Article Text

Abstract
The interface between luminal contents and intestinal epithelium constitutes the largest area of interaction between the host and the environment. There is now strong evidence that the gene product of the multidrug resistant pump (MDR) plays a critical role in host-bacterial interactions in the gastrointestinal tract and maintenance of intestinal homeostasis. This review highlights the efflux mechanism in the intestinal epithelium which is mediated by the multidrug resistant pump, also known as P-glycoprotein 170. Current studies promise to provide further insights into the contribution of the MDR1 gene in the pathogenesis of inflammatory and malignant disorders of the gastrointestinal tract.
- multidrug resistance 1 gene
- P-glycoprotein 170
- drug resistance
- inflammatory bowel disease
- colorectal cancer
- MDR, multidrug resistance
- IBD, inflammatory bowel disease
- NFκB, nuclear factor κB
- IκB, inhibitory protein κB
- SNP, single nucleotide protein
- PBL, peripheral blood lymphocytes
- CYP3A, cytochrome P450 3A
Statistics from Altmetric.com
- multidrug resistance 1 gene
- P-glycoprotein 170
- drug resistance
- inflammatory bowel disease
- colorectal cancer
- MDR, multidrug resistance
- IBD, inflammatory bowel disease
- NFκB, nuclear factor κB
- IκB, inhibitory protein κB
- SNP, single nucleotide protein
- PBL, peripheral blood lymphocytes
- CYP3A, cytochrome P450 3A
SUMMARY
The interface between luminal contents and intestinal epithelium constitutes the largest area of interaction between the host and the environment. Critical to this interface is the ability to allow transcellular permeation of essential luminal molecules such as nutrients, but at the same time to exclude or counter potentially harmful substances in the intestinal lumen. The cells of the mucosal immune system are protected from the large antigenic load in the gut lumen by a single layer of epithelial cells. Studies involving transgenic mice models have highlighted the critical role of the mucosal microflora in the pathogenesis of mucosal inflammation and the importance of the epithelial cell barrier function in providing protection against the extensive stimulation of mucosal immune system by the microflora.
Together with the activity of xenobiotic metabolising enzymes in the epithelium, efflux mechanisms have become the subject of considerable interest in recent studies of gut mucosal defence and pharmacokinetics of drugs. This review concentrates on the efflux mechanism in the intestinal epithelium which is mediated by the multidrug resistant pump (MDR1), also known as P-glycoprotein 170. Recent data strongly implicate MDR1 as a determinant of homeostatic interactions between bacteria and host in humans. Current studies promise to provide further insights into the contribution of the MDR1 gene in the pathogenesis of inflammatory and malignant disorders of the gastrointestinal tract.
INTRODUCTION
The multidrug resistance pump (P-glycoprotein) is a 170 kDa phosphorylated and glycosylated plasma membrane protein belonging to the ATP binding cassette superfamily of transport proteins encoded by the multidrug resistance genes (MDR), first described by Juliano and Ling in 1976.1 In humans, two MDR genes, MDR1 and MDR3 (also called MDR2) have been described; in rodents, three (mdr1a, mdr1b and mdr2).2–4 Only the MDR1 gene in humans and mdr1b and mdr1a genes in rodents appear to be involved in drug transport and the development of drug resistance.5 The human MDR3 and murine mdr2 genes encode a P-glycoprotein that does not seem to have a role in drug transport in the intestinal epithelium but has a role in the secretion of phosphatidylcholine into bile in the biliary tract.6 In humans, mutations in the MDR3 genes are linked to the development of progressive familial cholestasis.7,8
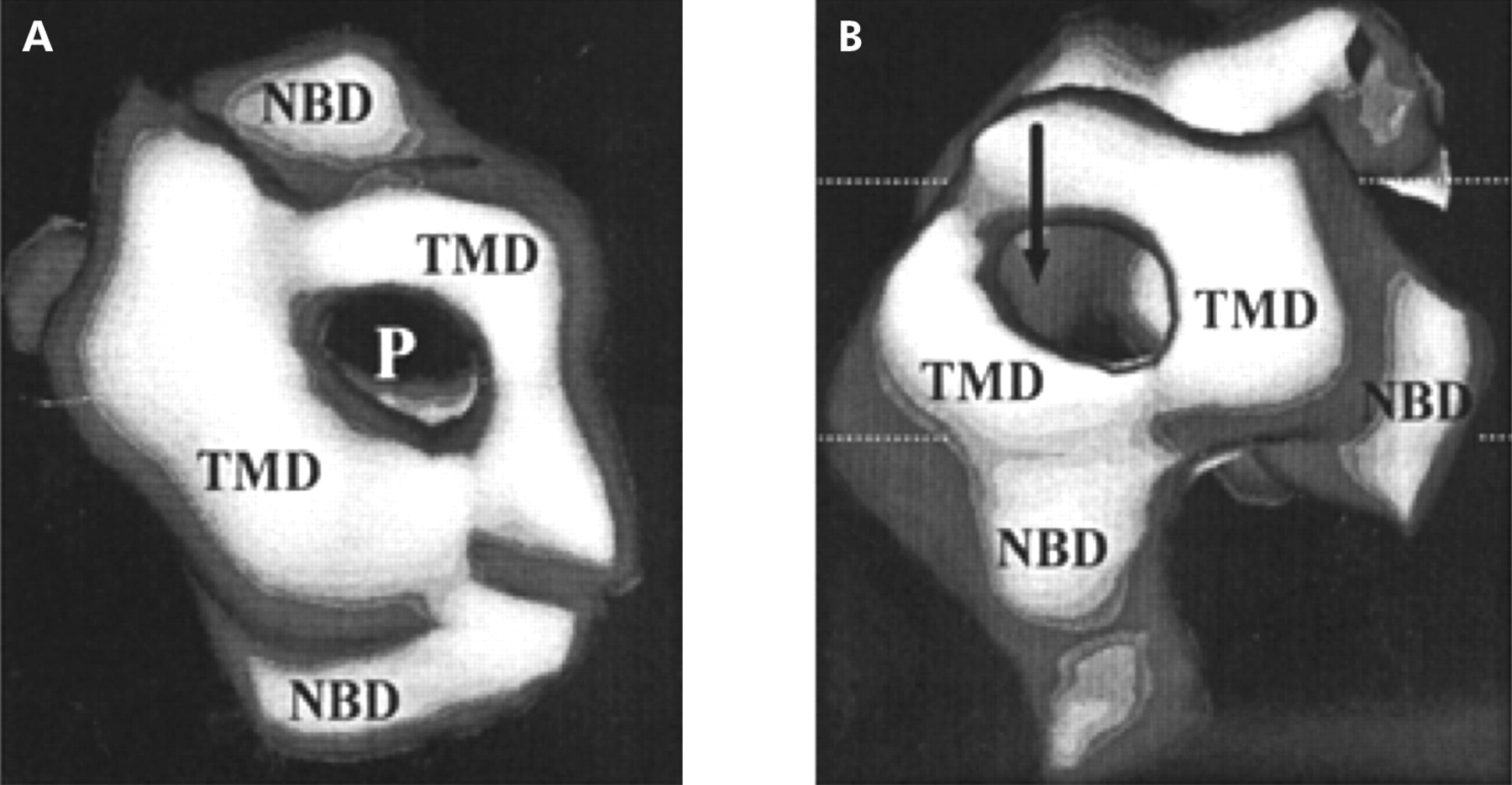
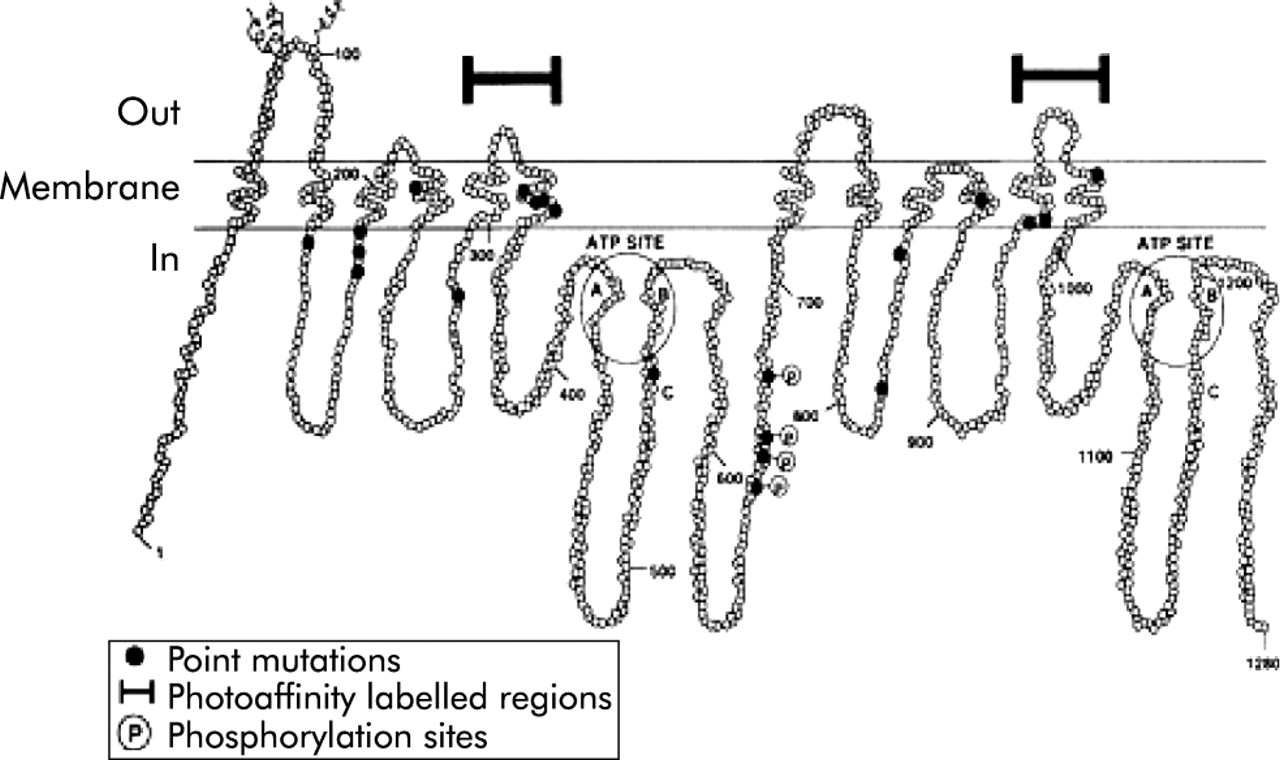
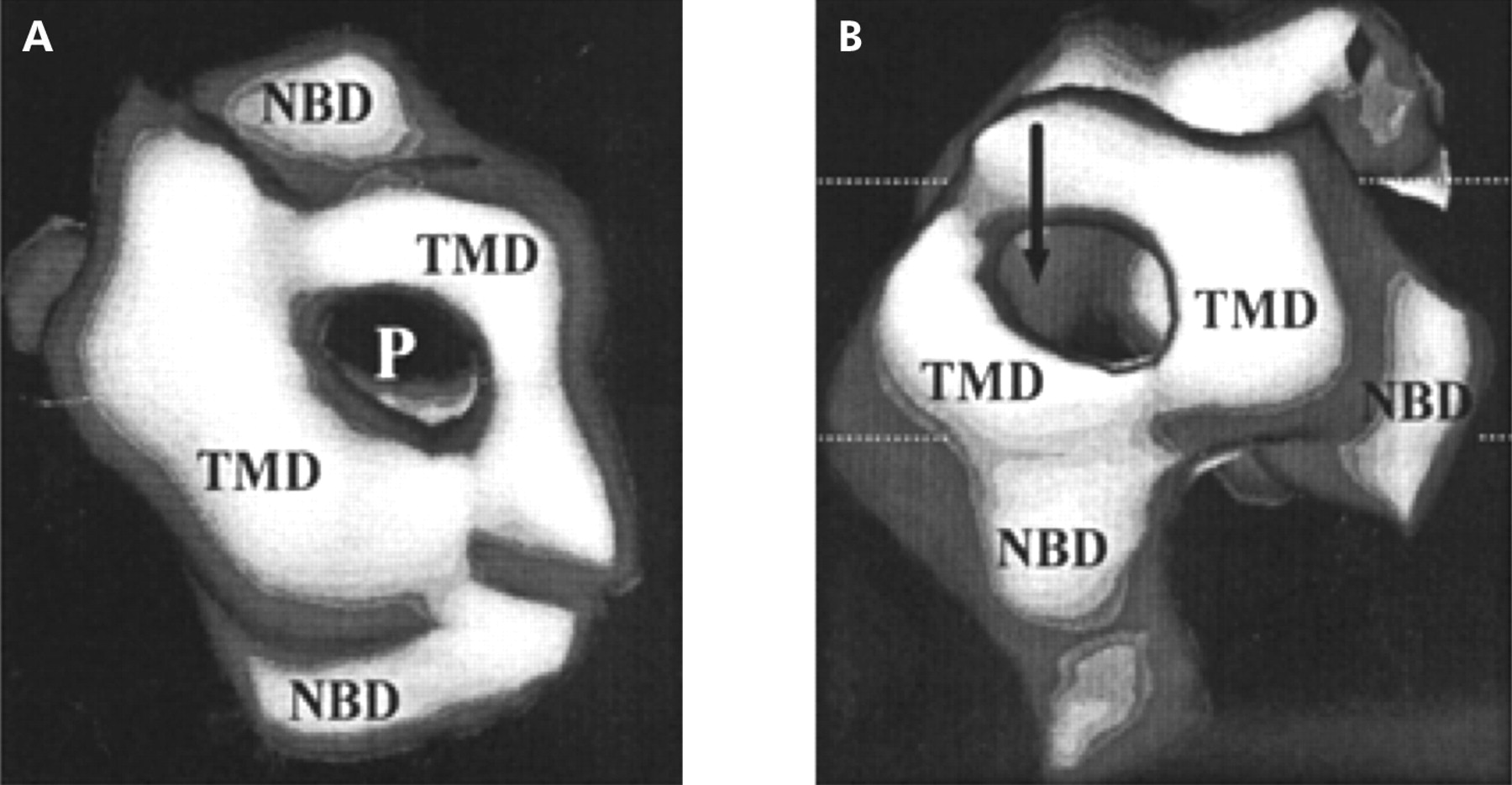
P-glycoprotein is a transmembrane protein which is 1280 amino acids long and consists of two homologous halves of 610 amino acids joined by a flexible linker consisting of 60 amino acids. Each half has an N terminal hydrophobic domain containing six transmembrane domains followed by a hydrophilic domain containing a nucleotide binding site. The nucleotide binding sites can bind ATP and its analogues and both halves are essential as inactivation of either site inhibits substrate stimulated ATPase activity (figs 1, 2 ). However, the two sites are likely to be functionally independent and cleavage probably occurs at one site at a time.9

Two dimensional hypothetical structure of the P-glycoprotein 170 pump.
{kind=link}
{kind=link}
Three dimensional hypothetical structure of P-glycoprotein viewed from above (A) and below (B). NBD, nuclear binding domain; P, pore; TMD, transmembrane domain.
In the human gastrointestinal tract, P-glycoprotein is found in high concentrations on the apical surfaces of superficial columnar epithelial cells of the colon and distal small bowel. High levels of P-glycoprotein are also found on the apical surfaces of epithelial cells in small biliary ductules, small ductules of the pancreas, proximal ductules of the kidneys, and adrenal glands.10 P-glycoprotein is richly expressed on the subapical surface of the epithelium of the choroid plexus of the brain (which forms the blood-cerebrospinal fluid barrier) as well as the luminal surface of the endothelium of the blood capillaries of the brain (blood-brain barrier).11–14 In the haemopoietic system, peripheral blood mononuclear cells, macrophages, natural killer, dendritic cells, and T and B lymphocytes all express P-glycoprotein at varying levels.15
In the gastrointestinal tract, there is regional variation in P-glycoprotein expression. Various methodologies have been used to investigate this (table 1). Within the small intestine there is clear evidence demonstrating that P-glycoprotein is maximally expressed in the epithelial cells of the ileum with a gradual decrease proximally into the jejunum, duodenum, and stomach. Variations of P-glycoprotein expression across the length of the colon are less well defined.16
Regional variation in P-glycoprotein expression in humans and rodents
THE ROLE OF P-GLYCOPROTEIN
Host-bacterial interactions
The available literature suggests that P-glycoprotein acts as a transmembrane efflux pump which removes drugs from the cell membrane and cytoplasm. However, the normal physiological function of P-glycoprotein in the healthy gastrointestinal tract remains under investigation and may be complex. Expression of P-glycoprotein on the luminal surfaces of epithelial cells of the small and large intestines, biliary ductules, and proximal tubules of the kidney suggest a role in decreasing17,18 absorption from the gut19 and secretion of endogenous and exogenous hydrophobic amphipathic toxins.20
Perhaps the greatest insight into the physiological importance of P-glycoprotein in the gastrointestinal tract has come from the description of the phenotype of genetically engineered mice lacking the mdr1a gene. The investigators had been stimulated to develop this model by the finding that the MDR1 gene is present in a region of the human genome (7q21.1) that may harbour a disease gene involved in susceptibility to inflammatory bowel disease (IBD).21 Panwala et al demonstrated that spontaneous colitis developed in these mdr1a knockout mice when maintained under specific pathogen free conditions.22 This was reversed and prevented by administration of antibiotics, suggesting that the intestinal flora was necessary in the initiation and perpetuation of colitis in this model. Other studies have confirmed that these mice have normal viability, fertility, immunology, and a range of biochemical parameters.11,23 Together with the protective effect of antibiotics, these observations imply that loss of the xenobiotic efflux mechanism underlies the development of colitis in this model. Bone marrow transfer studies involving wild-type donor cells into mdr1a deficient recipients demonstrated that colitis develops in these mice because of the mdr1a deficiency in the epithelial cells rather than in lymphoid or myeloid cells. Colitis associated with mdr1a deficiency has been regarded as a model of ulcerative colitis with the presence of long dysregulated crypts, crypt abscesses, and superficial mucosal inflammation. However, transmural infiltration of B and T cells was also observed, not dissimilar to the histological appearances of Crohn’s disease.
Susceptibility to the development of colitis increased as the mice (mdr1a−/−) aged.22 The gradual acquisition of commensal luminal organisms in the background of defective efflux capacity of epithelial cells may be the reason, although clearly other effects of aging may be pertinent. Sequential faecal analyses over time are necessary to investigate this hypothesis.
In addition, a recent study has added further intriguing insight into the interaction between P-glycoprotein and the gut flora. In mdr1a−/− mice, infection with Helicobacter bilis accelerated the development of colitis while Helicobacter hepaticus infection delayed the development of spontaneous disease.24Helicobacter induced IBD in mdr1a−/− mice may be mediated via a family of bacterial toxins (for example, cytolethal distending toxins) which are virulence factors in certain Helicobacter infections. However, it is important to emphasise that these mice developed spontaneous colitis independent of Helicobacter infection. These observations suggest that specific luminal bacteria may critically influence the development of colitis and its progression in genetically susceptible animals. Both Helicobacter bilis and hepaticus have been shown to induce colitis in mice lacking T and B cells (SCID mice).24–27
Moreover, these findings are consistent with the hypothesis that mdr1a deficient mice developed mucosal inflammation not because of increased permeability but as a result of an increase in bacterial activation of epithelial cells. This effect might be pro or anti-inflammatory and again the identity of the bacterium involved may be the critical determinant. It has been demonstrated that bacterial flagellin signalling through the TLR5 receptor can lead to increased nuclear factor κB (NFκB) expression in epithelial cells.28 On the other hand, Neish et al demonstrated that certain non-pathological organisms can lead to a block in the ubiquitination of inhibitory protein κB (IκBa) (necessary for IκBa degradation) and thus a subsequent block in NFκB translocation to the nucleus.29 In either of these cases, if these events are caused by bacteria entering the epithelial cells, it is possible to envisage how the function of the epithelial cells can be influenced by a defective efflux transporter and the bacterial contents of the lumen.
Pharmacokinetic effects
The effect of P-glycoprotein on the pharmacokinetics of drugs has been well demonstrated in two studies.30,31 In mdr1a−/− mice, systemic bioavailability for the substrates digoxin and paclitaxel was markedly increased compared with wild-type mice following oral administration. Similarly, faecal levels of these drugs were reduced following intravenous administration. The range of substrates transported by P-glycoprotein is enormous and includes a variety of structurally and pharmacologically distinct hydrophobic compounds (table 2).
List of P-glycoprotein substrates
“The range of substrates transported by P-glycoprotein is enormous and includes a variety of structurally and pharmacologically distinct hydrophobic compounds”
There is accumulating evidence that interactive alliances also exist between xenobiotic metabolising enzymes and P-glycoprotein. The best defined alliance is between cytochrome P450 3A (CYP3A) and P-glycoprotein. These proteins colocalise in the small intestinal epithelial cells and have a considerable overlap in substrate specificities.32 As P-glycoprotein can influence the intracellular concentrations of many CYP3A substrates, it may also affect the availability of those substrates to CYP3A and thereby the extent of CYP3A metabolism of those substrates.
REGULATION OF P-GLYCOPROTEIN
Regulation of MDR1 gene expression and P-glycoprotein activity is unexpectedly complex and far from being understood. The transcriptional regulators of the human multidrug resistance gene are discussed in detail in the review by Labialle and colleagues.33 P-glycoprotein dependent drug transport activity depends on the level of expression of the MDR1 gene as well as on the functionality of the MDR1 encoded P-glycoprotein. It is clear that intestinal P-glycoprotein expression and function shows wide interindividual differences, influenced by both environmental and genetic factors.
Pharmacological regulation of P-glycoprotein
Shapiro et al showed that P-glycoprotein mediated drug transport was stimulated by the antihypertensive drug prazosin and the hormone progesterone.34 Rifampicin has also been shown to induce P-glycoprotein expression and underlies the mechanism responsible for reduced digoxin levels during concomitant therapy.35 In healthy male volunteers, the oral bioavailability of digoxin decreased by 30% and intestinal P-glycoprotein levels were induced 3.5-fold during rifampicin therapy. Induction appears to take place at the transcriptional level as MDR1 mRNA levels are also elevated after treatment with inducer drugs.36,37 It has been suggested that induction of MDR1 is mediated by the pregnane X receptor (which is also responsible for induction of CYP3A) which binds to PXR response elements situated upstream in the MDR1 gene.38
“The influence of corticosteroids on expression of P-glycoprotein has been an area of considerable recent interest as a possible mechanism for corticosteroid resistance in chronic inflammatory diseases”
The influence of corticosteroids on expression of P-glycoprotein has been an area of considerable recent interest as a possible mechanism for corticosteroid resistance in chronic inflammatory diseases. High levels of expression of P-glycoprotein in the adrenal glands and endometrium suggest a role in the transport of steroid related compounds. In addition, these compounds are all substrates of P-glycoprotein. Nevertheless, at the present time, the available literature is complex and inconsistent (table 3).
Studies on the effects of corticosteroids on multidrug resistance/P-glycoprotein 170 expression
In mouse hepatoma cell cultures treated with dexamethasone, Zhao et al demonstrated induction of mdr1 and mdr3 but not mdr2 transcripts. Similarly, MDR1 mRNA levels were elevated in a dexamethasone treated human hepatoma line. This effect was not seen in a non-hepatoma cell line (HeLa).14 Several studies have investigated the effect of corticosteroids in animal models. In a study examining expression of CYP3A and mdr in rat liver, dexamethasone was found to induce mdr2 expression only in male livers. Curiously, mdr1a and 1b were not induced in either sex.39 Lin et al showed that dexamethasone induces P-glycoprotein in the intestines and liver of rats with the functional effect of decreased plasma substrate level (indinavir).40 However, Demeule et al found no increase in intestinal expression. In these animals, expression was upregulated in the lungs and liver whereas an opposite effect was observed in kidney tissue.41
Although P-glycoprotein expression may be influenced by corticosteroids or other pharmacological agents, recent data illustrate that the level of expression may not necessarily correlate with its function. Murakami et al demonstrated that although increased P-glycoprotein expression was evident in rats with induced acute renal and liver failure, in vivo P-glycoprotein activity was significantly depressed. Plasma from these diseased rats (with renal and liver failure) demonstrated a significantly greater inhibitory effect on P-glycoprotein function (in a Caco-2 cell line) compared with dexamethasone treated rats.42 It is hypothesised that in disease states, endogenous related compounds such as corticosterone (a P-glycoprotein inhibitor) may participate to suppress MDR1 activity.
Another intriguing aspect of the pharmacological regulation of expression of P-glycoprotein is seen in the multidrug resistance phenomenon. This is observed in tumour cells which, following initial contact with one anticancer agent, become rapidly resistant to other unrelated agents. This phenotype is mainly due to overexpression of the MDR1 gene. One possible mechanism for this may be through gene rearrangement. Mickley et al found that in several drug resistant cancer cell lines as well as samples from two leukaemic patients who had developed drug resistance, gene rearrangement had occurred resulting in activation or increased expression of MDR1.43 Although a feasible hypothesis, there is no evidence to support selection of aberrant clones of the MDR gene as the cause of the development of multidrug resistance.44,45
The above studies suggest that regulation of MDR1 gene expression is likely to be gene, cell, and perhaps gender specific. Furthermore, future work in this area will have to consider not only expression but also the function of P-glycoprotein.
Genetic influences on MDR
The MDR1 gene lies on chromosome 7q21.1. Several polymorphisms of the gene have been characterised (table 4).46 Hoffmeyer et al first described a single base polymorphism in exon 26 (C3435T) and suggested that this variant influences P-glycoprotein expression in the duodenum and P-glycoprotein activity.47 In this study, healthy individuals homozygous for this variant (TT genotype) had significantly lower duodenal expression of P-glycoprotein and higher serum digoxin levels. Moreover, evaluation of maximum plasma concentrations during steady state digoxin administration showed a statistically significant increase (38%) in the homozygous TT genotype compared with the CC genotype.
Single nucleotide polymorphisms (SNPs) in the MDR1 gene
It is particularly noteworthy that the MDR1 C3435T allelic variant, a single nucleotide polymorphism (SNP), is non-coding and does not affect amino acid sequence change. Although a number of coding mutations within the MDR1 gene (table 4) have been identified, none of these variants is in strong linkage disequilibrium with the C3435T variant.46,48,49 The observed functional effect ascribed to the C3435T variant may none the less reflect linkage disequilibrium with a polymorphism elsewhere in the genome that modifies MDR1 expression and function. However, at present this hypothesis requires further detailed studies of the gene. In parallel studies, Kimchi-Sarfaty et al demonstrated that there were no functional consequences of the five commoner amino acid altering SNPs (including 2677 SNP) in this gene, on both cellular localisation and transport function of P-glycoprotein.50
“Given the likely complicated interactions with the cytochrome P450 system and the presence of other poorly characterised transporter systems, the relationship between the function of P-glycoprotein and drug substrate levels measured in vivo or in vitro is debatable”
This exonic variant (C3435T) has received considerable attention although controversy still exists regarding the functional effect of this polymorphism. Of the five published studies investigating the effect of this polymorphism on expression of MDR1, three studies reported decreased expression and two studies increased expression in association with this SNP (table 5). Kim et al showed that the MDR 1*2 haplotype (G2677T/C3435T) was associated with increased P-glycoprotein function in vitro and low concentrations of a known substrate, fexofenadine.51 Fellay et al demonstrated that the MDR1 3435 TT genotype was associated with low expression of MDR1 transcripts and P-glycoprotein expression in peripheral blood mononuclear cells with low plasma levels of nelfinavir and efavirenz (antiretroviral agents).52 Hitzl et al showed that in CD56+ cells (natural killer cells), the MDR1 3435 TT genotype was associated with lower P-glycoprotein function and expression compared with the CC genotype.53 In another study, Nakamura et al investigated the effect of the C3435T mutation on the expression level of MDR1 mRNA in duodenal enterocytes of healthy Japanese subjects54 and found significantly elevated levels of transcripts in those patients with the TT compared with the CC allele. Given the likely complicated interactions with the cytochrome P450 system and the presence of other poorly characterised transporter systems, the relationship between the function of P-glycoprotein and drug substrate levels measured in vivo or in vitro is debatable. This consideration and the differences in study designs may explain these apparently contradictory results. Moreover, there may be cell specific differences in regulation. There is a clear need for further well designed studies using in vivo and ex vivo systems to reconcile these differences.
Cell populations and transporter substances used in functional studies of MDR1 3435TT genotype
The MDR1 gene lies within the region of chromosome 7q which a recent two stage genome wide search has implicated as a putative susceptible locus for the development of IBD.21 The effects of polymorphisms within this gene on intestinal expression suggest that MDR1 gene and function may be important determinants of the development of IBD or of disease behaviour. This hypothesis is even more attractive given the effects of targeted gene deletion in mice, as described previously.22 Current investigations will provide further insights into this area.
MDR1 AND INFLAMMATORY BOWEL DISEASE
The finding of the development of spontaneous colitis in genetically engineered mdr1a−/− mice might suggest that reduced MDR1 expression or activity in intestinal epithelial cells may be a factor in the pathogenesis of IBD. However, at present, there is surprisingly little published literature to investigate this hypothesis in humans. Two studies have focused on MDR1 expression in IBD.55,56
Farrell et al investigated the hypothesis that altered P-glycoprotein expression on lymphocytes may predict responsiveness to medical therapy.55 Corticosteroids and cyclosporin are both substrates of P-glycoprotein.57 Peripheral blood lymphocytes (PBL) from patients with active Crohn’s disease (15 patients) and ulcerative colitis (28 patients) in whom medical therapy had failed and surgical intervention had been necessary were shown to have higher expression of P-glycoprotein compared with those from patients who had inactive disease and required surgery for obstruction (25 patients) or dyplasia (13 patients) (p<0.0001 and 0.0002, respectively). Interestingly, in ulcerative colitis, there was no significant difference when PBL P-glycoprotein expression was compared between patients requiring colectomy for failed medical treatment and the group with active ulcerative colitis (p=0.6). Farrell et al went on to demonstrate that by blocking the action of P-glycoprotein on human cell lines, higher intracellular levels of cyclosporin and corticosteroids were achieved58 and speculated that this may be a potential target for pharmacotherapy.
In Farrell’s initial study,55 MDR1 expression of the intestinal epithelial cells in inflamed areas of colon in patients with active ulcerative colitis (n=6) was not different from expression in non-inflamed areas. Comparisons were not made with normal controls and it remains to be evaluated whether altered levels of MDR1 expression in the colon in humans lead to increased susceptibility to the development of IBD (as observed in studies involving mice models lacking the mdr1a gene). Although, intraepithelial lymphocytes/colonic epithelial expression of MDR1 correlated significantly with PBL expression, overall expression in the control group was in fact similar to groups of patients with inactive ulcerative colitis/Crohn’s disease.
With respect to MDR1 overexpression as a factor in corticosteroid resistance, this has also been studied in other conditions. In patients with glucocorticoid resistant asthma, P-glycoprotein expression on PBL was not found to be increased compared with those with responsive or mild asthma.59 In patients with rheumatoid arthritis treated with glucocorticoids, expression of P-glycoprotein on PBL was increased compared with those without therapy.60 Patients with rheumatoid arthritis without corticosteroid therapy did not demonstrate higher expression of MDR1 and this again may reflect a secondary effect to corticosteroids. In a subset of patients with IBD not requiring surgery (n=72), multiple regression analysis of several clinical variables showed that only concurrent corticosteroid therapy was significantly associated with high PBL MDR1 expression (p=0.001).55
Intraepithelial MDR1 expression and function were suggested to be lower in ulcerative colitis compared with Crohn’s disease and healthy controls
There are several caveats to these studies. Firstly, circulating lymphocytes consist of different subsets of lymphocytes which express MDR1 at varying levels.61 Indeed, within the gastrointestinal tract it is now well established that lymphocytes homing to the gut associated mucosal lymphoid tissue are phenotypically distinct from circulatory lymphocytes. It is also pertinent that Yacyshyn et al have demonstrated that the intraepithelial, lamina propria, and PBL all demonstrate different levels of MDR1 expression and activity in ulcerative colitis, Crohn’s disease, and healthy controls. In this study, CD3 and CD8 lymphocytes were also present in different proportions in these groups of subjects. These subsets were also shown to have intrinsically different levels of P-glycoprotein function (CD8 has higher levels of P-glycoprotein). Overall, intraepithelial MDR1 expression and function were suggested by these investigators to be lower in ulcerative colitis compared with Crohn’s disease and healthy controls.56
Secondly, the effect of corticosteroids (or indeed other treatments used in IBD) on MDR1 expression is not fully established. It remains to be seen whether higher expression of P-glycoprotein reflects a secondary phenomenon to corticosteroid therapy or a primary one, influencing the response to treatment. Finally, as discussed earlier, increased expression does not necessarily imply increased function in diseased states.
MDR1 AND COLORECTAL CANCER
P-glycoprotein expression appears to be highest in tumours originating from normal tissues expressing P-glycoprotein13,10 which include colorectal epithelium. In studies of colorectal carcinoma, expression of P-glycoprotein correlated with pathological grading of tumours, being most intense in well differentiated tumours and low in poorly differentiated ones.62
“Expression of P-glycoprotein correlated with pathological grading of tumours, being most intense in well differentiated tumours and low in poorly differentiated ones”
In view of the presence of intrinsic drug resistance to natural products of anticancer drugs, the high P-glycoprotein expression reported in colorectal cancer, and in vitro studies demonstrating successful P-glycoprotein reversal in colon cancer cell line, specific inhibition of P-glycoprotein has been carried out in clinical trials to determine whether a more favourable response to chemotherapy is achieved. As yet the results have been disappointing and inconclusive. The reasons for this appear to be a combination of drug toxicity of the resistance modifying agents together with an inability to interpret and compare results in different studies as a consequence of the lack of standardisation in the measurement of P-glycoprotein expression.63–65
P-glycoprotein expression has been associated with a poor prognosis in neuroblastoma, sarcoma, and acute myeloid leukaemia.66,67 Its prognostic value in colorectal cancer is less clear. Weinstein et al reported a significant correlation between the presence of P-glycoprotein positive tumour cells at the leading edge of the tumour and an increased incidence of tumour vessel invasion and lymph node metastases.68 Another study reported P-glycoprotein to have prognostic value in Dukes’ stage B colon cancers. However, this study was small (52 patients) and the p value was of marginal significance.69 Three other studies failed to demonstrate a relationship between P-glycoprotein expression and shorter (disease free) survival.70–72
“P-glycoprotein expression has been associated with a poor prognosis in neuroblastoma, sarcoma, and acute myeloid leukaemia”
An intriguing study has identified the MDR1 gene as a target of transactivation by the T cell factor 4/β-catenin complex in early colorectal carcinogenesis.73 Mutational inactivation of the tumour suppressor gene APC results in accumulation of cytoplasmic β-catenin through failure of degradation.74 β-Catenin forms a complex with T cell factor 4 which acts as a transcription factor. Target genes have included c-myc, cyclin D1, matrilysin, T cell factor 1, and peroxisome proliferator activated receptors which are all implicated in the development of colorectal cancer.75–80
Colorectal cell lines that are mutant for APC or β-catenin have high levels of T cell factor mediated transcription which is thought to be the basis of tumorigenesis. Yamada et al performed a large scale comparison of gene expression by involving cDNA microarray technology in the colorectal cell line DLD1 in which T cell factor 4/β catenin transactivation had been suppressed and gene expression data were compared with wild-type DLD1 cells.81 More than 7000 genes were studied, and MDR1 was found to be transcriptionally downregulated after inactivation of T cell factor 4. It is clearly pertinent that the promoter region of the MDR1 gene contains several T cell factor 4 binding sequences. The authors subsequently demonstrated that in adenomatous polyps from patients with familial adenomatous polyposis syndrome, MDR1 expression was significantly increased and correlated with β-catenin.
This study thus provided strong evidence of a link between colorectal cancer and the MDR1 gene. Further functional studies are necessary to establish the functional significance of these findings. It is hypothesised that overexpression of MDR1 may influence apoptotic pathways by secreting endogenous or xenobiotic toxic substances into the lumen or by mediating a more generic antiapoptotic function. The MDR1 gene product, P-glycoprotein, protects the cell against a wide variety of caspase dependent death stimuli including fas ligand, tumour necrosis factor, and ultraviolet irradiation.82
“The MDR1 gene may have an important role in the tumorigenesis of colorectal cancer”
A number of in vitro studies have focused on the effect of the tumour suppressor gene p53 on expression of the MDR1 gene. These report an inhibitory role of the wild-type p53 on the MDR1 promoter region whereas mutant p53 variants act as activators, probably due to loss of its inhibitory role.83–86 Together with the T cell factor elements described earlier, these may represent only a relatively small contribution to the overall complex transcriptional regulation of the MDR1 gene by other transactivating genes. However, it has become apparent that the MDR1 gene may have an important role in the tumorigenesis of colorectal cancer.
CONCLUSION
There is now strong evidence that the gene product of MDR plays a critical role in host-bacterial interactions in the gastrointestinal tract and maintenance of intestinal homeostasis. Perhaps the most important current challenge is to define the physiological role of MDR1 in the healthy intestine. Expression and function of the MDR1 gene in common gastrointestinal diseases is the subject of detailed evaluation, and progress in understanding the pathogenesis of chronic IBD and colorectal cancer may result from these studies.
Moreover, there is real hope that further studies of the MDR 1 gene may lead to direct clinical application, particularly in the field of pharmacogenomics. The possibility of predicting the responsiveness of IBD or colorectal cancer to therapy by measuring P-glycoprotein expression or allelic variation is exciting and appears increasingly realistic. Measurement of this and other subclinical markers may help in optimising the use of current and novel therapeutic agents.55,87
Acknowledgments
This work was supported by the Chief Scientist Office, Scottish Executive (GTH) and Medical Research Council (FMM).