to the ATP-binding site of P-gp, as has been reported for the flavonoids quercetin, apigenin, kaempferide, naringenin, genistein, and rutin (Shapiro and Ling, 1997a; Conseil et al., 1998).
To understand the interaction of flavonoids with P-gp and its substrate binding, we investigated the effects of flavonoids on [3H]azidopine photoaffinity labeling of P-gp. The inhibition of [3H]azidopine photoaffinity labeling of P-gp has been commonly used to indicate direct interaction with P-gp substrate binding (Castro and Altenberg, 1997). Both P-gp substrates and modulators such as vinblastine and verapamil have been shown to inhibit azidopine photoaffinity labeling of P-gp (Tamai and Safa, 1991; Christensen and LeBlanc, 1996). Flavonoids silymarin and morin were shown to significantly inhibit [3H]azidopine photoaffinity labeling of P-gp, but to different extents, suggesting that these flavonoids may interact with P-gp substrate binding directly either by competitive binding to the substrate-binding site or by binding to other drug-binding sites and changing protein conformation that results in decreased photoaffinity labeling. The inhibition by 100 μM biochanin A, morin, and phoretin is moderate (23.2 ± 6.3%, p > 0.05; 37.9 ± 28.4%, p < 0.05; and 30.2 ± 11.6%, p > 0.05, respectively) and might be comparable with that reported for genistein (42 ± 15% at 200 μM concentration) (Castro and Altenberg, 1997). Silymarin showed stronger inhibition (61.2 ± 20.7%, p < 0.001), which is comparable with that produced by 100 μM vinblastine.
In the preincubation studies, we have shown that preincubation with biochanin A, morin, or phloretin had no effect on [3H]DNM accumulation in MCF-7/ADR cells; consistent with this observation is our finding that the cellular P-gp level also showed no significant changes after preincubation with these flavonoids. Preincubation with silymarin was shown to increase [3H]DNM accumulation in MCF-7/ADR cells slightly but significantly (38 ± 5%, p < 0.001); however, Western blot analysis was not able to detect any significant changes in P-gp level after preincubation with silymarin.
In conclusion, the naturally occurring flavonoids biochanin A, morin, phloretin, and silymarin can inhibit P-gp-mediated drug efflux, and biochanin A and silymarin can potentiate doxorubicin cytotoxicity in P-gp positive cells. The underlying mechanism(s) may involve direct interaction with P-gp as evidenced by flavonoid modulation of P-gp ATPase activity, inhibition of [3H]azidopine photoaffinity labeling, and the observation that flavonoids do not change cellular P-gp level. In addition, the interactions of these flavonoids with P-gp may be different for specific flavonoids because these flavonoids demonstrated different effects on P-gp ATPase activity. Inferring the exact mechanism(s) by which flavonoids inhibit P-gp is difficult due to the complexity of the P-gp-substrate/modulator interactions. It is possible that some of these flavonoids (for example, biochanin A) might be used alone, or in combination with other P-gp inhibitors, to reverse MDR in the treatment of certain cancers. In addition, dietary supplements containing the flavonoids investigated in this study, such as red clover extracts (biochanin A as one of the main components) and milk thistle (silymarin), are widely used and it is possible that they may interact with coadministered drugs that are P-gp substrates, resulting in altered bioavailability and/or clearance.
Footnotes
-
This study was supported by grants from the New York State Health Research Science Board (EMPIRE grant), the Susan G. Komen Breast Cancer Foundation, and the U.S. Army Breast Cancer Program Contract DAMD17-00-1-0376.
-
DOI: 10.1124/jpet.102.044412
- Abbreviations:
- MDR
- multidrug resistance
- P-gp
- P-glycoprotein
- MRP1
- multidrug resistance-associated protein 1
- DNM
- daunomycin
- PBS
- phosphate-buffered saline
- DMSO
- dimethyl sulfoxide
- SRB
- sulforhodamine B
- Received September 16, 2002.
- Accepted November 22, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}