


Abstract
The aryl hydrocarbon receptor (AhR) is a ligand-induced transcription factor that is activated by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and other related compounds, leading to toxicity. There is considerable variation in the response to TCDD among different species, and this may be correlated to differences in the AhR. Variations in the structure of the AhR could result in altered biochemical properties of the receptor, such as ligand affinity or transactivation potential. The difference between the mouse AhR b-1 allele (mAhRb-1) and human AhR (hAhR), in terms of their relative affinity for a photoaffinity ligand (2-azido-3-[125I]iodo-7,8-dibromodibenzo-p-dioxin), was assessed using both in vitro assays and assays performed directly in cell culture. Results revealed that the hAhR has a lower affinity for the photoaffinity ligand compared with mAhRb-1. In contrast with a previous study, we found that a single amino acid (valine 381) in hAhR is responsible for the lower ligand affinity, and mutating this residue to alanine results in restoration of high ligand affinity in hAhR. In vitro ligand binding assays are limited by the low concentrations of protein in the assays, and it is not appropriate to compare ligand affinities of different receptors using this method without performing a competition assay or increasing the protein concentration in the assay. Because of the limitation of the in vitro assay, the relative ligand occupancy of mAhRb-1 and hAhR was compared most effectively within cells, revealing that mAhRb-1 has a 10-fold higher relative ligand affinity in cells, whereas mAhRd has a 2-fold higher relative ligand affinity than hAhR.
Exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and related compounds results in a number of adverse biological effects, such as chloracne, blood and liver disorders and birth defects (Huff et al., 1980). The toxic and carcinogenic effects of TCDD are mediated through the aryl hydrocarbon receptor (AhR), which is a ligand-activated transcription factor. In the cytoplasm, the AhR is present in a complex with a dimer of hsp90, the hepatitis B virus X-associated protein 2 (XAP2), and p23 (as reviewed by Petrulis and Perdew, 2002). Upon binding ligand, the AhR translocates to the nucleus, forms a heterodimer with the aryl hydrocarbon nuclear translocator, and regulates transcription of target genes (Reyes et al., 1992).
There are a number of intra- and interspecies differences in sensitivity to the effects of TCDD (Henck et al., 1981; Chapman and Schiller, 1985; Pohjanvirta et al., 1993). There are also a number of differences in the primary amino acid sequence of the AhR in various species, and it is thought that these differences may be correlated to the susceptibility of a given species to TCDD (Poland et al., 1994; Pohjanvirta et al., 1998; Korkalainen et al., 2000). The AhR contains a basic helix-loop-helix motif and a periodicity/aryl hydrocarbon receptor nuclear translocator/single-minded (Per/ARNT/Sim) homology domain in the N-terminal half of the protein and a complex transactivation domain in the C-terminal half of the protein. The N-terminal portion of the AhR is fairly well conserved across species, whereas the C-terminal half is poorly conserved.
Mouse models are used in toxicologic studies to determine the effects of TCDD, and the data are then extrapolated to estimate risk to humans. Differences between the mouse and human AhRs may result in a variation in response to TCDD exposure. Some of the important differences between the receptors in different species include transactivation potential, target gene specificity, relative affinity for hsp90 and XAP2, and the receptors' relative affinities for different ligands. hAhR is less stable than mAhR in vitro and requires molybdate in the homogenization buffer to maintain the receptor in a conformation that can bind ligand (Manchester et al., 1987). hAhR also seems to have a lower affinity for hsp90 than mAhR and binds about 50% less hsp90 (Hogenesch et al., 1997). Transiently expressed mAhR and hAhR differ in subcellular localization in COS-1 cells (Ramadoss et al., 2004). The hAhR may also bind less XAP2 than mAhR, and XAP2 exhibits differing effects on the two receptors in terms of subcellular localization and its ability to increase receptor levels (Ramadoss et al., 2004).
There are four AhR alleles in inbred strains of mice: Ahb-1, Ahb-2, Ahb-3, and Ahd (Poland and Glover, 1990). Although the three Ahb alleles have a high affinity for ligand, the Ahd allele binds ligand poorly; this has been shown to be the result of a single amino acid change in the ligand binding domain of the receptor (Poland et al., 1994). The mAhRb-1 allele is the most commonly studied receptor of the four mouse alleles. The relative ligand affinities of mAhRb-1 and hAhR have not been studied extensively, but hAhR has been shown to have a lower affinity for [3H]TCDD than mAhRb-1 in vitro (Harper et al., 1988; Ema et al., 1994). Because hAhR apparently has a lower affinity for ligand, it is difficult to study hAhR's ligand binding characteristics in vitro. In this study, we examined the relative ligand affinities1 of mAhRb-1 and hAhR for a photoaffinity ligand, 2-azido-3-[125I] iodo-7,8-dibromodibenzo-p-dioxin, using in vitro assays as well as assays in cultured cells. Initial experiments were carried out with a reversible ligand, 2-[125I]iodo-7,8-dibromodibenzo-p-dioxin. However, the hAhR seemed to have a very low affinity for this reversible ligand; consequently, it was difficult to compare mAhRb-1 and hAhR without performing density gradient analysis. In reversible binding assays, excess free ligand has to be removed using dextran-coated charcoal, which effectively stripped the ligand off the hAhR complex. The photoaffinity ligand is less hydrophobic than the reversible ligand and is not stripped off the receptor complex by charcoal to a great extent. Although assays using the photoaffinity ligand cannot measure absolute KD values, these assays are not complicated by inaccuracies in measurements of free ligand and nonspecific binding. In this report, we show that the mAhRb-1 has a higher relative affinity for the photoaffinity ligand than hAhR. We also show that the difference in relative ligand affinity for the photoaffinity ligand between the two receptors may not be accurately detected in vitro, at least not when using low protein concentrations in the assay. However, differences in the off-rate of ligand binding were easily seen in the presence of high concentrations of a competing ligand or if high protein concentrations were used in the assays. To avoid the limitations of the in vitro assay, we also studied the differences in relative ligand affinity between mAhRb-1 and hAhR directly in cells; again, the mAhRb-1 had a higher relative affinity for the photoaffinity ligand. Studying the level of Ah receptor occupancy by ligand directly in cells is likely to be a superior method to study mutant receptors, polymorphic receptors, or receptors from different species.
Materials and Methods
Construction and Sources of Expression Vectors
pcDNA3/mAhR/FLAG (i.e., mAhRb-1) and pCI/hAhR/FLAG were generated previously (Meyer et al., 1998). pcDNA3/mAhRd was generated by subcloning mAhRd from pALTER-1/mAhRd (kindly provided by Dr. Chris Bradfield) using HindIII and XhoI restriction sites. The receptor mutants pcDNA3/mAhRK13A/FLAG, pCI/hAhRK14A/FLAG and pCI/hAhR/V381A/FLAG were generated by site-directed mutagenesis (QuikChange; Stratagene, La Jolla, CA) using pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG as templates.
Cell Culture
COS-1 cells were grown in α-minimal essential medium (Sigma, St. Louis, MO) supplemented with 10% fetal bovine serum (Hyclone Labs, Logan, UT), 100 IU/ml penicillin, and 0.1 mg/ml streptomycin (Sigma), at 37°C in 95% air/5% CO2.
Photoaffinity Ligand Synthesis
2-Azido-3-[125I]iodo-7,8-dibromodibenzo-p-dioxin was synthesized as described earlier (Poland et al., 1986). The specific activity of the photoaffinity ligand was 2176 Ci/mmol, and it was stored in methanol.
Transient Transfections and Cytosol Preparation
For in vitro ligand-binding experiments, COS-1 cells were transfected in 100-mm dishes with 6 μg of receptor using LipofectAMINE Plus (Invitrogen, Carlsbad, CA) according to the manufacturer's directions. Cells were harvested approximately 20 h after transfection by trypsinizing the cells, collecting the cell pellet, and washing the pellet with PBS. Cells were then resuspended in MENG with 20 mM sodium molybdate and homogenized for 40 strokes in a Dounce homogenizer. The crude lysate was then centrifuged at 100,000g for 45 min, and the supernatant was collected. Total protein concentrations were estimated using the bicinchoninic acid assay kit (Pierce, Rockford, IL). For ligand binding assays in cultured cells, COS-1 cells were transfected in six-well plates with 1.5 μg of receptor per well using LipofectAMINE Plus.
Ligand Binding Assays
Saturation Binding. All binding experiments were carried out in the dark, until UV-mediated activation of photoaffinity ligand. Cytosol from transiently transfected COS-1 cells was diluted to a final protein concentration of 0.5 mg/ml. Increasing amounts of photoaffinity ligand were added to borosilicate glass tubes containing 5 μl of dimethyl sulfoxide (Sigma) and counted in a γ-counter. Each point on the curve was determined by duplicate experimentes. Cytosol (150 μl at 0.5 mg/ml) was added to the tubes containing ligand and incubated for 30 min at room temperature (∼25°C). The samples were placed on ice for 5 min; dextran-coated charcoal was added to the samples to a final concentration of 1%/0.1% and incubated on ice for 20 min. The cytosol was then centrifuged at 3000g for 10 min at 4°C. The samples were photolysed at >302 nm, at a distance of 8 cm, for 4 min using two 15-W UV lamps (Dazor Mfg. Corp. St. Louis, MO). The supernatant was transferred into a microcentrifuge tube containing SDS sample buffer, heated for 5 min at 95°C, and followed by TSDS-PAGE (8% acrylamide). Gels were dried and exposed to film, and bands were subsequently excised and counted in a γ-counter (Amersham Biosciences, Piscataway, NJ).
Competition Experiment. A single saturating concentration of photoaffinity ligand (0.13 pmol; i.e., 5 × 105 cpm per tube) was used, and each point was determined by duplicate experiments. Competition experiments were carried out in a similar way to the saturation binding experiments, except that after 30 min of cytosol incubation with ligand at room temperature, a 200-fold molar excess of nonradioactive 2,3,7,8-tetrachlorodibenzofuran (TCDBF) was added to the tubes and incubated for the specified time. The samples were then placed on ice, charcoal was added, and the same procedure as that in the saturation binding experiment was followed.
High Protein Concentration Experiments. Cytosol from transiently transfected COS-1 cells was diluted to 6 different concentrations, such that the amount of cytosolic protein in each sample on the curve was 75 μg but the final volume of the sample remained 150 μl. The total amount of protein in the reaction tubes ranged between 0.075 and 1.5 mg. BSA was added (in solution) to bring the protein concentration to the required amount and the volume to 150 μl. A single saturating concentration of photoaffinity ligand (0.13 pmol; i.e., 5 × 105 cpm per tube) was added to each tube and incubated with the protein solution. Each point was done in duplicate. Incubation and subsequent steps were carried out the same way as with saturation binding experiments. An aliquot of the photolysed sample was resolved by TSDS-PAGE.
Ligand Binding Experiments in Cell Culture (PL-Cells Assays). These experiments were done essentially as described previously (Swanson and Perdew, 1991). Media from the transfected cells in six-well plates was removed, and fresh media containing the appropriate amount of photoaffinity ligand was added to each well. Each concentration of ligand was done in duplicate. The plates were then incubated at 37°C in 95% air/5% CO2 for 1 h. Media was removed, cells were washed once with PBS, and then the plates were exposed to UV light using two 15-W UV lamps at a distance of 8 cm for 4 min. Cells were then lysed on the plate, in MENG buffer containing 1% Nonidet P-40. The lysates were transferred to microcentrifuge tubes, and centrifuged at 14,000 rpm for 10 min at 4°C. Supernatants were removed, mixed with loading buffer and heated, and proteins were separated by TSDS-PAGE. Gels were dried and exposed to film. Band intensities were quantitated by filmless autoradiographic analysis using a Cyclone storage phosphor screen instrument (PerkinElmer Life and Analytical Sciences). Band intensities estimated by γ-counting or filmless autoradiographic analysis gave similar results, and filmless autoradiographic analysis was performed for PL-Cells assays because it was easier to estimate low-intensity bands in these experiments using this method.
Western Blot for Protein Expression Levels
Cytosolic proteins from transfected COS-1 cells were separated by TSDS-PAGE and transferred to polyvinylidene difluoride membrane (Millipore, Bedford, MA). The AhR was detected using the monoclonal antibody RPT1 (Perdew et al., 1995) and an iodinated goat-antimouse secondary antibody (Amersham Biosciences). For the PL-Cells assay, a triplicate set of transfections was set up using the same transfection mix as the ligand binding plates. The cells were lysed directly on the plate using MENG containing 1% Nonidet P-40, and lysates were collected, centrifuged, and supernatants resolved by TSDS-PAGE. Proteins were transferred to polyvinylidene difluoride membrane and Western blotting was performed as described above.
Results
mAhRb-1 and hAhR Have Similar Saturation Binding Curves. The most commonly studied mouse AhR allele is mAhRb-1, and we used this allele to compare the mouse and human Ah receptors. COS-1 cells were transfected with either pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG. Cytosol was extracted, and ligand-binding assays were performed using increasing concentrations of ligand and a fixed amount of cytosolic protein. The samples were treated with dextran-coated charcoal, centrifuged, photolysed, and resolved by TSDS-PAGE (Fig. 1A.). Untransfected COS-1 cell lysate was used in control ligand binding assays. Radioactive AhR bands were excised from the gel and counted in a γ-counter. A plot of the radioactive counts in the AhR band versus the input radioactivity for each point on the curve (average values from each duplicate) was made. From the saturation binding curve, mAhRb-1 and hAhR seem similar in their ability to bind ligand (Fig. 1B) The expression levels of the two receptors can be seen to be similar in the western blot of the cytosol from transfected COS-1 cells (Fig. 1C). Because these results were unexpected, we hypothesized that the result may have been influenced by the low concentration of cytosolic protein in the assay. Under these conditions, the hydrophobic ligand may be “forced back” on to the Ah receptor after dissociation. mAhRb-1 Has a Greater Affinity for Ligand Than hAhR in Competition Binding Experiments. To test whether the dissociation rates of ligand from the ligand-receptor complex differed between mAhRb-1 and hAhR, we performed a competition experiment with a single saturating concentration of photoaffinity ligand (0.13 pmol/tube) and a 200-fold molar excess of 2,3,7,8-tetrachlorodibenzofuran, a high-affinity ligand for the AhR. Cytosol from COS-1 cells transfected with pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG was incubated with 0.13 pmol of photoaffinity ligand and subsequently incubated with TCDBF for the indicated time. Samples were treated with dextran-coated charcoal, centrifuged, photolysed, and resolved by TSDS-PAGE (Fig. 2A). Radioactive bands were excised, and a plot of the radioactive counts in the AhR band versus the incubation time with TCDBF was made (Fig. 2B). The expression levels of mAhR and hAhR are similar, because the cytosol used for this experiment was the same cytosol used in the saturation binding experiment (Fig. 1C). The mAhRb-1 seems to have a lower rate of ligand dissociation compared with hAhR, in that the rate at which the photoaffinity ligand displaced by TCDBF is much higher in the case of hAhR. Therefore, the apparent similarity in binding curves of mAhRb-1 and hAhR may be an artifact of the (in vitro) experimental conditions used. Thus, the saturation binding curves under the experimental conditions used in Fig. 1 may not be representative of the binding affinity of the AhR.

Saturation binding analysis of mAhR compared with hAhR in cell lysates. Approximately 75 μg of cytosol from untransfected COS-1 cells or COS-1 cells transfected with pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG was incubated with increasing amounts of photoaffinity ligand for 30 min. Excess free ligand was removed using dextran-coated charcoal. Receptor-ligand complexes were photolysed and resolved by TSDS-PAGE. The gels were dried and exposed to film; thereafter, bands were excised from the gel and counted in a γ-counter. A, autoradiograph of gels. B, graphic analysis of data from A. C, determination of AhR levels using protein blot analysis of cytosol used in binding experiments. The data shown are representative of three experiments; each data point in an experiment is an average of a duplicate set.

Comparison of mAhR and hAhR in cell lysates by competitive displacement of the photoaffinity ligand. Approximately 75 μg of cytosol from COS-1 cells transfected with pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG was incubated with 0.13 pmol of photoaffinity ligand for 30 min, followed by the addition of a 200-fold molar excess of TCDBF for the indicated time. Excess free ligand was removed using dextran-coated charcoal, and receptor-ligand complexes were photolysed and resolved by TSDS-PAGE. A, autoradiograph of gels containing the competing ligand displacement samples. B, plot of AhR band intensity versus time of incubation with competitor ligand. The data shown are representative of two experiments; each data point in an experiment is an average of a duplicate set.
Increasing Protein Concentrations in Ligand Binding Assays Demonstrates that hAhR Has a Lower Affinity for Ligand. Because ligand binding assays are carried out in an aqueous cytosol solution, we hypothesized that the ligand may bind to the AhR more readily because the environment is less favorable for it to dissociate from the receptor and exist free in solution or bind to low-affinity protein binding sites. This may be the reason why the mAhRb-1 and hAhR seem to have similar saturation binding curves, as shown in Fig. 1. It was demonstrated previously that increasing the protein concentration of the cytosol solution had an effect on the KD using the reversible ligand 2-[125I]iodo-7,8-dibromodibenzo-p-dioxin (Bradfield et al., 1988). We wanted to test whether increasing the protein concentration of the cytosol solution would also affect binding of the photoaffinity ligand to the mAhR and hAhR. The amount of cytosolic protein was maintained at 0.075 mg per tube, whereas the total amount of protein was varied from 0.075 to 1.5 mg per tube by adding BSA. A single saturating concentration of photoaffinity ligand (0.13 pmol/tube) was incubated with the cytosol for 30 min, and excess ligand was removed by dextran-coated charcoal treatment. Samples were centrifuged, photolysed, and resolved by TSDS-PAGE. The gels were then dried and exposed to film (Fig. 3A). AhR band intensities were determined by filmless autoradiographic analysis, and a plot of AhR band intensity versus total protein in the assay tube was made (Fig. 3B). An increase in protein concentration resulted in a decrease in the level of ligand binding to the AhR, suggesting that as the protein concentration in the assay tube increases, the ligand is not forced to remain bound to the AhR and could be free in solution or bound by other proteins. The difference in relative ligand affinity between mAhRb-1 and hAhR can be detected in the presence of high concentrations of protein. Increasing protein concentration in the assay tube has a more profound effect on ligand binding to the hAhR than to mAhRb-1, suggesting that hAhR has a lower affinity for the ligand than mAhRb-1.

Influence of protein concentration on photoaffinity ligand binding in cell lysates. Ligand binding assays were carried out with approximately 75 μg of cytosolic protein in each assay tube, whereas the total amount of protein ranged from 0.075 to 1.5 mg by the addition of BSA, with 0.13 pmol of photoaffinity ligand for 30 min. Excess free ligand was removed, and complexes were photolysed and resolved by TSDS-PAGE. Band intensities were estimated by filmless autoradiographic analysis. A, autoradiograph of gels containing photolysed samples with varying protein concentrations. B, plot of AhR band intensity in each lane. C, AhR visualized on a protein blot containing an aliquot of each sample used in A. The data shown are representative of three experiments; each data point in an experiment is an average of a duplicate set.
mAhRb-1 Has a Greater Affinity for Ligand Than hAhR in COS-1 Cells. Because hAhR is known to be less stable than mAhRb-1 in vitro (Manchester et al., 1987) and the amount of protein used in the in vitro assay affects the results obtained in this study, we decided to test whether the two receptors have differing ligand affinities directly in cells, using PL-Cells Assay. COS-1 cells were transfected with pcDNA3/mAhR/FLAG or pCI/hAhR/FLAG in six-well plates. Photoaffinity ligand was added directly to the media and incubated for 1 h. The medium containing photoaffinity ligand was removed, cells were washed with PBS, and the plates were exposed to UV light, leading to the covalent attachment of the ligand to the AhR. The cells were then lysed, cytosol was extracted, and protein complexes were resolved by TSDS-PAGE (Fig. 4A). The level of receptor occupancy by ligand is lower for the hAhR, compared with mAhRb-1, in COS-1 cells. mAhRb-1 and hAhR localize differently in COS-1 cells (Ramadoss et al., 2004); mAhR is predominantly nuclear, whereas hAhR is predominantly cytoplasmic. To eliminate the possibility that this could contribute to differences in ligand affinity, we carried out a PL-Cells assay with mouse and human Ah receptors that had mutations in the nuclear localization signals (mAhRK13A-FLAG and hAhRK14A-FLAG). These mutant receptors are cytoplasmic when expressed in COS-1 cells (Ramadoss et al., 2004). In this experiment, mAhRK13A-FLAG seemed to have a higher affinity for ligand than hAhRK14A-FLAG (Fig. 4C), indicating that difference in localization of the receptors in COS-1 cells does not affect ligand binding. Western blot analysis for each of these experiments showed that mAhRb-1 and hAhR, as well as the NLS-mutant receptors, are all expressed at similar levels, and differences in protein levels of the receptors are not the reason for the differences observed in ligand binding (Fig. 4, B and D).

Use of photoaffinity ligand to determine the relative level of ligand binding to mAhR and hAhR in cell culture COS-1 cells that had been transiently transfected with pcDNA3/mAhR/FLAG, pCI/hAhR/FLAG, pcDNA3/mAhRK13A/FLAG, or pCI/hAhRK14A/FLAG were treated directly with the indicated amount of ligand for 1 h. Cells were washed with PBS and photolysed directly on the plate. Cells were lysed, and lysates were resolved by TSDS-PAGE. Band intensities were estimated by filmless autoradiographic analysis imaging. A, autoradiograph of gels, visualization of radiolabeled mAhR-FLAG and hAhR-FLAG (exposure time 6 h). B, Western blot of lysates, immunochemical visualization of mAhR-FLAG and hAhR-FLAG. C, autoradiograph of gels, visualization of radiolabeled mAhRK13A-FLAG and hAhRK14A-FLAG (exposure time 16 h). D, Western blot of lysates, immunochemical visualization of mAhRK13A-FLAG and hAhRK14A-FLAG. The data shown are representative of three experiments; each data point in an experiment is an average of a duplicate set.
hAhR and mAhRd Have Comparable Ligand Affinities. It has been shown previously that the mAhRd allele has a lower affinity for ligand than the mAhRb-1 allele (Poland and Glover, 1990). We wanted to test whether hAhRs affinity for ligand was similar to mAhRd in COS-1 cells. A PL-Cells assay was performed to compare the relative ligand affinities of mAhRb-1, hAhR, and mAhRd using three different ligand concentrations (0.522, 1.04, and 1.56 pmol, respectively). hAhR and mAhRd both appear to have a lower relative affinity for ligand than mAhRb-1 (Fig. 5A). hAhR still has a lower relative affinity for ligand than mAhRd, but hAhR and mAhRd are more comparable than hAhR and mAhRb-1. The difference in the level of receptor occupancy between mAhRb-1 and hAhR is approximately 10-fold at the highest concentration of ligand tested, whereas the difference between mAhRd and hAhR is only 2-fold at this ligand concentration. All three receptors were expressed at similar levels, as seen in the Western blot (Fig. 5B).
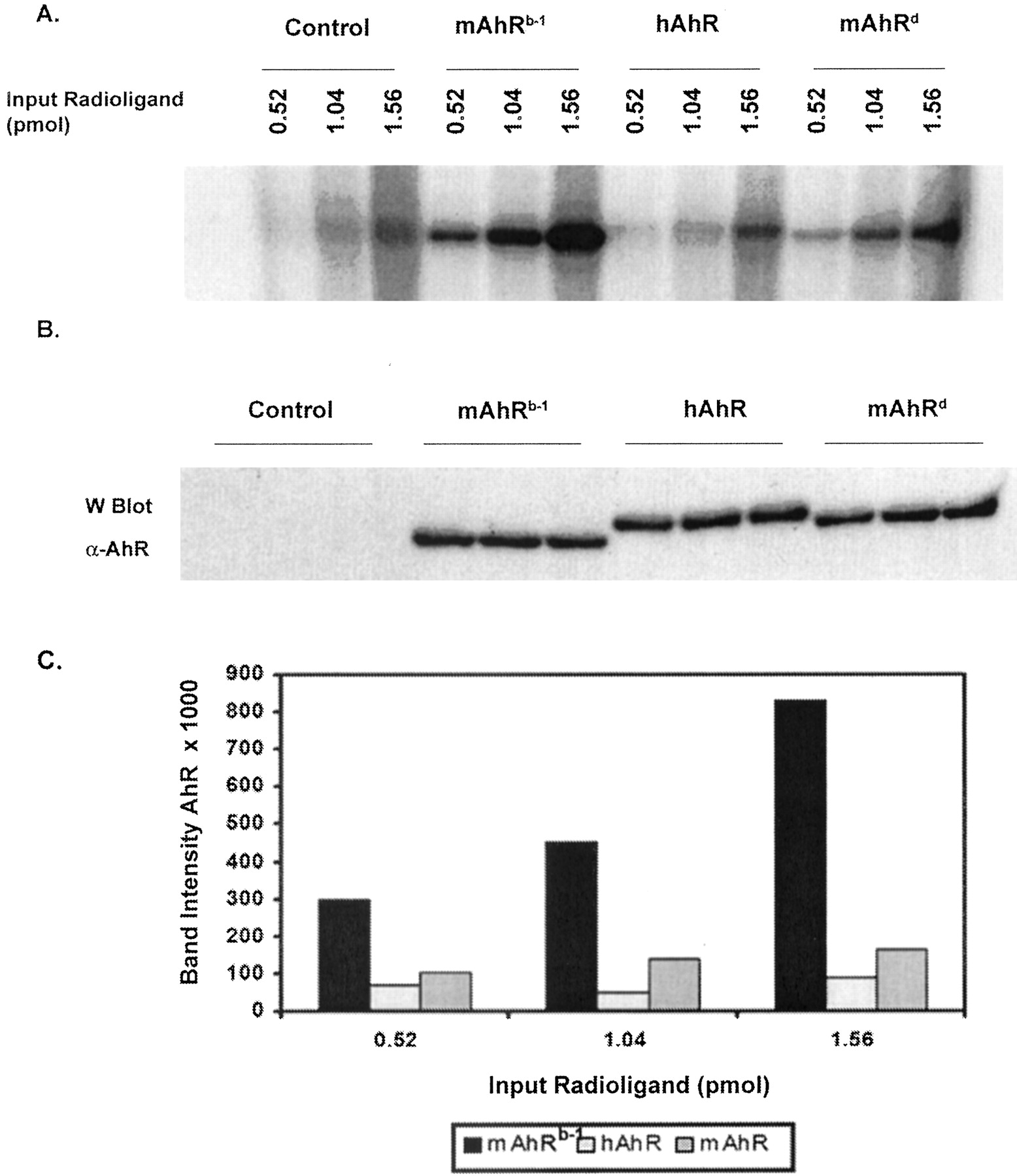
mAhRd and hAhR have similar ligand affinity in COS-1 cells. COS-1 cells were transiently transfected with pcDNA3/mAhR/FLAG (i.e., mAhRb-1), pCI/hAhR/FLAG or pcDNA3/mAhRd and a PL-Cells assay was performed. A, autoradiograph of cell lysates resolved by TSDS-PAGE. B, immunochemical visualization of AhR in cell lysates. C, plot of band intensities in A, estimated by filmless autoradiographic analysis. For each data point, the corresponding background band in the control lane (untransfected COS-1 cells) was subtracted. Each data point is an average of a duplicate set.
A Single Amino Acid Change in hAhR's Ligand Binding Domain (V381A) Restores Ligand Affinity to a Level Similar to That of mAhRb-1. It has been shown that a single amino acid change in mAhRds ligand binding domain is responsible for reduced affinity for ligand compared with mAhRb (Poland et al., 1994). The amino acid at position 375 in mAhRb is alanine, whereas in mAhRd it is valine. It is noteworthy that in hAhR, amino acid 381 (corresponding to amino acid 375 in mAhR) is also valine. However, in a previous report, changing valine 381 to alanine in hAhR restored ligand affinity in hAhR (Ema et al., 1994) only partially. We performed a PL-Cells assay to compare mAhRb-1, hAhR, and hAhR/V381A using three different concentrations of ligand (0.522, 1.04, and 1.56 pmol, respectively). hAhRV381A seems to have a relative ligand affinity similar to mAhRb-1, indicating that a single amino acid difference (valine 381) is responsible for a lower relative ligand affinity observed with the human receptor (Fig. 6A).
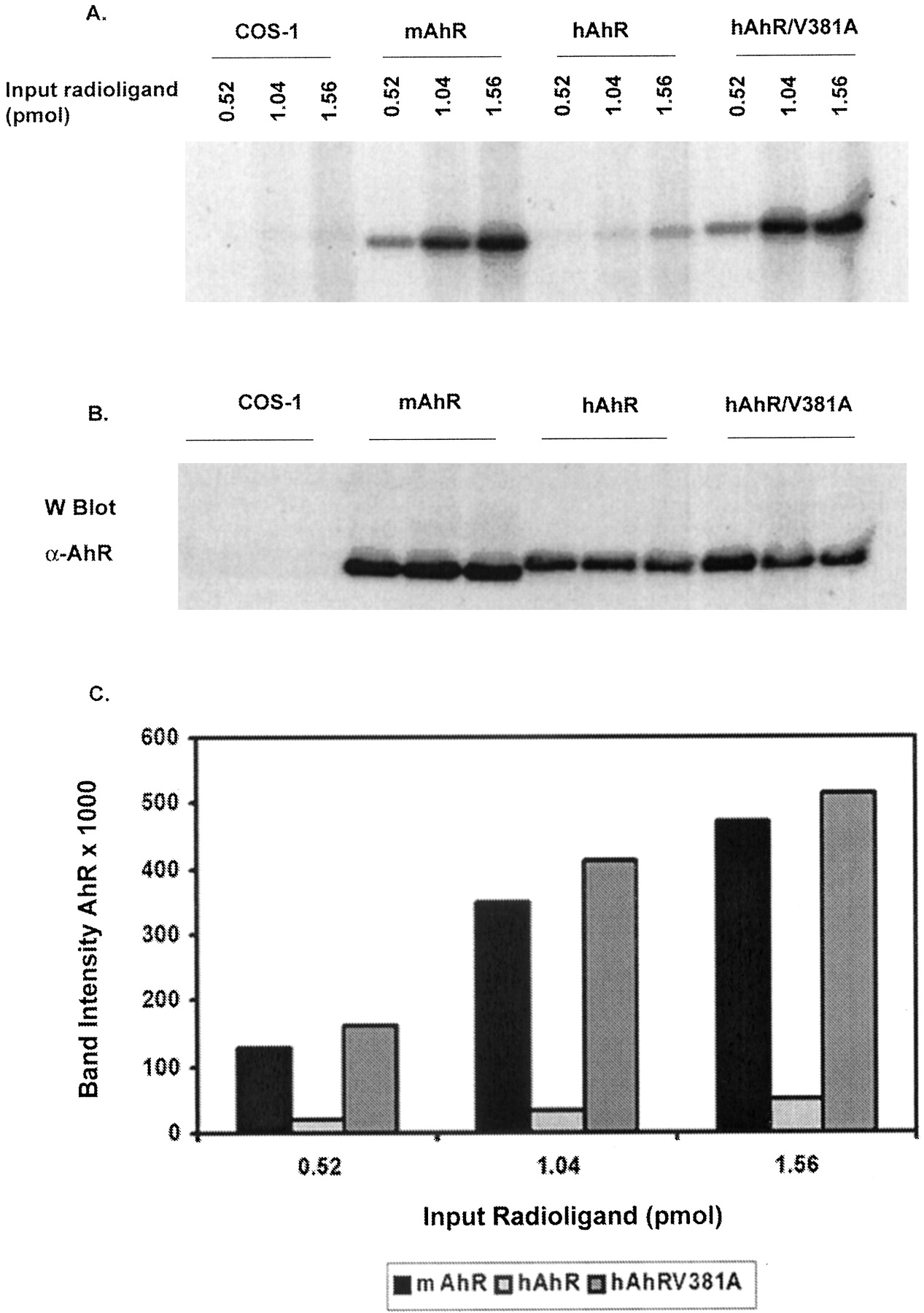
mAhRb-1 and hAhR/V381A have a similar ligand affinity in COS-1 cells. COS-1 cells were transiently transfected with pcDNA3/mAhRb-1/FLAG, pCI/hAhR/FLAG, or pCI/hAhR/V381A/FLAG and a PL-Cells assay was done. A, autoradiograph of cell lysates resolved by TSDS-PAGE. B, immunochemical visualization of AhR in cell lysates. C, plot of band intensities in A, estimated by filmless autoradiographic analysis. For each data point, the corresponding background band in the control lane (untransfected COS-1 cells) was subtracted. Each data point is an average of a duplicate set.
Discussion
The differences between species in terms of their sensitivity to TCDD may be caused by variation in the Ah receptor. A number of different properties of the AhR could vary between species, one of which is the AhR's relative ligand affinity. In mice, it has been shown that the mAhRd has a lower ligand affinity compared with that of mAhRb (Poland and Glover, 1990). In addition, mouse strains carrying the Ahd allele are more resistant to treatment with polycyclic aromatic hydrocarbons compared with the strains carrying the Ahb-1 allele (Chapman and Schiller, 1985). The differences in relative ligand affinity between mAhRb-1 and hAhR have not been characterized extensively, but it has been shown that the human receptor appears to have a lower relative affinity for ligand than the mouse receptor (Harper et al., 1988; Ema et al., 1994). These assays were done in vitro using glycerol density gradients. Dextran-coated charcoal was used to remove excess free ligand, which could potentially strip ligand off the receptor-ligand complex, particularly in the case of the human receptor. In addition, ligand could dissociate from the AhR during the long centrifugation and fractionation on a linear glycerol gradient. In this study, we wanted to compare the relative ligand affinities of mAhRb-1 and hAhR for a photoaffinity ligand, which may allow the detection of the relative level of binding at or near steady state levels, in contrast to the density gradient method.
Saturation binding experiments showed very little difference in relative ligand affinity between mAhRb-1 and hAhR (Fig. 1). This result was surprising, because it has been shown previously that hAhR appears to have a lower relative affinity for ligand compared with mAhR (Harper et al., 1988; Ema et al., 1994). However, this result does prove that the photoaffinity ligand can be covalently bound to each receptor to an equal extent, upon exposure to UV light. Therefore, the differences in relative ligand affinity seen in subsequent assays are not caused by a difference in the efficiency of covalent attachment of ligand to each receptor. When competitive displacement experiments were performed, mAhRb-1 seemed to have a higher affinity for the ligand than hAhR, because the photoaffinity ligand was more readily displaced by a competing ligand in the case of hAhR (Fig. 2). After 40 min of incubation with excess TCDBF, approximately 70 and 12% of the initial amount of photoaffinity ligand remained bound to mAhR and hAhR, respectively.
We then hypothesized that because the ligand is hydrophobic, it is forced to bind to the receptor in a more polar environment, such as in cytosol, in which saturation binding experiments were carried out. Even if the ligand dissociates from the receptor complex, it may re-associate with the complex because of the unfavorable environment. This may have been the reason we were initially unable to detect a difference between mAhRb-1 and hAhR. To determine whether this was the case, we increased the protein concentration in the in vitro assays, which would increase the hydrophobicity or increase the low-affinity nonspecific binding sites. It has been shown that lower protein concentrations in ligand binding assays lower the KD for 2-[125I]iodo-7,8-dibromodibenzo-p-dioxin (Bradfield et al., 1988). In the current study, the AhR bound to the ligand less efficiently in the presence of high concentrations of protein (Fig. 3), which is consistent with results published previously (Bradfield et al., 1988). The hAhR bound relatively less ligand than the mAhRb-1 in the presence of high concentrations of protein. Therefore, the hAhR has a lower affinity for the photoaffinity ligand than mAhRb-1, but the results obtained in in vitro assays are dependent on the concentration of cytosolic protein used in the assay.
To circumvent the potential problems with interpretation of the results from in vitro assays, we decided to examine ligand binding within cells. This approach could test the level of occupancy of the receptor within cells at low concentrations of ligand and may therefore yield more physiologically relevant data. Because the cellular environment is very different from cytosolic preparations used in in vitro assays (e.g., in terms of protein concentration, temperature, and redox potential), we considered it likely that differences in relative ligand affinity between mAhR and hAhR would be detected in cells. The hAhR binds approximately 10-fold less ligand than mAhRb-1 in cells (Fig. 4), which is consistent with the observation that under conditions of high protein concentration, hAhR binds less ligand. A comparison of mAhRb-1, hAhR, and mAhRd showed that hAhR and mAhRd have lower ligand affinities than mAhRb-1 (Fig. 5). Because mAhRd seems to have a relative ligand affinity for the photoaffinity ligand that is more comparable with hAhR, their affinities for other ligands may also be similar. If this is the case, strains of mice carrying the Ahd allele may serve as a better model for toxicological studies that aim to estimate risk to humans.
Amino acid 375 is alanine in mAhRb-1, whereas it is valine in mAhRd. In hAhR, amino acid 381 (homologous to amino acid 375 in mAhR) is also valine. It has been shown that in mAhRd, when valine 375 is mutated to alanine, the ligand affinity of mAhRd is restored to levels similar to mAhRb (Poland et al., 1994). It has also been shown that when valine 381 in hAhR is mutated to alanine, there is a partial recovery of ligand affinity, in the case of hAhR, and that a deletion of 40 amino acids at the C-terminal end of the receptor in addition to the V381A mutation was required to further enhance ligand affinity (Ema et al., 1994). Because these assays were done in vitro, we wanted to examine whether the human V381A mutant receptor would behave the same way in COS-1 cells as in the in vitro experiment. In this study, we found that hAhR/V381A seems to be able to bind ligand as efficiently as mAhRb-1 (Fig. 6). It is possible that the results from the previous study (Ema et al., 1994) differ from the results in this study because of the limitations of in vitro experiments. This result also indicates that the divergence between the mAhR and hAhR in the C-terminal half of the receptor does not influence the ligand-binding potential.
The biological response to exposure to TCDD or other ligands in a given species is determined by physiological factors, as well by the properties and behavior of the Ah receptor at the cellular/molecular level. The fact that mAhRb-1 has a higher affinity for some ligands compared with hAhR may result in differences in response to ligand exposure. Other properties of the receptor, such as transactivation potential, specificity of target genes, and interaction with other cellular proteins, are also likely to influence the toxicological endpoint induced upon exposure to different ligands. The difference in ligand affinities is also a factor that needs to be kept in mind when assessing differences in other properties of the receptors. For example, a difference in the extent to which mAhR or hAhR can activate gene expression may be a combined effect of ligand affinity and transactivation potential. To compare the transactivation potential of mAhR and hAhR, without the bias of differences in ligand affinities, mAhRb-1 and hAhRV381A could be compared.
It is likely that the in vitro assay may not reflect actual differences between different receptors because the experiment is dependent on the protein concentration in the assay tube. One way to overcome this limitation is to increase the protein concentration in in vitro assays, but the optimal protein concentration at which to carry out the experiment has to be determined. The differences in in vitro stability between receptors could also influence the results of ligand binding experiments. The PL-Cells assay, using the photoaffinity ligand, is a more reliable way to study differences in relative ligand affinity between different Ah receptors. The experiment is not limited by possible differences in in vitro lability of different receptors, and because the protein concentrations within cells represent a more physiologically relevant condition, differences in ligand affinity between receptors as seen in this experiment are likely to be significant. The PL-Cells assay is less laborious than the in vitro experiment done using density gradients. It is also a suitable system to study receptors that have a low affinity for ligand, such as hAhR, which cannot be studied as effectively using the reversible ligand 2-[125I]iodo-7,8-dibromodibenzo-p-dioxin. This report demonstrates that there is a difference between the hAhR and mAhRb-1 in terms of their relative affinities for a photoaffinity ligand, but this may only be representative of a group of similar ligands. The relative affinities of mAhRb-1 and hAhR for other ligands may be different.
In summary, there is a difference in relative ligand affinity between mAhRb-1 and hAhR for the photoaffinity ligand, which is not seen in saturation binding experiments carried out using cytosol with low protein concentrations. Competition binding experiments demonstrated a difference in ligand affinity between the two receptors. Increasing the protein concentration in the assay demonstrated that ligand binds less efficiently to the AhR, suggesting that the results obtained in the in vitro assay are influenced and limited by the fact that these experiments are performed using relatively low protein concentrations. PL-Cells assays readily demonstrate the difference between mAhRb-1 and hAhR within the context of the cell and therefore may be a better system to use to firmly establish differences between receptors in terms of relative ligand affinity. In addition, the PL-Cells assay also allows the study of the level of ligand binding to be assessed under physiologically relevant conditions that more closely mimic the context of actual exposure to dioxin and other AhR ligands.
Acknowledgments
We thank Dr. Chris Bradfield for providing pALTER-mAhRd (PL2047) and Dr. Steve Safe for 2,3,7,8-tetrachlorodibenzofuran.
Footnotes
-
↵1 Throughout this article, we use the term “relative ligand affinity”, which we operationally define to mean the relative level of ligand occupancy of the AhR at a given concentration of the photoaffinity ligand at the time of photolysis.
-
This study was funded by National Institute of Environmental Health Sciences grants ES04869 and ES011834.
-
ABBREVIATIONS: TCDD, 2,3,7,8-tetrachlorodibenzo-p-dioxin; Ah, aryl hydrocarbon; AhR, aryl hydrocarbon receptor; XAP, X-associated protein; hsp, heat shock protein; hAhR, human aryl hydrocarbon receptor; MENG, 25 mM MOPS/2 mM EDTA/0.02% NaN3/10% glycerol; MOPS, 3-(N-morpholino)propanesulfonic acid; TSDS, Tricine SDS; PAGE, polyacrylamide gel electrophoresis; TCDBF, 2,3,7,8 tetrachlorodibenzofuran; BSA, bovine serum albumin; PBS, phosphate-buffered saline; PL-Cells assay, photoaffinity labeling of aryl hydrocarbon receptor in cultured cells.
- Received February 6, 2004.
- Accepted April 12, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}