Article Text

Abstract
Background/aims: The apical sodium dependent bile acid transporter ASBT (SLC10A2) contributes substantially to the enterohepatic circulation of bile acids by their reabsorption from the intestine. In the rat, its adaptive regulation was observed in the kidneys, cholangiocytes, and terminal ileum after bile duct ligation. Whether adaptive regulation of the human intestinal ASBT exists during obstructive cholestasis is not known.
Methods: Human ASBT mRNA expression along the intestinal tract was analysed by real time polymerase chain reaction in biopsies of 14 control subjects undergoing both gastroscopy and colonoscopy. Their duodenal ASBT mRNA expression was compared with 20 patients with obstructive cholestasis. Additionally, in four patients with obstructive cholestasis, duodenal ASBT mRNA expression was measured after reconstitution of bile flow.
Results: Normalised ASBT expression in control subjects was highest (mean arbitrary units (SEM)) in the terminal ileum (1010 (330)). Low ASBT expression was found in colonic segments (8.3 (5), 4.9 (0.9), 4.8 (1.7), and 1.1 (0.2) in the ascending, transverse, descending, and sigmoid colon, respectively). Duodenal ASBT expression in control subjects (171.8 (20.3)) was found to be approximately fourfold higher compared with patients with obstructive cholestasis (37.9 (6.5); p<0.0001). Individual ASBT mRNA expression was inversely correlated with bile acid and bilirubin plasma concentrations. In four cholestatic patients, average ASBT mRNA increased from 76 (18) before to 113 (18) after relief of cholestasis (NS). Immunohistochemical assessment indicated that ASBT protein was expressed on the apical surface of duodenal epithelial cells.
Conclusion: Obstructive cholestasis in humans leads to downregulation of ASBT mRNA expression in the distal part of the human duodenum.
- ASBT, apical sodium dependent bile acid transporter
- CBDL, common bile duct ligation
- UDCA, ursodeoxycholic acid
- 75SeHCAT, 75Se-homocholic acid taurine
- MRP, multidrug resistance associated protein
- ERCP, endoscopic retrograde cholangiopancreatography
- PPARα, peroxisome proliferator activated receptor α
- cholestasis
- ASBT
- transporter
- bile acids
Statistics from Altmetric.com
- ASBT, apical sodium dependent bile acid transporter
- CBDL, common bile duct ligation
- UDCA, ursodeoxycholic acid
- 75SeHCAT, 75Se-homocholic acid taurine
- MRP, multidrug resistance associated protein
- ERCP, endoscopic retrograde cholangiopancreatography
- PPARα, peroxisome proliferator activated receptor α
Bile acids are amphipathic steroidal compounds derived from the enzymatic catabolism of cholesterol in the liver by cytochrome P450 isoform 7A1 (CYP7A1). In the small intestine, bile acids emulsify dietary fats and lipid soluble vitamins. They participate in the regulation of pancreatic secretion and release of gastrointestinal peptides.1 Through the coordinated action of several transport proteins expressed in hepatocytes, cholangiocytes, as well as in enterocytes and in proximal tubular cells of the kidney, an efficient enterohepatic circulation of bile acids is maintained.2 One of these transport proteins, the apical sodium dependent bile acid transporter ASBT (SLC10A2) has been detected in the ileum, caecum, and kidney3 and mediates the uptake of bile acids from the lumen of the intestine, from renal tubules, and from cholangiocytes. ASBT is a 348 amino acid protein that transports conjugated and unconjugated bile acids with a high efficiency.3 Subjects with mutations in the ASBT gene suffer from congenital diarrhoea and steatorrhoea due to bile acid malabsorption.4 Pharmacological inhibition of ASBT leads to interruption of enterohepatic circulation of bile acids with changes in cholesterol and bile acid homeostasis.5–9 Adaptive induction of asbt mRNA expression, transporter protein, and transport activity was observed in cholic acid fed rats10 whereas decreased asbt expression was found in the ileum of bile duct ligated rats.11 In addition, in Wistar rats, a marked reduction in asbt protein expression was observed in microsomal membrane fractions from whole kidney after ligation of the common bile duct (CBDL) with consequent increased urinary bile acid excretion. These results indicate that asbt is adaptively regulated in different tissues during obstructive cholestasis in the rat.
Lanzini and colleagues12 studied the effects of cholestasis on intestinal bile acid transport in 14 subjects with chronic cholestasis due to primary biliary cirrhosis before and during ursodeoxycholic acid (UDCA) administration. Prolonged retention of the bile acid analogue 75Se-homocholic acid taurine (75SeHCAT) in patients with primary biliary cirrhosis was observed compared with healthy controls and patients with Crohn’s disease. Retention of 75SeHCAT decreased with UDCA treatment, inferring that luminal bile acid levels may be involved in the regulation of ASBT gene regulation.
Virtually all bile salt transporter systems are subject to extensive regulation, mainly at the level of gene transcription. These regulatory mechanisms represent adaptive responses to intracellular accumulation of bile salts and other amphipathic molecules. However, little is known about adaptive regulation of ASBT expression in humans. The aim of this study was therefore to investigate expression of ASBT in the duodenum of healthy subjects and compare the results with patients with obstructive cholestasis. Because direct analysis of ASBT gene expression in samples from the terminal ileum cannot be performed in humans with obstructive cholestasis for obvious ethical reasons, in the first part of this study we analysed ASBT mRNA expression in healthy subjects in different segments of the human intestine. We found that ASBT mRNA was also expressed in the duodenum but to a lesser extent than in the terminal ileum.
Additionally, we analysed gene expression of multidrug resistance associated protein 3 (MRP3), which is expressed on the basolateral membrane of the enterocyte13 and which is capable of transporting bile salts,14 as well as expression of multidrug resistance associated protein 2 (MRP2). Human duodenal MRP2 protein expression was downregulated during cholestasis.15 Biopsies of the duodenum of patients with obstructive cholestasis were obtained during therapeutic endoscopic retrograde cholangiopancreatography (ERCP).
PATIENTS AND METHODS
Patients
Fourteen healthy subjects (seven males, seven females) were enrolled into the first part of the study after giving informed consent. The indication for combined upper and lower gastrointestinal tract endoscopy was a cancer screening programme. Biopsies were obtained from the duodenum, terminal ileum, and from different defined regions of the colon. In the second part of the study, 20 cholestatic patients were enrolled after giving written informed consent. Biopsies were obtained from the duodenum in these patients. Of 10 patients with obstructive tumours, four had carcinomas of the pancreatic head, four had cholangiocarcinomas (Klatskin tumour), and two had metastatic diseases. Ten patients had benign diseases (eight patients with choledocholithiasis and two patients with a benign stenosis of the common bile duct). Obstructive jaundice was defined: (1) on the basis of biochemical parameters (bilirubin, γ-glutamyltransferase, and alkaline phosphatase) and (2) on imaging procedures (ultrasound and ERCP), demonstrating a dilated bile duct system. All control patients had normal values of the above parameters. Patients were only included if they were not taking any medication known to affect ASBT, MRP2, or MRP3 expression. Demographic details are given in table 1.
Patient characteristics of the two study groups
Materials
All chemicals were of the highest quality available and were obtained from commercial sources.
Real time PCR analysis of human ASBT, MRP2, MRP3, PPAR-α, and villin (VIL1) mRNA (TaqMan assay)
Using standard pinch forceps, four intestinal biopsies were taken from each anatomical site (the distal part of the duodenum, terminal ileum, and different segments of the colon) during an interventional ERCP procedure (patients with obstructive cholestasis) or during gastroscopy and colonoscopy (control subjects). Biopsies were immediately submerged in a tube with RNAlater (Ambion) and stored at −80°C until further processing. For RNA isolation, two biopsies from each donor were homogenised for 30 seconds (Polytron PT 2100; Kinematika AG, Switzerland) and RNA was extracted using the RNeasy Mini Kit (Qiagen GmbH, Hilden, Germany) following the manufacturer’s instructions. RNA was quantified with a GeneQuant photometer (Pharmacia, Uppsala, Sweden). After DNase I digestion (Gibco Life Technologies, Basel, Switzerland) 1.5 μg of total RNA were reverse transcribed by Superscript (Gibco Life Technologies) according to the manufacturer’s protocol, using random hexamers as primers.
TaqMan analysis was carried out on a 7900HT sequence detection system (Applied Biosystems, Rotkreuz, Switzerland). Polymerase chain reaction (PCR) cycling conditions were 10 minutes at 95°C followed by 40 cycles of 15 seconds at 95°C and one minute at 60°C. Each TaqMan reaction contained 10 ng of cDNA in a total volume of 10 μl. TaqMan Universal PCR Mastermix from Applied Biosystems was used. Concentrations of primers and probes were 900 nM and 225 nM, respectively. They were designed according to the guidelines of Applied Biosystems with help of the Primer Express 2.0 software (corresponding sequences are listed in table 2). Primers were synthesised by Invitrogen (Basel, Switzerland) and probes by Eurogentec (Seraing, Belgium). All samples were run in triplicate. TaqMan PCR products were confirmed by sequencing (Microsynth GmbH, Balgach, Switzerland). Non-reverse transcribed RNA served as a negative control. No significant amplification was observed in these samples.
Primers and probes for TaqMan analysis
For each sample, the number of gene transcripts (ASBT, MRP2, MRP3, peroxisome proliferator activated receptor α (PPARα)) and the number of villin transcripts were determined. By calculating the ratio of gene/villin mRNA, gene expression was normalised to enterocyte content. Determination of villin, an enterocyte specific, constitutively expressed protein, can be used to control for the variation in enterocyte content in biopsies.16,17 For absolute quantification we used external standard curves. Standards were gene specific cDNA fragments that cover the Taqman primer/probe area and were obtained by PCR amplification. For ASBT, MRP2, and MRP3, we used cDNA from Caco-2 cells as a template whereas for PPARα, cDNA from duodenal tissue was used. Gene specific PCRs were performed with components from Applied Biosystems (AmpliTaq Gold; 10× PCR buffer, dNTPs, MgCl2). Each reaction contained 25 ng of cDNA and 300 nM of each primer (corresponding sequences are listed in table 3) in a total volume of 25 µl. Thermal cycling was conducted using a Mastercycler personal from Eppendorf (Hamburg, Germany). All PCR products were purified by running a 1.5% agarose gel and a subsequent gel extraction (gel extraction kit; Qiagen). Standards were quantified using the PicoGreen reagent (Molecular Probes, Eugene, Oregon, USA) and were checked by sequencing (Microsynth GmbH, Balgach, Switzerland).
Primers for cDNA standards
Standard curves were generated by plotting the obtained Ct values (cycle number when a sample reaches the threshold) as a function of the log(10) of the corresponding dilution factor and using linear regression. Curves were linear with correlation coefficients of 0.996, 0.998, 0.999, 0.997, and 0.998 for ASBT, villin, MRP2, MRP3, and PPARα, respectively. The corresponding PCR efficiencies (calculated as 10(1/−slope)−1) were 93%, 95.5%, 89.1%, 95%, and 97.1%, respectively. Each standard curve also included no template controls.
The sensitivity of all TaqMan assays was found to be adequate for quantifying all transcripts in the analysed intestinal tissues. Only in samples from the sigmoid colon did expression of ASBT mRNA appear to be so low that accurate quantification of transcript numbers was not possible for all biopsies. The precision of the ASBT determination was assessed by measuring 10 aliquots of duodenal cDNA (each in triplicates). The intra-assay coefficient of variation (CV) was 4.69%. Interassay CV was not determined as for every gene all samples could be placed onto one 384 well plate. Accuracy of the method was determined by measuring a standard with a known amount of template (n = 10 in triplicates). Deviation of the mean value from the true value was 14.85%.
Immunohistochemical assessment
For immunohistochemical assessment of ASBT expression in human intestinal tissue, a polyclonal rabbit antihuman ASBT (generous gift from Dr PA Dawson, Wake Forest University Baptist Medical Center, Winston-Salem, North Carolina, USA) was used. This antibody was raised against the carboxyl terminal 39 amino acids of human ASBT that was expressed as a glutathione S-transferase-ASBT fusion protein. The human ASBT antibody has been previously used to measure ASBT protein expression in human ileal biopsies.18
Human intestinal tissue was mounted in OCT compound (Sakura Finetek, Zooterwoude, the Netherlands), frozen in liquid nitrogen, and stored at −70°C. Sections (5 μm) of human intestine were air dried overnight and a periodate-lysine-paraformaldehyde solution (3%) was used for post fixation. Then sections were washed with washing solution (TBS/NaCl, Tween 0.05%) and incubated with goat serum (Vectastain, Elite kit; Vector Laboratories, Burlingame, California, USA) for 30 minutes at room temperature as blocking solution. Tissue sections were incubated with a 1:400 dilution of the polyclonal rabbit antihuman ASBT overnight at 4°C. Samples were washed three times with washing solution and incubated with the goat antirabbit IgG secondary antibody for 30 minutes at room temperature. After three washes with the washing solution, a perhydral solution (H2O2 (0.3%), sodium azide (0.1%) in phosphate buffered saline) was used to destroy endogenous peroxidase activity. Staining was performed with the avidin/biotinylated enzyme complex (ABC method) according to the manufacturer’s instructions (Vectastain, Elite kit; Vector Laboratories). For detection, 3-amino-9-ethylcarbazole, which forms a red end product, was used (Biogenex, San Ramon, California, USA). Sections, which served as negative controls, were incubated only with the goat antirabbit IgG secondary antibody. Biopsies from normal terminal ileum were used as a positive control.
Bilirubin and bile acid plasma concentrations
Blood samples from subjects were obtained shortly before the endoscopic procedure. Bilirubin plasma concentrations were measured by a modified Malloy-Evelyn method (BIL-T; Roche Diagnostics, Mannheim, Germany). Fasting plasma levels of bile acids were determined as described previously.19 In brief, bile acids were extracted with Bond-Elut C18 cartridges (Analytichem International, San Diego, California, USA), solvolysis was performed to cleave sulphate groups, and enzymatic hydrolysis was performed to deconjugate bile acid amidates. Deconjugated bile acids were isolated by extraction on Lipidex 1000 (Packard Instruments, Groningen, the Netherlands) and were then methylated and trimethylsilylated for gas chromatography. Capillary gas chromatography was performed using a Carlo Erba Fractovap 4160 gas chromatograph (Carlo Erba Instruments, Hofheim, Germany). Bile acid derivatives were separated on a fused silica capillary CP Sil 19 CB column coated with chemically bonded OV-1701 (25 m×0.33 mm; Chrompack, Middelburg, the Netherlands). Hydrogen was the carrier gas (P = 0.6 kg/cm2). A temperature programme from 140°C to 270°C with 8°/min was started after on-column injection. Eluted bile acid derivates were detected by a flame ionisation detector. Fasting plasma samples were stored at −20°C until analysed.
Statistics
All values are expressed as means (SEM). The impact of different parameters (bilirubin and bile acid concentration, body mass index, age, and sex) on the variability of ASBT expression was investigated by multilinear regression analysis. ASBT expression in icteric patients was compared with that of healthy controls by analysis of variance (ANOVA). Regional ASBT mRNA expression was compared with expression in the duodenum by repeated measurement ANOVA with linear contrasts. Correlation of serum bilirubin and bile acid concentrations was performed using Spearman’s rank correlation coefficient (rho). Differences in demographic characteristics between icteric patients and controls were analysed by ANOVA of the χ2 test, as appropriate. All comparisons were performed as two sided comparisons using the SPSS for Windows software (version 12.0). Level of significance was p<0.05
RESULTS
Expression pattern of ASBT mRNA in the human intestine
Human ASBT mRNA expression was studied in 14 control subjects (seven women and seven men) who were undergoing a combined gastroscopy and colonoscopy. Results were normalised by calculation of the ASBT/villin ratio. Normalised ASBT expression (mean (SEM) arbitrary units) was 171.8 (20.3) in the duodenum, 1010 (330) in the terminal ileum, 8.3 (5) in the ascending colon, 4.9 (0.9) in the transverse colon, 4.8 (1.7) in the descending colon, and 1.1 (0.2) in the sigmoid colon, respectively (fig 1).

mRNA expression of apical sodium dependent bile acid transporter (ASBT) in different gut segments. Data represent the means (SEM) of biopsies from 14 healthy subjects. AU, arbitrary units.
Duodenal expression of ASBT mRNA and PPARα mRNA
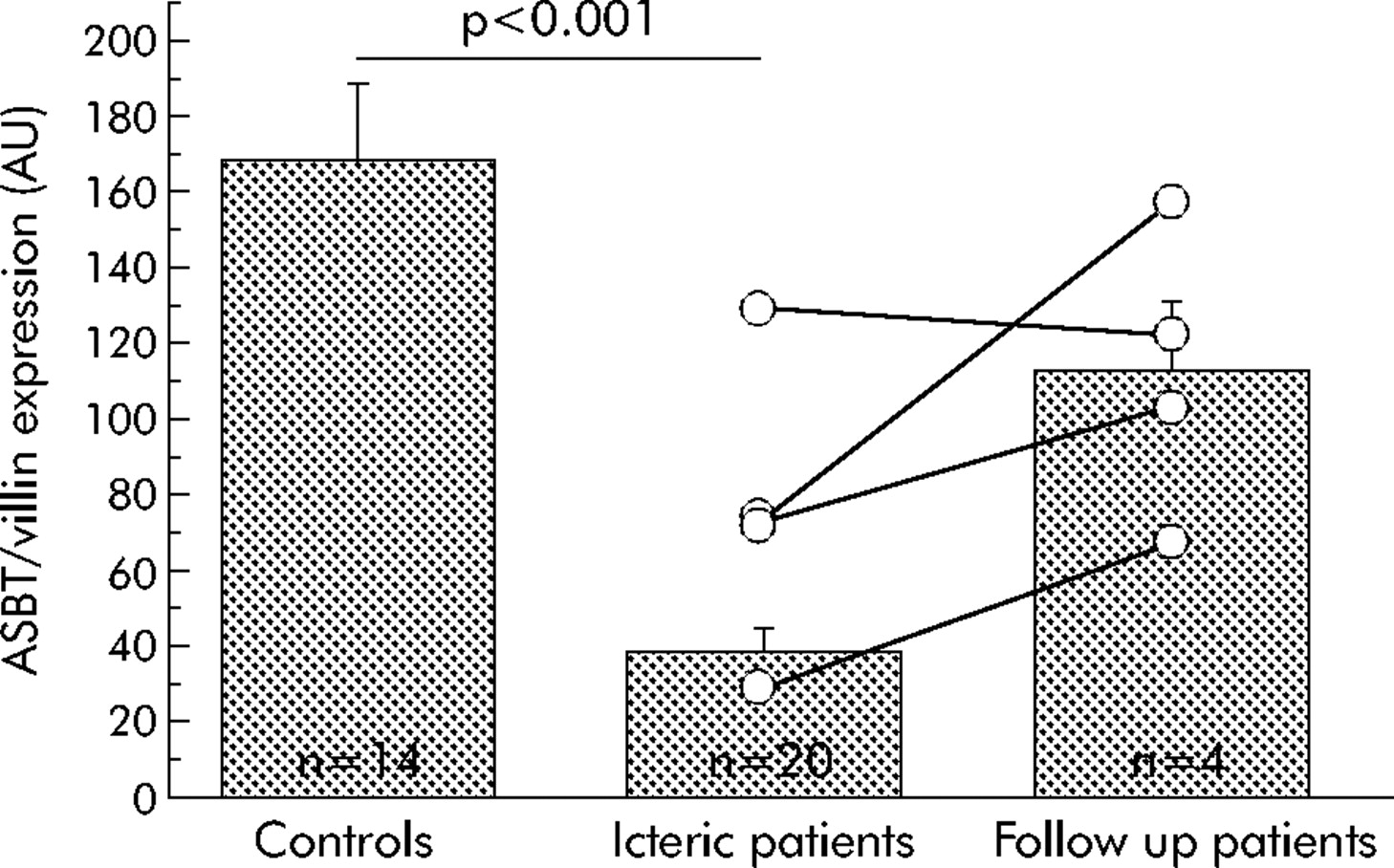
Adaptive regulation of ASBT expression in obstructive cholestasis was investigated by quantification of ASBT mRNA levels in duodenal biopsies of 20 patients with obstructive cholestasis and compared with levels obtained in 14 control subjects. As shown in fig 2, ASBT mRNA expression (ASBT/villin ratio (mean (SEM)) was about fourfold lower in patients with obstructive cholestasis (37.9 (6.5)) compared with control subjects (171.8 (20.3); p<0.001). Patients with obstructive cholestasis due to a tumour showed a trend for lower ASBT mRNA expression compared with patients with a benign aetiology of obstructive cholestasis (31.2 (7.3) for tumour induced and 44.6 (10.8) for benign obstruction); however, the difference was not statistically significant. Plasma bilirubin levels were 301.6 (64.9) and 112.6 (20.0) mmol/l (p = 0.02) and bile acid levels were 168.4 (45.3) and 76.8 (22.8) μmol/l (NS) for patients with and without tumours, respectively. PPARα mRNA expression was not significantly different between cholestatic patients and controls.

Comparison of apical sodium dependent bile acid transporter (ASBT) mRNA expression in patients with obstructive cholestasis (n = 20) compared with control subjects (n = 14) and follow up patients (n = 4). Data are means (SEM). In the scatterplot, lines connect individual ASBT mRNA expression pre- and post reconstitution of bile flow in four follow up patients. AU, arbitrary units.
Correlation of duodenal ASBT mRNA expression with bilirubin and bile acid plasma concentration
Bilirubin and bile acid plasma concentrations were inversely correlated with ASBT mRNA expression (rho = −0.863, p<0.001 and rho = −0.722, p<0.001, respectively). The correlation with ASBT mRNA expression was similar for bilirubin and bile acid concentrations. Using ASBT mRNA expression, an almost perfect separation was obtained between icteric patients and healthy controls (fig 3A, B).

Individual correlation of bilirubin (A) and bile acid (B) plasma concentrations with apical sodium dependent bile acid transporter (ASBT) mRNA expression in patients with obstructive cholestasis and control subjects. Data are means (SEM). Arrows connect individual values pre- and post reconstitution of bile flow in four follow up patients. AU, arbitrary units.
Effect of reconstitution of bile flow on ASBT mRNA expression
In five of the 20 icteric patients, follow up gastroscopy was performed between 10 and 34 weeks after reconstitution of bile flow (follow up endoscopy was only performed when medically indicated). Three of the five patients received stents into the common bile duct and two patients had a Whipple operation, one due to a tumour in the head of the pancreas and one due to a cholangiocarcinoma. The patient who had the Whipple operation due to a tumour in the head of the pancreas could not be included in the follow up analysis because duodenal biopsies could not be obtained after complete duodenopancreatectomy. In the patient with cholangiocarcinoma, only a partial duodenopancreatectomy was performed so duodenal biopsies were obtained in the follow up endoscopy.
At follow up endoscopy, bilirubin and bile acid plasma concentrations had normalised in all patients and three of the four patients showed increased expression of ASBT mRNA compared with the baseline value obtained. These three follow up patients with bilirubin (bile acid) plasma concentrations of 373 (431), 170 (204), and 115 (170) μmol/l before endoscopic intervention showed an increase in ASBT expression from 73 to 103, 29 to 68, and 71 to 157 after reconstitution of bile flow, respectively. One patient, who had a bilirubin (bile acid) level of 65 (32.2) μmol/l before intervention and 24 (14.5) μmol/l after reconstitution of bile flow showed an ASBT expression of 130 before intervention, which decreased slightly to 122 after reconstitution of bile flow (n = 4). The difference in ASBT expression before (76 (18)) and after (113 (18)) reconstitution of bile flow for these four follow up patients was not significant (paired t test; p = 0.15).
Immunohistochemistry of ASBT
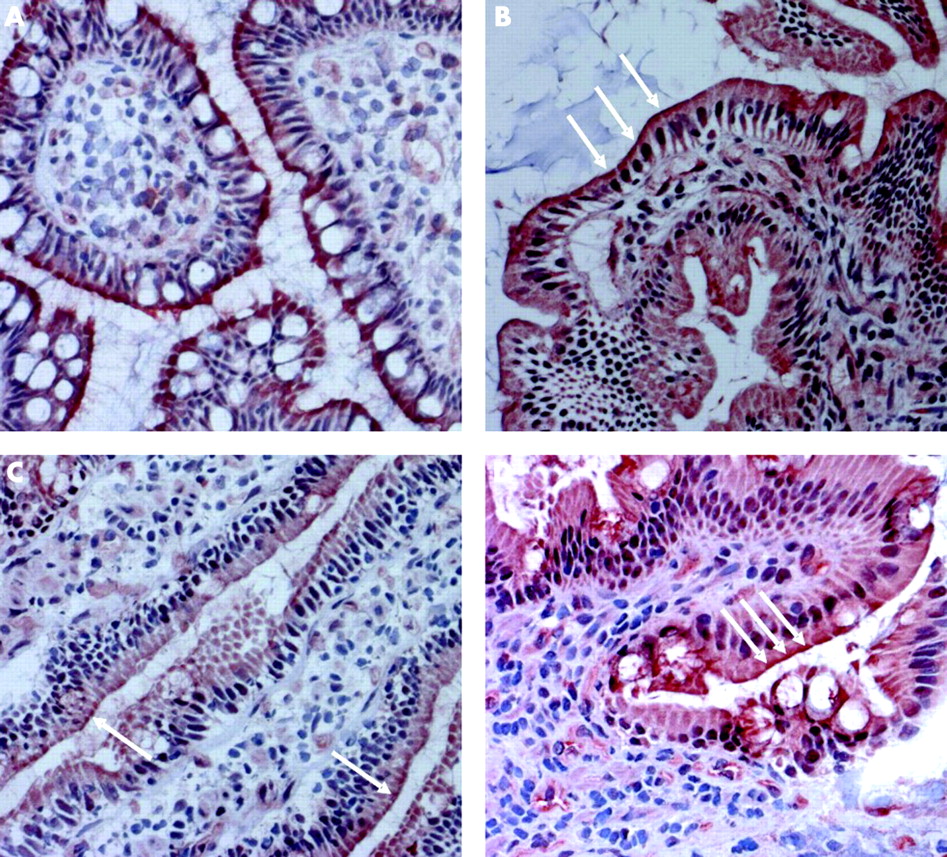
Immunohistochemistry analysis in healthy subjects revealed clear staining of ASBT in ileal and duodenal mucosa (fig 4A and B, respectively). Staining of duodenal mucosa was clearly less intense than that from ileal tissue. In the duodenal mucosa of icteric patients, staining for ASBT was almost completely abolished (fig 4C). After reconstitution of bile flow, a more intense staining was found in the same tissue (fig 4D).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A–D) Immunohistochemical localisation of apical sodium dependent bile acid transporter (ASBT) protein on the apical membrane of the ileum and duodenum of humans using a polyclonal rabbit antihuman ASBT antibody. Clear staining of ileal (A) and duodenal (B) epithelial cells of control subjects. Staining of duodenal epithelial cells in a patient with obstructive cholestasis was almost abolished before (C) and restored after (D) reconstitution of bile flow (all pictures at 200-fold magnification).
Duodenal expression of MRP2 mRNA and MRP3 mRNA
Additionally, we investigated whether there was also adaptive regulation of MRP2 and MRP3 mRNA expression in the distal part of the duodenum.
Patients with obstructive cholestasis showed significantly lower MRP2 mRNA expression (0.21 (0.02)) compared with control subjects (0.32 (0.03), p = 0.004). After reconstitution of bile flow in four patients with obstructive cholestasis, MRP2 expression increased to 0.28 (0.02) (NS). No difference between patients with obstructive cholestasis and control subjects was observed with regard to MRP3 expression (data not shown).
DISCUSSION
Bile acids undergo extensive enterohepatic and cholehepatic circulation through the coordinated action of several transport proteins in hepatocytes, cholangiocytes, and enterocytes.20 The uptake of bile acids at the apical membrane of enterocytes by the apical sodium dependent bile acid transporter ASBT (SLC10A2) reflects an important mechanism for enterohepatic circulation of bile acids. Human ASBT is an efficient transport system for conjugated and unconjugated bile acids.3 In the present study we were able to show that human ASBT mRNA and protein are expressed in the duodenum of the human intestine but to a lesser extent than in the terminal ileum. In addition, significant downregulation of duodenal ASBT mRNA expression was demonstrated in patients with complete or near complete obstruction of bile duct compared with control subjects. Finally, restoring bile flow to the duodenum increased ASBT mRNA expression in a subpopulation of patients with bile duct obstruction.
Downregulation of human duodenal ASBT mRNA may be compared with the adaptive response of bile acid transport proteins during obstructive cholestasis in the terminal ileum, the kidney, as well as in the cholangiocytes of the rat.11,21 In addition to downregulation of asbt in the terminal ileum, the intestinal absorption rate of taurocholate in bile duct ligated rats was lower and the absorption rate was inversely correlated to serum bile acid concentrations compared with sham operated rats.11 Patients with mutations in the ASBT gene can suffer from congenital diarrhoea and steatorrhoea, which is explained by an interrupted enterohepatic bile acid circulation.4 Further clinical data indicate that in patients with primary sclerosing cholangitis, biliary enrichment of UDCA decreases with increasing cholestasis.22 Moreover, in patients with bile duct obstruction and external biliary drainage, intestinal absorption of UDCA was decreased, before relief of cholestasis.23 These data suggest that in humans, intestinal bile acid absorption is reduced during obstructive cholestasis.
What factors influence ASBT gene expression in the enterocyte are largely unknown. Animal studies suggest that luminal bile acids may be one such regulatory factor. Reduction of intestinal bile acid concentration during fasting24 or after biliary diversion25 or bile duct ligation26 is accompanied by a decrease in ileal bile acid transport. Stravitz et al demonstrated that increasing intestinal bile acid concentrations by cholic acid feeding leads to an increase in the sodium dependent transport rate.10 Furthermore, adaptive induction of asbt mRNA levels and transporter protein as well as increased asbt transport activity by cholic acid fed rats was documented.10
In contrast with these observations, other authors have demonstrated in rats that the ileal taurocholate absorption rate correlated inversely to serum bile acid concentrations after bile duct ligation and after biliary diversion compared with sham operated animals.11 The authors concluded that cholestasis leads to decreased, and biliary diversion to increased, active ileal absorption of taurocholate, in which the systemic bile acid load seems to be the decisive factor. In the kidney, bile acid excretion in urine increased progressively in both cholestatic animal models but also in clinical cholestatic disorders.21,27 In Wistar rats, a marked reduction in asbt protein expression in microsomal membrane fractions from whole kidney after CBDL was observed, resulting in reduced levels of asbt expression on the luminal membrane of the proximal tubule of the kidney, a change that is associated with a diminished capacity to reabsorb bile acids from the glomerular filtrate.21 In the same study, upregulation of Mrp2 protein expression on the apical membrane of the rat renal proximal tubule was shown. MRP2 is associated with an increased ability to excrete divalent organic anions such as bile salt sulphates and glucuronides (bilirubin) that accumulate during cholestasis and would therefore facilitate extrahepatic pathways for bile acid and bilirubin excretion during cholestasis. This adaptive response of asbt and Mrp2 in the kidney of the rat does not seem to be regulated by luminal bile acids.
Here we showed that ASBT mRNA expression correlated inversely with bilirubin as well as with bile acid plasma concentrations, which were both used as markers for obstructive cholestasis.
Bile acids are synthesised from cholesterol in the liver; their production is a major mechanism of cholesterol elimination and important for the maintenance of cholesterol homeostasis.28 Downregulation of ASBT expression in obstructive cholestasis might therefore be of clinical interest. In a previous publication, it was shown that inhibition of ileal bile acid transport with SC-435, a competitive inhibitor of ASBT, lowered plasma cholesterol levels: (1) by inactivating the hepatic farnesoid x receptor and (2) by stimulating cholesterol 7α-hydroxylase (CYP7A1).29 This cytochrome is the rate limiting enzyme of chenodeoxycholic acid synthesis. It is located in the endoplasmic reticulum of the hepatocyte. In patients with biliary obstruction, a rise in serum 7α-hydroxycholesterol was observed after biliary drainage.30 The 7α-hydroxylation rate was significantly lower for patients with obstructive cholestasis compared with healthy subjects.31 These data suggest that in the case of a substantial bile acid malabsorption, the activity of 7α-hydroxylase in the liver and synthesis of bile acids is increased.
Other authors suggest that inhibition of ASBT with SC-435 reduces low density lipoprotein cholesterol and ApoB by enhanced plasma clearance of low density lipoprotein ApoB.6,9 Treatment of Caco-2 cell monolayers with 25-hydroxycholesterol significantly inhibited Na+ dependent 3H-taurocholate uptake. Inhibition in hASBT activity was associated with reduction in both the level of hASBT mRNA and its promoter activity.32 asbt protein expression did not change in rats but rose by 31% in rabbits when they were fed with cholesterol (cholest-5-en-3β-ol).33 Administration of SC-435 to apo E−/− mice lowered serum total cholesterol by 35% and reduced aortic root lesion area by 65%. The authors concluded that specific inhibition of abst could be a novel therapeutic approach for treatment of hypercholesterolaemia, resulting in a decreased risk for atherosclerosis.7 Taken together, all of these data suggest that expression of ASBT in the enterocyte seems to be an important component in cholesterol homeostasis and lipid metabolism in humans.
Intracellular factors that influence ASBT gene expression remain largely unknown. One potential factor might be activation of nuclear receptor PPARα, a ligand activated transcription factor that regulates expression of a number of genes involved in peroxisomal and mitochondrial β-oxidation of fatty acids. PPARα binds as a heterodimer with the retinoid X receptor to a peroxisome proliferator response element located in the promoter region of target genes. PPARα was shown to play a critical role in the adaptive response to fasting in mice.34–36 PPARα also influences bile acid composition by induction of the sterol 12α-hydroxylase, which acts at a branch-point in the bile acid synthetic pathway by catalysing the conversion of 7α-hydroxy-4-cholesten-3-one to 7α,12α-dihydroxy-4-cholesten-3-one. This reaction determines the ratio of cholic acid to chenodeoxycholic acid.37 In Hepa 1c1c7 cells, which were transiently transfected with an expression plasmid for PPARα and retinoid X receptor α and a luciferase reporter construct containing copies of the peroxisome proliferator response element from the rat ACOX gene, inhibition of PPARα reporter gene expression was shown with increasing concentrations of chenodeoxycholic acid in the presence or absence of Wy-14,643 ( = PPARα ligand).38 It was concluded that during certain pathophysiological states, where intracellular bile acid concentrations might be elevated, effects on PPARα dependent target gene regulation are possible.38 Recently, PPARα was shown to transactivate the human ASBT gene.39 Incubation of human hepatoma HepG2 cells with chenodeoxycholic acid resulted in significant induction in PPARα mRNA levels. In addition, hPPARα gene expression was upregulated by taurocholic acid in human primary hepatocytes.40 However, no difference between patients with obstructive cholestasis and control subjects was observed with regard to duodenal PPARα mRNA expression in our study.
In a previous study, Dietrich et al demonstrated that obstructive cholestasis promotes downregulation of intestinal MRP2 protein expression in rats and humans.15 The reduction was correlated with duration of cholestasis and was reversible after reconstitution of bile flow. In the same study, in patients with obstructive cholestasis, a non-significant decrease in MRP2 mRNA expression was detected compared with control subjects. However, the authors observed that decline of intestinal rat Mrp2 mRNA occurred more slowly and gradually than downregulation of its protein expression. Our results showed a significant downregulation of MRP2 mRNA expression compared with control subjects and a non-significant increase after reconstitution of bile flow in four patients. One explanation for the downregulation of MRP2 gene expression in our study compared with Dietrich and colleagues15 might be higher plasma bilirubin concentrations in our patients with obstructive cholestasis. Another reason might be duration of cholestasis, which was not clearly stated in both studies and therefore may have been different. Other factors such as food derived compounds or substrates might also be involved in the regulation of gene expression. For example, it was shown that grapefruit and orange juice inhibited the transport by MRP2,41 components of our daily diet, such as the flavenoid epicatechin in tea,42 chrysin, and its metabolites are substrates of MRP2.43 Drugs are unlikely to have contributed to the different MRP2 mRNA expression in the present study and the study performed by Dietrich and colleagues15 because patients with drugs, which are known to affect MRP2 expression, were excluded from both studies. Finally, our data support the results of Dietrich and colleagues15 which demonstrated an adaptive downregulation of duodenal MRP2 in patients with obstructive cholestasis.
MRP3 is expressed on the basolateral membrane in the proximal small bowel13 and is capable of transporting bile salts, including taurocholate, glycocholate, taurochenodeoxycholate-3-sulphate, and taurolithocholate-3-sulphate.14 This suggests that MRP3 might be an important transporter in the enterohepatic circulation of bile acids. However, in the present study, we did not observe any difference in human duodenal MRP3 gene expression in patients with obstructive cholestasis compared with control subjects. Similarly, no compensatory upregulation of human duodenal MRP3 expression in cholestatic patients was previously described.15
In conclusion, human ASBT mRNA is expressed in the small intestine, predominantly in the terminal ileum but also, to a lesser extent, in the duodenum. Adaptive downregulation of ASBT and MRP2 mRNA in the duodenum can be observed in patients with obstructive cholestasis compared with control subjects. Furthermore, duodenal ASBT mRNA expression levels inversely correlated with bilirubin and bile acid plasma concentrations in patients with obstructive cholestasis as well as in control subjects. This adaptive gene regulation may represent a mechanism preventing the accumulation of hepatotoxic bile acids in cholestasis. After relief of cholestasis, an increase in ASBT mRNA was observed in three of four patients.
Acknowledgments
We are very thankful to Uschi Behrens, Ursula, Dürmüller and Gudrun Schwertfeger for their excellent technical assistance.
REFERENCES
Footnotes
-
Published online first 8 September 2005
-
Conflict of interest: None declared.