


Abstract
CYP2C9 was recently resequenced in 150 Asian subjects from Singapore. Several new coding variants were reported, and these variants are now named CYP2C9*14 (R125H), CYP2C9*15 (S162X), CYP2C9*16 (T299A), CYP2C9*17 (P382S), CYP2C9*18 (D397A), and CYP2C9*19 (Q454H). The CYP2C9*18 variant also contained an I359L change previously associated with the CYP2C9*3 allele. In this study, we assessed the functional consequences of the new coding changes. cDNAs containing each of the new coding changes were constructed by site-directed mutagenesis and expressed in a bacterial cDNA expression system, the allelic proteins were partially purified, and their ability to hydroxylate a prototype CYP2C9 substrate was assayed. Expression of cDNAs in Escherichia coli containing either the D397A change or the S162X (premature stop codon) could not be detected either spectrally or at the apoprotein level. CYP2C9.14 and CYP2C9.16 exhibited 80 to 90% lower catalytic activity toward tolbutamide at two substrate concentrations compared with wild-type CYP2C9.1. Kinetic analysis confirmed that CYP2C9.14 and CYP2C9.16 have a higher Km and a >90% lower intrinsic clearance of tolbutamide compared with wild-type CYP2C9.1. Both CYP2C9.17 and CYP2C9.19 proteins exhibited modest 30 to 40% decreases in catalytic activity toward tolbutamide. Thus, CYP2C9*15 and CYP2C9*18 may represent null alleles, whereas CYP2C9*14 and CYP2C9*16 allelic variants produce proteins that are clearly catalytically defective in vitro, indicating the existence of new defective putative alleles of CYP2C9 in Asians.
CYP2C9 is known to be one of the most important members of the drug-metabolizing cytochrome P450 (P450) isoforms in the human liver. Some of these drugs have narrow therapeutic indices, such as the anticoagulant warfarin and the anticonvulsant phenytoin (Goldstein et al., 1994; Lee et al., 2002). Other drugs metabolized principally by CYP2C9 include the antidiabetic drugs tolbutamide and glipizide, the diuretic torsemide (Miners and Birkett, 1998), the anti-hypertensive drug losartan, and numerous nonsteroidal anti-inflammatories, including flurbiprofen, ibuprofen, and diclofenac.
Many polymorphic alleles of CYP2C9 have been reported, with at least eight known or putative poor metabolizer (PM) alleles of CYP2C9 found in different racial groups. Two defective alleles, CYP2C9*2 (R144C) (Crespi and Miller, 1997) and CYP2C9*3 (I359L) (Sullivan-Klose et al., 1996), have been extensively studied both in vitro and in clinical studies (Lee et al., 2002). The CYP2C9*3 allele has lower affinity and markedly lower intrinsic clearance for numerous drugs both in vitro and in vivo. The CYP2C9*2 allele is more moderately defective. These two alleles have received the most study in white populations, where the frequencies of the defective CYP2C9*2 and CYP2C9*3 alleles are 11 and 8%, respectively. However, these alleles are found in much lower frequencies in African-American (3.0 and 0.5%, respectively) and most East Asian populations (0 and 1.6%, respectively) (Xie et al., 2001, 2002). CYP2C9 *4 (Ieiri et al., 2000) (I359T), which has been reported in Asians, and CYP2C9*5 (D360E) (Dickmann et al., 2001) (found in African-Americans) resemble CYP2C9*3 kinetically, exhibiting a high Km and low intrinsic clearance for a number of drugs. Both I359 and D360 occur in the substrate binding pocket. A null allele, CYP2C9*6, containing a base deletion and frame shift has also been identified in an individual with severe phenytoin toxicity (Kidd et al., 2001). Six new coding alleles, CYP2C9*7 to *12, were recently reported by our laboratory by resequencing genomic DNA from a racially diverse population (Blaisdell et al., 2004). Of these, CYP2C9*11 and perhaps CYP2C9*12 were predicted to be putative defective alleles based on in vitro metabolism of tolbutamide by the recombinant enzymes. Recently, there has been a report of an individual heterozygous for CYP2C9*3 and a novel CYP2C9 allele, CYP2C9*13 (L90P), in an Asian population. The latter was suggested to contribute to the PM phenotype (Si et al., 2004).
It is well known that there are racial and ethnic differences in both the frequency and types of polymorphisms of CYP2C9 (Xie et al., 2001, 2002). The identification of polymorphisms in different racial and ethnic populations is vital to understanding differences in clinical responses to drugs in these populations. An extensive review of allelic variants in the intron, exon, and 3′-untranslated regions of 11 candidate phase I drug-metabolizing genes, including CYP2C9, in white, African-American, and Asian populations has recently been published (Solus et al., 2004). This report catalogues the different polymorphisms reported in the various ethnic groups, some of which may contribute to ethnic differences in metabolism.
In our recent studies in a Southeast Asian population (59 Chinese, 37 Malay, and 26 Indians), CYP2C9 was sequenced for new mutations (Zhao et al., 2004) including the promoter, 300 base pairs downstream in the 3′-untranslated region, the splice-site junctions, and each of the nine CYP2C9 exons. The CYP2C9*2 and CYP2C9*3 alleles were found with varying frequencies in these groups (Indians > Chinese or Malay). In addition to the known coding variants, new alleles with coding changes were discovered. These included a new null variant, CYP2C9*15, containing a premature stop codon, S162X. Although warfarin dose requirements were measured in this population, the function of the new alleles could not be adequately assessed since these occurred in the heterozygous state.
In the present study, the function of the six new coding alleles found in Chinese and Indians was further assessed in a bacterial cDNA expression system. The newly discovered coding changes were introduced by site-directed mutagenesis into CYP2C9*1 and the recombinant proteins expressed in Escherichia coli. The recombinant CYP2C9 allelic proteins were solubilized, partially purified, and then assayed for tolbutamide hydroxylase activity. Two allelic variants, including one with a premature stop codon, S162X, and one with a D397A, could not be expressed as recombinant proteins in E. coli, suggesting that both may represent null alleles. Other recombinant allelic variants (T299A and R125H) exhibited markedly defective catalytic activity toward tolbutamide, suggesting they may also be poor metabolizer alleles. The linkages of the new coding changes to other single nucleotide polymorphisms (SNPs) are also reported herein.
Materials and Methods
[ring-U-14C]Tolbutamide was purchased from GE Healthcare (Little Chalfont, Buckinghamshire, UK). Human cytochrome b5 as well as NADPH-P450 oxidoreductase were purchased from Oxford Biomedical Research (Oxford, MI). All restriction enzymes used were purchased from New England Biolabs (Beverly, MA). All other chemicals used were purchased from Sigma-Aldrich (St. Louis, MO) or were the highest grade commercially available.
Site-Directed Mutagenesis of Expression Plasmids. Cloning and modification of the wild-type CYP2C9*1 cDNA for expression in pCW ori+ in a bacterial expression system was performed as previously described for other alleles (Ibeanu et al., 1996). The newly discovered coding SNPs were introduced into the wild-type construct using a Quick Change Site-Directed Mutagenesis kit (Stratagene, La Jolla, CA) with primers listed in Table 1. The SNP 1190A>C (D397A) was introduced alone without the I359L to determine the affect of this amino acid change on CYP2C9 expression or function. All mutants were confirmed by DNA sequencing.
Primers used for site-directed mutagenesis of CYP2C9*1
Expression and Partial Purification of CYP2C9 Allelic Variant Proteins. Each expression clone was grown in overnight cultures at 37°C from a single colony. The cultures contained Terrific Broth (ampicillin 100 μg/ml) and were then diluted 100-fold into 500 ml. Once the cultures achieved log phase (OD600 = 0.4–0.6), they were induced with isopropyl β-d-thio-galactoside (0.5 mM) and δ-aminolevulinic acid (0.5 mM) and left to shake (150 rpm) at 23 to 25°C for at least 72 h. Reduced CO spectra using a DW-2000 Spectrophotometer were used to measure cytochrome P450 content as described earlier (Omura and Sato, 1964). Once adequate levels of cytochrome P450 expression were detected (typically ∼72 h), the recombinant P450 was solubilized and partially purified by hydroxyapatite chromatography as previously described (Richardson et al., 1995; Ibeanu et al., 1996).
Tolbutamide Hydroxylation Assays. The tolbutamide assay has previously been described in detail (Blaisdell et al., 2004). Briefly, 20 pmol of partially purified recombinant P450 proteins was reconstituted with 0.3 μg/pmol P450 of 1,2-didodecanoyl-sn-glycero-3-phosphocholine, 4 pmol/pmol P450 of recombinant human NADPH-P450 oxidoreductase, and 2 pmol/pmol P450 of human cytochrome b5 at 37°C for 5 min, then placed on ice. Samples were then diluted and mixed into a final reaction volume of 250 μl containing 20 mM HEPES, pH 7.4, 0.1 mM ethylenediaminetetraacetic acid, 1.25 mM MgCl2, and [ring-U-14C]tolbutamide as substrate. Mixtures were preincubated at 37°C for 3 min, and then the reaction was initiated by the addition of 50 mM NADPH. For single-point assays, three to four separate purified preparations of each allele were reconstituted and then assayed with either 0.1 or 1 mM [ring-U-14C]tolbutamide; for kinetics, preparations from a single date were assayed with varying concentrations (0.075–3 mM) of substrate. All reactions were linear with time and protein under these conditions. The reactions were stopped after 45 min by the addition of methanol in an equal volume to reaction mixture. After centrifugation at 10,000g for 10 min, an aliquot of the supernatant was checked for total radioactivity by liquid scintillation (Beckman LS6500 Multi-Purpose Scintillation Counter; Beckman Coulter, Fullerton, CA). High-performance liquid chromatography was used to separate and quantitate metabolites as described previously (Blaisdell et al., 2004), with the mobile phase consisting of acetonitrile/0.1% trifluoroacetic acid (40:60). Retention times of the radiolabeled substrate and metabolite were compared with those of the unlabeled tolbutamide and hydroxytolbutamide standards (Sigma-Aldrich; BD Gentest, Woburn, MA).
Statistical Analysis. For each allelic protein, three separate rounds of expression and purification were performed concurrently with expression and purification of CYP2C9.1. The resulting three preparations of each mutant allele and four preparations of CYP2C9.1 were assayed for tolbutamide activity in triplicate. The data for each mutant were calculated as a percentage of the activity of the CYP2C9.1 wild type, which was copurified on the same date to minimize the effects of any day-to-day variability in the P450 purifications. Statistical significance of single point values was determined using SigmaStat 3.1 (Systat Software, Point Richmond, CA). Since there were four preparations of wild type and three of each mutant allele, analysis of variance was followed by Dunn's test, which allows for comparison of unequal groups of values. A single rectangular hyperbola nonlinear regression model (Michaelis-Menten) was used to analyze kinetic data using SigmaPlot version 9.0 (Systat Software Inc.).
Results
In the present study, we assess the functional consequences of six new alleles recently reported by one of our laboratories (Zhao et al., 2004). These new alleles are as follows: CYP2C9*14 (R125H), CYP2C9*15 (S162X), CYP2C9*16 (T299A), CYP2C9*17 (P382S), CYP2C9*18 (D397A/I359L), and CYP2C9*19 (Q454H). Their linkages to other SNPs have not been reported and are summarized in Table 2. Linkages are considered definitive for several alleles since the new SNPS occurred in individuals who were homozygous for the associated SNP. The allele CYP2C9*14 contains an SNP 374G>A coding for R125H but no other linkages. The allele CYP2C9*15 contains an SNP 485C>A coding for S162X that was found in a single individual heterozygous for a frequent promoter SNP–1188T>C. Therefore, additional studies with larger populations will be necessary to determine whether the two SNPs are on the same allele or separate alleles. The CYP2C9*16 allele, containing an SNP 895 A>G coding for T299A, was unequivocally linked to –1188T>C. The CYP2C9*17 allele contained a single SNP at 1144C>T (coding for P382S). The CYP2C9*18 allele contains three exonic SNPs: 1075A>C (I359L), 1190A>C (D397A), and 1425A>T (silent 475G) as well as five promoter SNPs (–1911T>C, – 1885C>G, – 1537G>A, – 1188T>C, and –982G>A). The individual was heterozygous for the new SNP 1190A>C but homozygous for the other SNPs (genotyped CYP2C9*18/CYP2C9*3). This allele is presumably derived from the CYP2C9*3 allele since all of the mutations except for 1190A>C (D397A) also occur on the CYP2C9*3 allele (Blaisdell et al., 2004). The CYP2C9*19 (1362G>C) (Q454H) allele contains a promoter SNP at –1188T>C. Three of the new variants (CYP2C9*14, CYP2C9*15, and CYP2C9*18) were reported in Indians, whereas CYP2C9*16, CYP2C19*17, and CYP2C9*19 were found in Chinese.
Structure of newly discovered CYP2C9 alleles in Southeast Asians
P450 Expression. In the present study, each of the new coding changes was introduced individually by site-directed mutagenesis, the cDNAs were expressed in E. coli, and the recombinant proteins were solubilized after ∼72 h and partially purified. Of the six mutant alleles, only four could be expressed in bacteria. Despite three separate attempts to express and purify any solubilized P450 proteins, expression of CYP2C9.15 could not be detected spectrally in bacteria through 72 h, and Western blot analysis of the partially purified bacterial extracts with a polyclonal antibody to CYP2C9 showed no evidence of CYP2C9 apoprotein (data not shown). This is not surprising since the CYP2C9*15 allele has a premature stop codon. However, it was somewhat surprising that the mutant allele containing D397A (found in the CYP2C*18 allele) could also not be expressed in bacteria at the spectral or apoprotein level (Western blotting of the solubilized bacterial extracts showed no apoprotein in contrast to that seen with wild-type CYP2C9.1 in three separate experiments). We made no subsequent attempts to construct the D397A amino acid change on the defective CYP2C9*3 background since the single mutation D397A appeared to prevent P450 expression in bacteria.
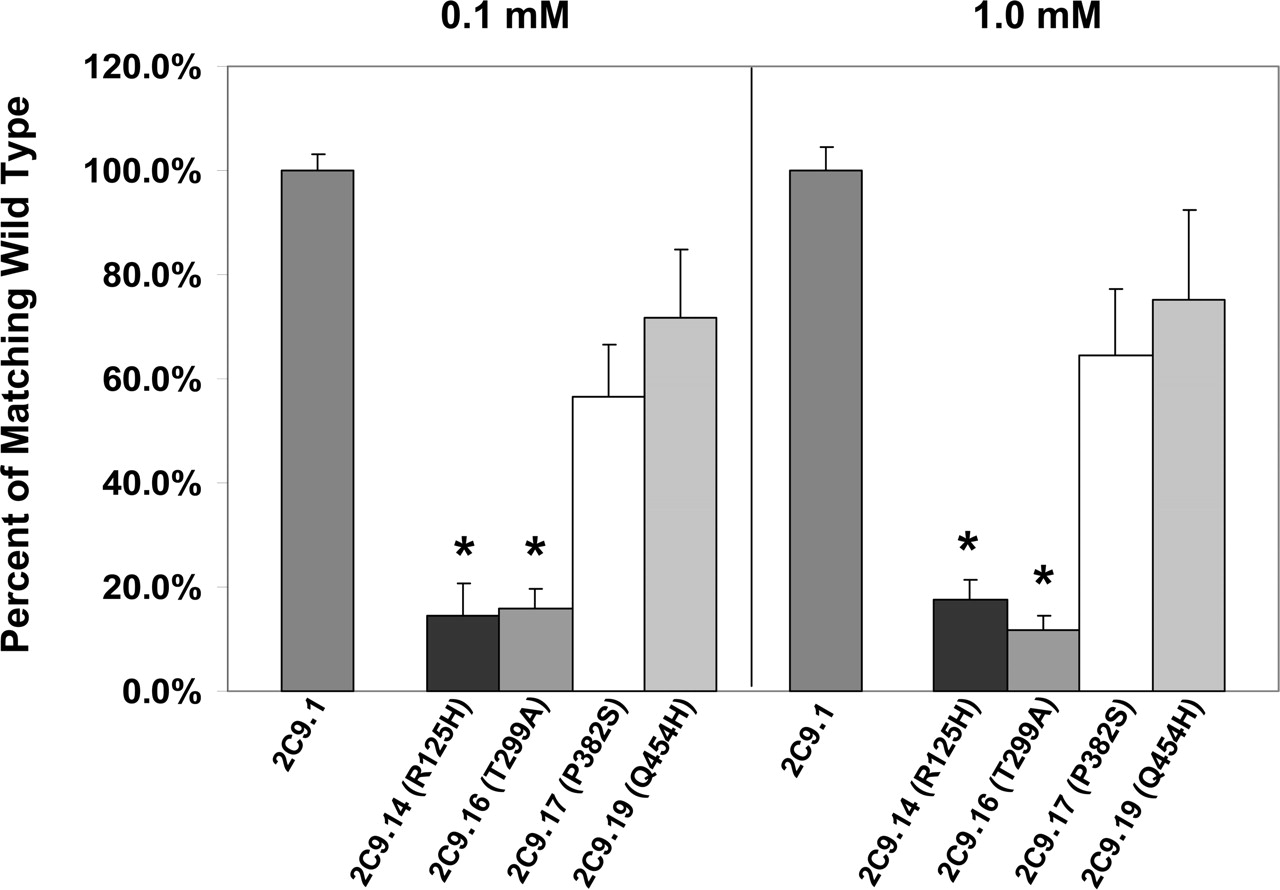
Tolbutamide Activity and Kinetics Data. The remaining four recombinant CYP2C9 allelic proteins were expressed in bacteria at levels comparable with those seen with wild-type CYP2C9.1 and were then partially purified (three separate rounds of expression and purification for each mutant allele and four concurrent preparations of the wild-type allele). All 16 partially purified preparations were assayed for tolbutamide hydroxylase activity. Results are shown in Fig. 1. Recombinant CYP2C9.14 and CYP2C9.16 were markedly defective in the metabolism of tolbutamide, exhibiting a >80% decrease in catalytic activity at low and high concentrations of substrate (0.1 and 1.0 mM). These results were significantly different from wild type (P < 0.05). Moreover, these data represent three separate preparations of each enzyme, ruling out variation in purification. Detailed kinetic analysis of the CYP2C9.14 allelic protein versus wild-type demonstrated a 5-fold increase in the Km, a 65% decrease in the Vmax, and a >93% decrease in the intrinsic clearance compared with wild-type CYP2C9.1 (Table 3 and Fig. 2). Examination of the crystal structure of CYP2C9 indicates R125 is on the surface of the protein near the heme. This amino acid is equivalent to R126 in CYP2B6, which is one of several amino acids that appears to be part of binding sites for cytochrome b5 and cytochrome P450 reductase (Bridges et al., 1998). Recombinant CYP2C9.16 had a 3.5-fold increase in the Km, a 69% decrease in the Vmax, and a >92% decrease in the intrinsic clearance of tolbutamide compared with CYP2C9.1. The T299A substitution found in CYP2C9.16 is on the back side of the I helix, close to the predicted oxygen binding site. Threonine 299 is highly conserved in family 2. CYP2C9.17 (P382S) and CYP2C9.19 (Q454H) exhibited somewhat lower tolbutamide hydroxylase activity than wild-type CYP2C9 (40 and 20% decreases, respectively), but this was not statistically significant with the number of samples analyzed. Moreover, more complete kinetic analysis indicated that the Km, Vmax, and intrinsic clearance of CYP2C9.17 and CYP2C9.19 were similar to CYP2C9.1.
Comparison of kinetic values for different 2C9 allelic proteins
Data are values ± S.E. Triplicate analyses were performed at each substrate concentration. Alleles were assayed on the same day. Kinetic values were determined using SigmaPlot 9.0 (Systat Software) as described under Materials and Methods. Values are representative of two separate experiments using two different purified preparations of each allelic protein.

Tolbutamide hydroxylase activity of recombinant mutant CYP2C9 allelic proteins compared with that of wild-type CYP2C9.1 protein. The new mutant CYP2C9 cDNAs were constructed by site-directed mutagenesis from the wild-type CYP2C9*1 cDNA, expressed in E. coli and partially purified as described under Materials and Methods. Separate rounds of expression and purification resulted in three purified preparations of each variant allele, which were expressed and purified simultaneously with wild-type CYP2C9.1. All 16 partially purified preparations (three of each mutant allele and four of wild-type CYP2C9.1) were assayed the same day in triplicate at high (1 mM) and low (0.1 mM) substrate concentrations. The values of each preparation of the mutant alleles were calculated as a percentage of enzymatic activity of the wild-type allele that had been purified on the same day. *, significantly different from wild-type CYP2C9, P < 0.05.
Discussion
Polymorphisms of CYP2C9 have been studied extensively in whites. CYP2C9*2 and CYP2C9*3 are the most common defective alleles in the white populations (11 and 8.4%, respectively) but have significantly lower frequencies in African populations (2.9 and 1.3%, respectively) and Eastern Asian populations such as Chinese (0 and 3.3%), Japanese (0 and 2.2%), and Koreans (0 and 1.1%, respectively) (Xie et al., 2001, 2002). However, higher frequencies of these two alleles have been reported in Indian populations compared with Eastern Asians (4–6% and 8–13.8%) (Zainuddin et al., 2003; Jose et al., 2005). Similarly, recent studies from one of our laboratories (Zhao et al., 2004) reported frequencies of 4 and 11.5%, respectively, for the CYP2C9*2 and CYP2C9*3 alleles in Indians, 0 and 3.3% in Chinese, and 0 and 6.3% in Malays from Singapore who were receiving maintenance warfarin therapy. Importantly, several new alleles with coding changes in CYP2C9 were discovered in Asians. Three of the new coding mutations were detected in Indians and three in Chinese. Since these new mutations were found in single individuals in the heterozygous state, it was difficult to draw meaningful conclusions concerning the functional consequences of these new variant alleles based on warfarin dosage requirements. Therefore, in the present study, we assessed the function of these new alleles using a cDNA expression system and tolbutamide as the probe drug to study metabolism of the recombinant proteins in vitro.
One of the new alleles in this study, CYP2C9*15, contained a nonsense mutation, Ser162X, producing an early termination codon (Zhao et al., 2004). The present cDNA expression studies confirm that this is a null allele. Neither CYP2C9 apoprotein nor holoprotein was expressed in E. coli. This is the second report of a null allele of CYP2C9. The first reported null allele, CYP2C9*6, was discovered in an African-American woman with severe phenytoin toxicity. This individual was found to be homozygous for a deletion of an adenine at base pair 818 of the cDNA (Kidd et al., 2001).
The CYP2C9*18 allele, occurring in a single Indian (1 of 52 alleles), contained two coding mutations, D397A and I359L (also found on the defective CYP2C9*3 allele). Since the CYP2C9*3 allele itself is markedly defective, we first tested the effect of the single mutation D397A. Similar to the null variant CYP2C9*15, the cDNA carrying the D397A mutation could not be expressed in E. coli despite three repeated rounds of expression, followed by solubilization and purification of the extract. There was no spectral evidence for CYP2C9 holoprotein at any time during the 72-h expression period or after hydroxyapatite purification of the extract. Western blotting with a polyclonal antibody to CYP2C9 also showed no evidence of the apoprotein in partially purified bacterial extracts. These data suggest this amino acid change may prevent expression in E. coli. In the crystal structure of CYP2C9 (Williams et al., 2003), D397 is on the surface of the protein; however, it is highly conserved in all the structures that have been determined (CYP2A6, CYP2B4, CYP2C5, CYP2C8, CYP2C9, and CYP3A4) (Johnson et al., 2002; Mestres, 2004; Scott and Halpert, 2005) as well as most members of cytochrome P450 families 1 to 3. It is uncertain whether this D397A change would also prevent expression in humans; however, this change may represent a null allele. This hypothesis is based on a single cDNA expression system. Additional experimentation with other cDNA expression systems and additional clinical data will be helpful to clarify this possibility.

Kinetics of tolbutamide hydroxylation by CYP2C9.1, CYP2C9.14, CYP2C9.16, CYP2C9.17, and CYP2C9.19 allelic proteins. All reactions include 20 pmol CYP2C9, 80 pmol human NADPH-cytochrome P450 oxidoreductase, 40 pmol P450 of human cytochrome b5, and tolbutamide concentrations between 0.5 and 3 mM [ring-U-14C] in a reaction volume of 250 μl for 45 min.
The CYP2C9*14 (R125H) allele was found in an Indian individual (1 of 52 alleles) (Zhao et al., 2004). The present in vitro results indicate that recombinant CYP2C9.14 is highly defective, with a >90% reduction in tolbutamide clearance. Examination of the crystal structure of CYP2C9 indicates R125 is on the surface of the protein near the heme ligand. R125 is highly conserved in family 2. This amino acid is equivalent to R126 in CYP2B6. R126 is one of several amino acids that appear to be part of binding sites for cytochrome b5 and cytochrome P450 reductase (Bridges et al., 1998). These basic amino acids were also proposed to help neutralize the charge of buried heme propionic acid residues, one of which forms hydrogen bonds with the adjacent R124 of CYP2C9.
The CYP2C9*16 (T299A) allele was found in one Chinese individual (1 of 118 alleles). The in vitro data indicated that the recombinant allelic protein had a markedly decreased affinity, maximum velocity, and 90% decrease in the intrinsic clearance for tolbutamide. Thr299 occurs on the back side of helix I, close to the predicted oxygen binding site. It is highly conserved in family 2. Our data indicate CYP2C9*16 is a new putative PM allele, which could possibly have an impact on the metabolism of CYP2C9 substrates in Asians. The clinical effects of this allele should be examined in a larger population.
Two coding changes resulted in only modest effects on catalytic activity. The results with recombinant CYP2C9.17 protein indicated a possible moderate reduction in tolbutamide hydroxylase activity, but this difference was not statistically significant. Moreover, kinetic parameters of this enzyme were similar to those of CYP2C9.1. The P382 is located on the surface of the enzyme in a turn following β-sheet 2, and it has been found to be highly conserved in all structurally characterized microsomal P450s. The CYP2C9*19 allele was discovered in one individual of Chinese origin (1 of 118 alleles). The tolbutamide hydroxylase activity of the recombinant CYP2C9.19 was also 20 to 30% lower than wild type, but again not statistically significant, nor did kinetic parameters appear to differ from wild-type CYP2C9.1. Q454 is also on the surface at the end of a helix. These two latter alleles will require further clinical study to determine whether they have modest effects on CYP2C9 activity in vivo.
Warfarin maintenance doses for Asians are known to be lower than those of whites (Loebstein et al., 2001; Zhao et al., 2004). However, the frequency of PM alleles is lower in most Asian groups than in whites. This discrepancy could be explained by the possible existence of new undiscovered alleles in Asians, including SNPs in the introns or upstream regions. However, warfarin dosage is also known to be affected by dietary changes and drug interactions, including induction of CYP2C9 by prior exposure to drugs and herbal remedies (Loebstein et al., 2001; Chen et al., 2004). Various upstream SNPs are more frequent in Asians, namely an SNP at –1188 (GenBank National Center for Biotechnology Information no. AL359672) (Shintani et al., 2001; King et al., 2004). This SNP is identical to the –1189 denoted in our earlier study (Zhao et al., 2004) due to the use of an alternate older reference sequence for numbering in the Asian literature (GenBank National Center for Biotechnology Information no. L16877). In Asians, the most frequent allele is –1188C, whereas in whites, the more frequent allele is –1188T. Several studies have addressed the possible effects of upstream polymorphisms on warfarin dosage (Shintani et al., 2001; King et al., 2004; Takahashi et al., 2004). To date, there is no clear evidence that any of these SNPs have a role in promoter activity. Recently, new studies have shown that polymorphisms in the vitamin K epoxide reductase complex 1 (the molecular target of warfarin) and other vitamin K-dependent proteins (factor II and factor VII genes) also modulate the anticoagulant effects of warfarin, thereby altering dose requirements (D'Ambrosio et al., 2004; Rost et al., 2004; D'Andrea et al., 2005; Yuan et al., 2005). Yuan et al. (2005) suggested the possibility that ethnic differences in warfarin dose requirements might be attributed to ethnic differences in the frequency of a mutation found in the promoter region of the VKORC1 gene in Chinese versus whites. Therefore, multiple genetic factors (including polymorphisms in CYP2C9 as well as vitamin K-dependent genes) modulate warfarin dosage.
In summary, the present study functionally assessed novel coding SNPs discovered in Chinese and Indians in a recombinant cDNA expression system. We confirm the existence of at least one new null allele (CYP2C9*15) and suggest that CYP2C9*18 may also potentially represent a null allele based on lack of expression in a cDNA expression system. In addition, CYP2C9*14 and CYP2C9*16 represent new putative defective alleles in Asian populations based on markedly decreased intrinsic clearance of tolbutamide by both recombinant allelic proteins. This study highlights the existence of ethnic variability in the pharmacokinetics and pharmacodynamics of common drugs metabolized by CYP2C9. Clinical studies of these variants in larger Asian populations are needed to assess the role of these mutations in man. Our results show that defective alleles of CYP2C9 are more common than previously realized in various Asian populations, and additional genetic testing for the new defective alleles may be helpful in these populations.
Acknowledgments
We thank Eric Johnson (Scripps Research Institute, La Jolla, CA) for expertise concerning the amino acid positions of the variant alleles based on the crystal structure of CYP2C9. We also thank Stephen Ferguson (Laboratory of Pharmacology and Chemistry, National Institute of Environmental Health Sciences) for useful discussions and insightful comments on this topic.
Footnotes
-
doi:10.1124/jpet.105.091181.
-
ABBREVIATIONS: P450, cytochrome P450; PM, poor metabolizer; SNP, single nucleotide polymorphism.
- Received June 17, 2005.
- Accepted August 8, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}