


Abstract
The plasma membrane distribution and related biological activity of nucleoside transporter proteins (NTs) were investigated in human syncytiotrophoblast from term placenta using a variety of approaches, including nucleoside uptake measurements into vesicles from selected plasma membrane domains, NT immunohistochemistry, and subcellular localization (basal, heavy, and light apical membranes as well as raft-enriched membranes from the apical domain). In contrast with other epithelia, in this epithelium, we have identified the high-affinity pyrimidine-preferring human concentrative nucleoside transporter (hCNT) 1 as the only hCNT-type protein expressed at both the basal and apical membranes. hCNT1 localization in lipid rafts is also dependent on its subcellular localization in the apical plasma membrane, suggesting a complex cellular and regional expression. Overall, this result favors the view that the placenta is a pyrimidine-preferring nucleoside sink from both maternal and fetal sides, and hCNT1 plays a major role in promoting pyrimidine salvage and placental growth. This finding may be of pharmacological relevance, because hCNT1 is known to interact with anticancer nucleoside-derived drugs and other molecules, such as nicotine and caffeine, for which a great variety of harmful effects on placental and fetal development, including intrauterine growth retardation, have been reported.
Introduction
Fetal exposure to xenobiotics is known to be harmful in many cases, compromising gestation itself and neonatal development. The placenta is known to express a broad scope of xenobiotic transporters (Leazer and Klaassen, 2003), which appear to be key players in the regulation of placental vectorial flux of nutrients and xenobiotics, potentiating either their maternofetal transfer or their detoxification. Among the plasma membrane proteins that might play this dual role of being both nutrient and xenobiotic transporters, most members of the SLC28 and SLC29 gene families have been reported to be expressed in the placenta at the mRNA level (Barros et al., 1995; Griffiths et al., 1997; Gray et al., 2004; Govindarajan et al., 2007; Errasti-Murugarren et al., 2009). SLC28 and SLC29 genes encode human concentrative nucleoside transporter (hCNT) and human equilibrative nucleoside transporter (hENT) proteins, respectively (Pastor-Anglada et al., 2008; Young et al., 2008). To date, only human equilibrative nucleoside transporter proteins hENT1 and hENT2 have been reported to be expressed in placenta, although their plasma membrane localization is still unclear (Barros et al., 1995; Govindarajan et al., 2007). Occurrence of hCNT proteins in human placenta has not been studied in detail, although the human syncytiotrophoblast-derived cell line BeWo has been shown to express hCNT3 mRNA and protein (Yamamoto et al., 2007).
The elucidation of what particular types of CNT proteins are expressed in the human syncytiotrophoblast, along with the analysis of their polarized expression relative to that of ENT-type proteins, may be of great interest in pharmacology. This assumption is based on the evidence that NT proteins mediate the translocation of natural nucleosides and also the uptake of most nucleoside-derived anticancer drugs and some nucleoside-derived inhibitors of retroviral reverse transcriptases used in antiviral therapies (Errasti-Murugarren and Pastor-Anglada, 2010). Some reports have related nucleoside-derived therapies with placental dysfunction. The demethylating agent 5-azacytidine, a good hCNT1 and hENT1 substrate (Huang et al., 2004; Rius et al., 2009), has been reported to induce a variety of harmful effects in rodent placenta, including reduced mass, altered structure, and impaired proliferation of trophoblast cells (Serman et al., 2007). Gemcitabine, a suitable hCNT1, hCNT3, hENT1, and hENT2 substrate (Mackey et al., 1999; Errasti-Murugarren et al., 2007), has been reported to induce developmental toxicity in mice (Eudaly et al., 1993), whereas it has been shown recently that placental tissue of HIV-1 infected antiretroviral therapy-exposed pregnancies shows mitochondrial DNA depletion (Gingelmaier et al., 2009). Moreover, HIV-uninfected children born to HIV-infected mothers are apparently at risk of long-term mitochondrial toxicity (Poirier et al., 2003).
Of interest, nucleoside transporters are also known to interact with compounds such as caffeine and nicotine with high affinity (Lang et al., 2004). Both xenobiotics have been comprehensively studied as risk factors during pregnancy. Among many other observations in the literature, it is known that caffeine intake increases the risk of early spontaneous abortion among nonsmoking women (Cnattingius et al., 2000), whereas modest maternal caffeine exposure may impair fetal cardiovascular function and growth (Momoi et al., 2008) and also promotes long-lasting behavioral changes in mouse offspring (Björklund et al., 2008). Nicotine exposure during pregnancy has also been related to a variety of harmful effects, including, among many others, intrauterine growth retardation (Einarson and Riordan, 2009).
On the basis of this previous evidence, the goal of this contribution was to investigate the expression, activity, and subcellular localization of nucleoside transporters in nonpathological syncytiotrophoblasts. This investigation is of crucial interest for the understanding of the role these transporters play in placental physiology, but, more importantly, this study also represents the first step in the future analysis of how drug-induced placental dysfunction can be associated with altered NT-related transport processes, thereby affecting fetal growth and performance.
Materials and Methods
Reagents.
Uridine, cytidine, adenosine, thymidine, guanosine, 5′-deoxy-5-fluorouridine, and zidovudine (AZT) were obtained from Sigma-Aldrich (St. Louis, MO). Tritiated uridine ([5,6-3H], 35–50 Ci/mmol), thymidine ([5,6-3H], 35–50 Ci/mmol), and adenosine ([5,6-3H], 35–50 Ci/mmol) were purchased from GE Healthcare (Chalfont St. Giles, Buckinghamshire, UK). Cytidine ([5-3H(N)], 21.5 Ci/mmol) and guanosine ([8-3H(N)], 7 Ci/mmol) were purchased from Moravek Biochemicals (Brea, CA).
RNA Isolation and RT-PCR.
Total RNA was extracted from placenta lysates using an RNeasy Mini Kit (QIAGEN, Barcelona, Spain). RNA was treated with DNase I from an RNase-Free DNase Set (QIAGEN) to eliminate contaminating DNA. cDNA was generated from 1 μg of total RNA by reverse transcription using TaqMan reagents as described by the manufacturer (Applied Biosystems, Foster City, CA). Different sets of primers were designed and synthesized for PCR analysis of each transporter. For amplifying hCNT1, the primers used were 5′-GAG GGG TCT AGC TCT TGC TG-3′ (forward) and 5′-CAC CTT CAC GGA GAT GGC GGC C-3′ (reverse), which generated a 822-bp product; for hCNT2, the primers were 5′-CCC GCC TGA GGC TTT GGA CG-3′ and 5′-CAA CCC CAA AGG CTA TGA AGG-3′, which generated a 650-bp product; for hCNT3, the primers were 5′-GCA CAC TCA AAC TGC TCC ACC-3′ and 5′-GGG CTC TGT GAA AGT TCA GC-3′, which generated a 446-bp product and a 622-bp insert-containing (hCNT3ins) product; for hENT1, the primers were 5′-GGG CCA CCG CCT GCT GAA ACG G-3′ and 5′-CCT TGA CCA GCG CCT CCC C-3′, which generated a 410-bp product; and for hENT2, the primers were 5′-CCT CAA CTC CTT CCT GTA CC-3′ and 5′-CCC ACA CAG GGC GTG ATA AAG-3′, which generated a 355-bp product. PCRs were performed using the following cycling conditions: 40 cycles of 94°C for 1 min; 55°C (hCNT1, hCNT2, hENT1, and hENT2) or 57°C (hCNT3) for 1 min and 72°C for 3 min, followed by a final extension at 72°C for 15 min and cooling to 4°C. The amplified fragments were separated and visualized on 1% ethidium bromide-stained agarose gels.
Preparation of Syncytiotrophoblast Plasma Membranes.
Placentae were obtained from normal pregnancies and collected at term immediately after delivery from the San José Hospital Maternity Unit (Santiago, Chile). Human placental plasma membrane vesicles were prepared from fresh normal-term placentae by a method described previously that allows the simultaneous isolation of apical and basal membranes from the same placenta (Jimenez et al., 2004). This method involves differential centrifugation, precipitation of nonmicrovillous membranes with magnesium ions, and a sucrose gradient step. All solutions were buffered with 20 mM Tris-maleate, pH 7.4. A portion (0.5–1 ml) of either the microvillous or basal membrane-enriched fraction was overlaid on the surface of a discontinuous sucrose gradient. The bands at the 10%/37% and 37%/45% sucrose interfaces correspond to the light microvillous membrane (LMVM) fraction and heavy microvillous membrane (MVM) fraction, respectively, whereas the band at the 47%/52% interface corresponds to the basal membrane (BM) fraction. Each fraction was collected and diluted 10-fold with 20 mM Tris-maleate, pH 7.4, and centrifuged at 110,000g for 30 min. The final pellets were resuspended in preload medium containing 250 mM sucrose, 0.2 mM CaCl2, 20 μM MgCl2, 10 mM HEPES, pH 7.5, and 150 mM KSCN and stored in liquid nitrogen. The protein concentration was determined using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA). The purity and enrichment of the apical and basal membrane fraction were determined routinely by assaying for classic marker protein activities as described previously (Jimenez et al., 2004). Alkaline phosphatase was used as an apical membrane marker. Enrichment of alkaline phosphatase activity was more than 20-fold for MVM and LMVM compared with the activity of the homogenate. The cross-contamination of purified basal membranes with apical membranes was low, as evidenced by the lack of alkaline phosphatase enrichment in the basal fraction. The ratio of placental alkaline phosphatase activity enrichment of BM compared with that of apical membranes was 0.1 for the placentae used in this study, lower or equivalent to that from several other reports for single or paired apical and basal membrane preparations (Glazier et al., 1990; Illsley et al., 1990) as we demonstrated previously (Jimenez et al., 2004). In addition, for both MVM and BM vesicles, orientation is preferentially right-side out, as reported previously (Illsley et al., 1990). For LMVM vesicles, data about orientation are not available. However, it is expected that after this isolation protocol, all three type of vesicles show a preferential right-side out orientation. In fact, when setting up these methods, we initially observed that preparations, including both MVM and LMVM fractions, showed at least 90% of properly oriented vesicles, as determined by measuring phosphatase alkaline activity before and after saponin treatment (Barros, 1993), suggesting a preferential right-side out orientation for LMVM vesicles.
Nucleoside Transport Assay.
Uptake studies in syncytiotrophoblast-derived plasma membrane vesicles were performed using the rapid filtration technique, as described previously (Ruiz-Montasell et al., 1992). In brief, uptake was initiated by mixing 10 μl of vesicle suspension with 40 μl of uptake medium (250 mM sucrose, 0.2 mM CaCl2, 20 μM MgCl2, and 10 mM HEPES, pH 7.5) containing either 150 mM NaSCN or KSCN and 1 μM 3H-labeled substrate. Reactions were terminated at the indicated times by adding 1 ml of ice-cold stop solution (250 mM sucrose, 150 mM NaCl, 0.2 mM CaCl2, and 10 mM HEPES, pH 7.5) and then filtered through 0.45-μm nitrocellulose filters. Filters were washed with 4 ml of ice-cold stop solution, and radioactivity on filters was determined subsequently by liquid scintillation counting. All experimental values were corrected by subtracting nonmediated uptake values obtained by adding the stop solution into the transport before the vesicles. In addition, to distinguish between hENT1 and hENT2 activities, 1 μM nitrobenzylthioinosine (NBTI), a specific ENT1 inhibitor, was added to the transport medium.
Preparation of Apical Lipid Microdomains.
Apical plasma membrane microdomains were isolated from MVM- and LMVM-enriched preparations as a separate detergent-resistant fraction by extraction with Triton X-100, as described by Godoy and Riquelme (2008). In brief, aliquots (0.6 mg) of both isolated membrane fractions were homogenized 30 times in a manual glass homogenizer with 1% Triton X-100 in MES-buffered saline (25 mM MES, pH 6.5, and 150 mM NaCl). After incubation for 90 min on ice, vesicles (1 ml) from both apical preparations (LMVM and MVM) were mixed with 1 ml of 80% sucrose to obtain a final sucrose concentration of 40%. A discontinuous gradient was then prepared by overlaying the 40% cushion successively with 2 ml of 35% sucrose and 1 ml of 5% sucrose, and then tubes were centrifuged at 21,700g for 20 to 22 h in a swinging-bucket rotor (AH-650 Sorvall; Thermo Fisher Scientific). After centrifugation, 10 fractions (0.5 ml each) were collected starting at the top of the gradient.
Immunohistochemistry.
Samples of human placental tissue from normal pregnancies were rinsed in 0.9% NaCl, frozen in Cryogel Cancer Diagnostics, Inc., Birmingham, UK), and sectioned. Sections (10-μm-thick) were fixed with 4% paraformaldehyde for 30 min and rinsed three times with Tris-buffered saline, pH 7.6, for 5 min. In parallel, 3.7% buffered formalin-fixed placentae were processed, and paraffin was embedded according to standard protocols. The 5-μm sections thus obtained were subsequently deparaffinized and rehydrated. Slides containing both types of tissue sections (fixed and paraffin-embedded) were treated with 0.1 M citrate buffer, pH 6.0 (antigen recovery treatment), and then were incubated with 4% bovine serum albumin for 1 h to block nonspecific binding. Slides were incubated overnight at 4°C in a humidified chamber with primary polyclonal antibodies (diluted 1:100) against hENT1, hENT2, or hCNT1 (Farré et al., 2004). Slides stained for individual transporters were costained with mouse anti-cytokeratin 7 antibody (clone OVTL 12/30; Zymed Laboratories, South San Francisco, CA) diluted 1:100 in phosphate-buffered saline (PBS), pH 7.4, to confirm the trophoblastic localization of nucleoside transporters. After tissue sections were rinsed with PBS, they were incubated for 1 h at room temperature with Rhodamine Red X-conjugated goat anti-rabbit secondary antibody (1:200; Jackson ImmunoResearch Laboratories, West Grove, PA), which recognizes anti-hENT1, anti-hENT2, and anti-hCNT1), and with Cy2-conjugated goat anti-mouse antibody (Jackson ImmunoResearch Laboratories), which recognizes the anti-cytokeratin 7 antibody. Control sections were incubated with secondary antibody after incubation in PBS buffer without the primary antibody. Sections were viewed using a Leica TCS SP5 laser scanning confocal microscope (Leica Microsystems Heidelberg GmbH, Mannheim, Germany) equipped with a DMI6000 inverted microscope, a diode-pumped solid-state argon laser (561 nm), and a 63× oil immersion objective (numerical aperture 1.4). Cy2 and Rhodamine Red X labeling was visualized by acquiring images sequentially at the 488- and 561-nm laser lines using an Acousto-Optical Beam Splitter, with emission detection at 500 to 550 and 571 to 650 nm, respectively. Optical sections were collected every 0.3 μm in a 1024 × 1024 format with a confocal pinhole (radius) of 1 airy unit, zoom = 4, and a pixel size of 60 × 60 nm. Differential interference contrast images were acquired simultaneously.
Western Blot Analysis.
All flotation gradient fractions were evaluated by SDS-polyacrylamide gel electrophoresis and immunoblotting. An aliquot (50 μl) of each fraction was incubated with 10% (v/v) trichloroacetic acid for 30 min on ice and centrifuged at 21,000g for 30 min at 4°C. The pellets were resuspended in sample buffer and sonicated for 30 min. Trichloroacetic acid-precipitated protein (40 μg) from LMVM and MVM preparations was separated on 10% polyacrylamide gels and transferred to nitrocellulose membranes (Bio-Rad Laboratories, Hercules, CA). Membranes were incubated with primary anti-hENT1, anti-hENT2, or anti-hCNT1 antibodies diluted 1:1000. In addition, anti-human placental alkaline phosphatase (clone 8B6, 1:1000; Sigma-Aldrich) and anti-human transferrin receptor (clone H68.4, 1:500; Zymed Laboratories) were used as raft and nonraft markers, respectively. After incubation with primary antibody, proteins were detected using a horseradish peroxidase-conjugated secondary antibody and an enhanced chemiluminescence detection kit (GE Healthcare).
Data Analysis.
Data are expressed as the mean ± S.D. of uptake values obtained in three filter inserts from three independent placentae. Data are representative of three experiments carried out on different days on different batches of membranes.
Results
A study of nucleoside transporter-related mRNA showed that the five nucleoside transporters analyzed (hCNT1, hCNT2, hCNT3, hENT1, and hENT2), as well as the recently characterized spliced isoform of hCNT3 (hCNT3ins), were expressed in human placenta (Fig. 1).

Qualitative RT-PCR analysis of nucleoside transporters in human placenta. RT-PCR was used to amplify hCNT1, hCNT2, and hCNT3 (top) and hENT1 and hENT2 (bottom) from human placenta (P). Kidney (K), liver (L), and pancreas (Pc) were used as positive controls for hCNT1, 2, and 3, respectively, and kidney was used as a positive control for both hENTs. Positive controls for both wild-type (446-bp) and the recently reported hCNT3 splicing isoform hCNT3ins (622-bp) are shown. Representative agarose gels showing the amplification of cDNA fragments of the anticipated size are shown. C−, negative control.
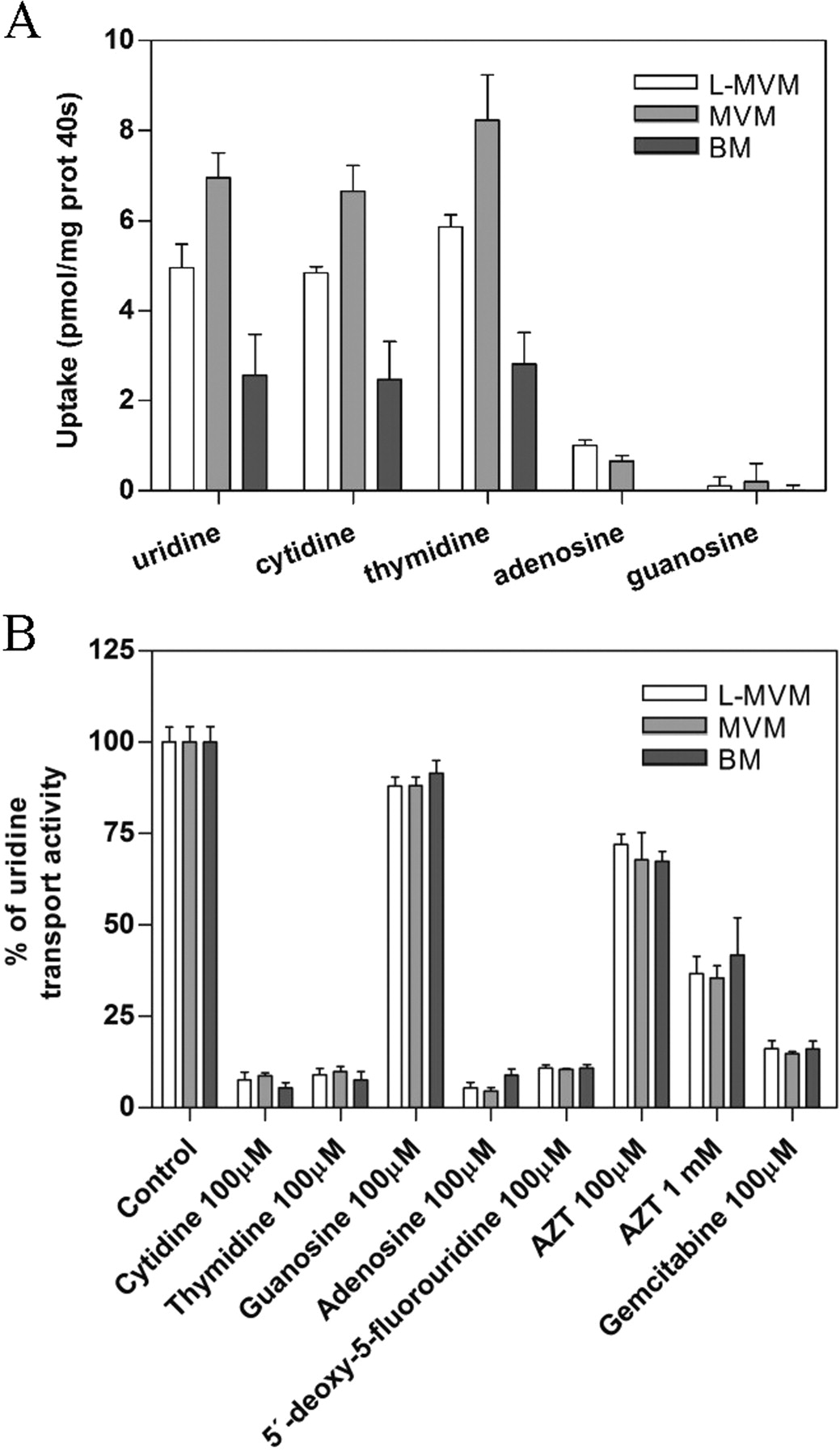
Nucleoside transport in syncytiotrophoblast plasma membrane-derived vesicles was characterized using either uridine or cytidine as substrates. Figure 2 shows the time course of uridine (Fig. 2A) and cytidine (Fig. 2B) uptake (1 μM) by vesicles derived from human placental LMVM (top), MVM (middle), and BM (bottom) (n = 3 independent placentae). Both sodium-dependent and -independent components were observed. Inhibition of sodium-independent uptake with NBTI revealed that both NBTI-sensitive and -insensitive carriers, hENT1 and hENT2, respectively, appear functional in these membrane preparations (Fig. 2), in agreement with the pattern of mRNA expression detected in this tissue (Fig. 1). Sodium-dependent transport of both nucleosides into plasma membrane vesicles was observed when uptake was measured in the presence of an outside-to-inside sodium gradient (150 mM, out > in), similar to that reported in renal and intestinal brush-border membrane vesicles (Gutierrez and Giacomini, 1993; Ngo et al., 2001). Because nucleoside uptake was linear up to at least 40 s and this time point provided greater sensitivity, values obtained at 40 s are reported for all experiments conducted with placental syncytiotrophoblast plasma membrane vesicles. To analyze to which transport agent this sodium-dependent component was attributable, the profiles of 3H-labeled natural nucleoside uptake in LMVM, MVM, and BM vesicles and inhibition of [3H]uridine uptake by nucleosides and nucleoside-derived drugs were analyzed (Fig. 3). Measurements of [3H]uridine, [3H]cytidine, [3H]thymidine, [3H]adenosine, and [3H]guanosine uptake in the presence of a sodium gradient demonstrated the existence of sodium-dependent transport for all three pyrimidine nucleosides in both apical and basal membrane vesicles (Fig. 3A). Sodium-coupled adenosine uptake in LMVM and MVM vesicles, although small, was significant, whereas sodium-dependent guanosine uptake was negligible. Moreover, sodium-dependent uptake of [3H]uridine (1 μM) was completely inhibited by 100 μM cytidine, thymidine, and adenosine but was not significantly blocked by 100 μM guanosine (n = 3 independent placentae) (Fig. 3B). The nucleoside derivatives, 5′-deoxy-5-fluorouridine and gemcitabine (100 μM), also significantly inhibited [3H]uridine uptake into apical and basal membrane vesicles, as did the nucleoside analog zidovudine, albeit to a lesser extent (Fig. 3B). All these results indicate that hCNT1 is the unique concentrative nucleoside transport activity present in syncytiotrophoblast plasma membrane-derived vesicles.

Time course of nucleoside uptake. Time course of [3H]uridine (A) and [3H]cytidine (B) (1 μM) uptake in syncytiotrophoblast LMVM (top), MVM (middle), and BM (bottom) vesicles. Uptake into vesicles was assayed in medium containing radioactive uridine or cytidine and either 150 mM sodium (NaSCN, ■) or potassium (KSCN, ▴) as described under Materials and Methods. Equilibrative nucleoside transport activities were identified by means of NBTI (1 μM) inhibition (▾). Results are presented as means ± S.D. of triplicate estimations made on three independent preparations.
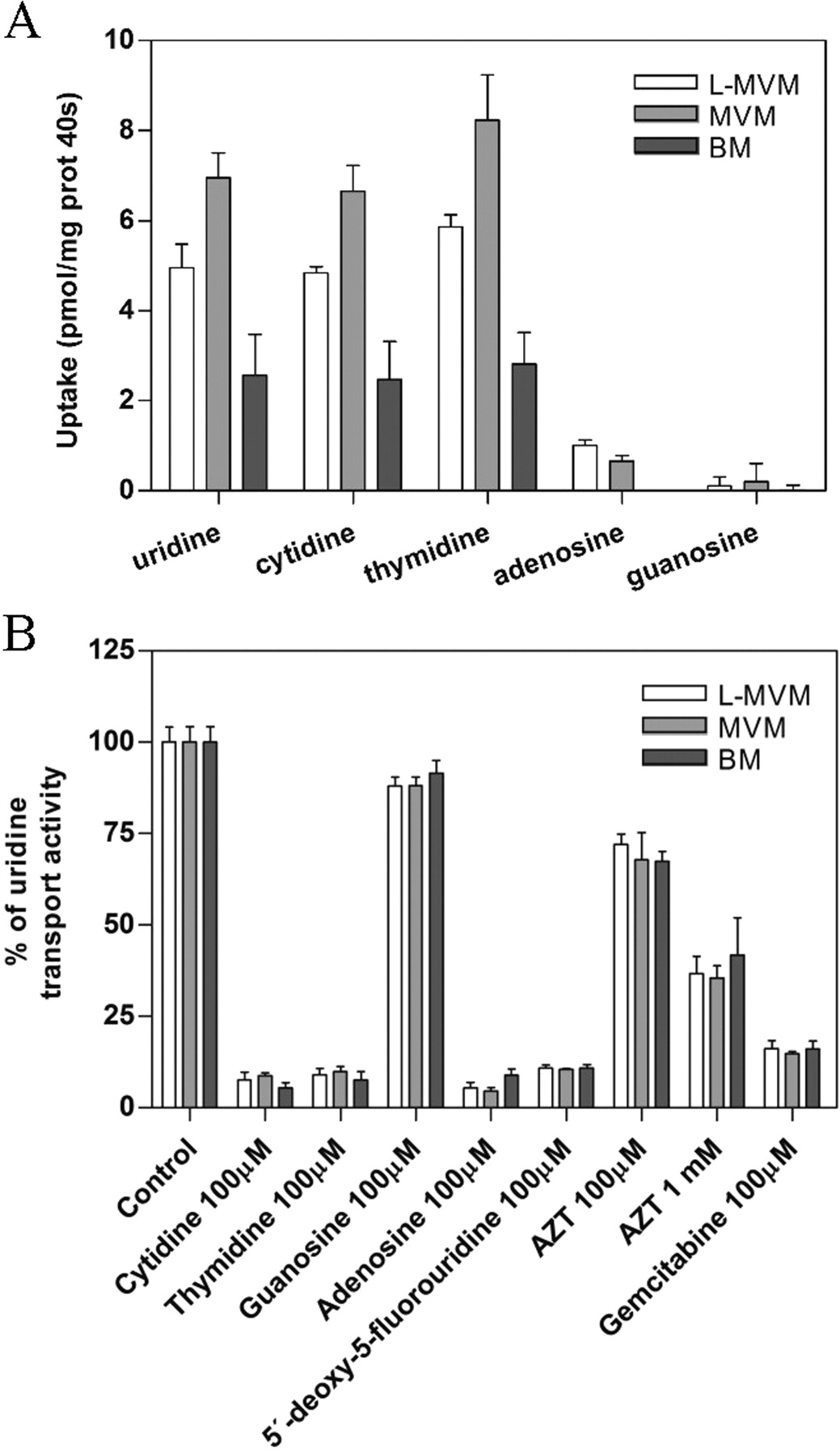
Characterization of CNT activity in plasma membrane vesicles. A, sodium-dependent uptake of pyrimidine and purine nucleosides (1 μM) by LMVM, MVM, and BM at 40 s was measured in transport medium containing 150 mM NaSCN or 150 mM KSCN. Sodium-dependent transport was calculated as uptake in sodium medium minus uptake in potassium medium. B, inhibition profile of sodium-dependent [3H]uridine uptake (1 μM) by LMVM, MVM, and BM at 40 s. Data are presented as a percentage of uridine transport normalized to the control. Transport experiments in A and B were performed as in the legend to Fig. 2. Data are expressed as the mean ± S.D. of triplicate estimations made on three independent preparations.
The immunohistochemical analysis of human placental tissue sections showed that hCNT1, hENT1, and hENT2 were expressed in the maternal-facing side of the syncytiotrophoblast (Fig. 4, A–C). Although some immunofluorescence was evident in intracellular compartments, especially in some nuclei, staining was clearly most apparent at the syncytiotrophoblast apical membrane. Increasing exposure to the laser source revealed light staining of hENT2 and hCNT1 in the basal membranes, but basal membrane staining of hENT1 was not detected under any laser exposure condition tested (data not shown). Double immunostaining with cytokeratin 7 confirmed the localization of these transporters to microvillous and basal syncytiotrophoblast membranes, demonstrating plasma membrane localization of nucleoside transporter proteins (Fig. 4, D–I). No significant fluorescence was observed in slides treated with secondary antibodies only (data not shown).

Expression of hCNT1, hENT1, and hENT2 proteins in human term placenta. Confocal fluorescence micrograph of immunohistochemical sections of a normal-term human placenta showing hCNT1 (A), hENT1 (B), and hENT2 (C) staining of the apical syncytiotrophoblast membrane. Double immunostaining with an anti-cytokeratin 7 (Cyt7) antibody (D, E, and F) shows intense staining of all three transporter proteins at the apical microvillous membrane of the syncytiotrophoblast (MVM) and in intracellular structures (G, H, and I).
The presence of distinct lipid rafts in the apical plasma membrane domain of placental syncytiotrophoblast has been reported (Godoy and Riquelme, 2008), although the presence of nutrient transporters, particularly nucleoside transporters, in these domains has not been established. In this regard, Western blot detection of nucleoside transporter proteins in purified syncytiotrophoblast membranes and analysis of the distribution of hCNT1, hENT1, and hENT2 in different lipid microdomains were performed. Nucleoside transporter proteins in paired samples of purified apical and basal fractions from four placentae were analyzed by Western blotting. Figure 5A shows a representative Western blot with bands corresponding to the molecular weight of hCNT1 (top), hENT1 (middle), and hENT2 (bottom) in both apical LMVM and MVM preparations as well as BM preparations. In agreement with uptake assay and immunohistochemical results, hCNT1 and hENT2 were present in all apical and basal purified fractions, whereas hENT1 was present only in light and heavy apical membranes.
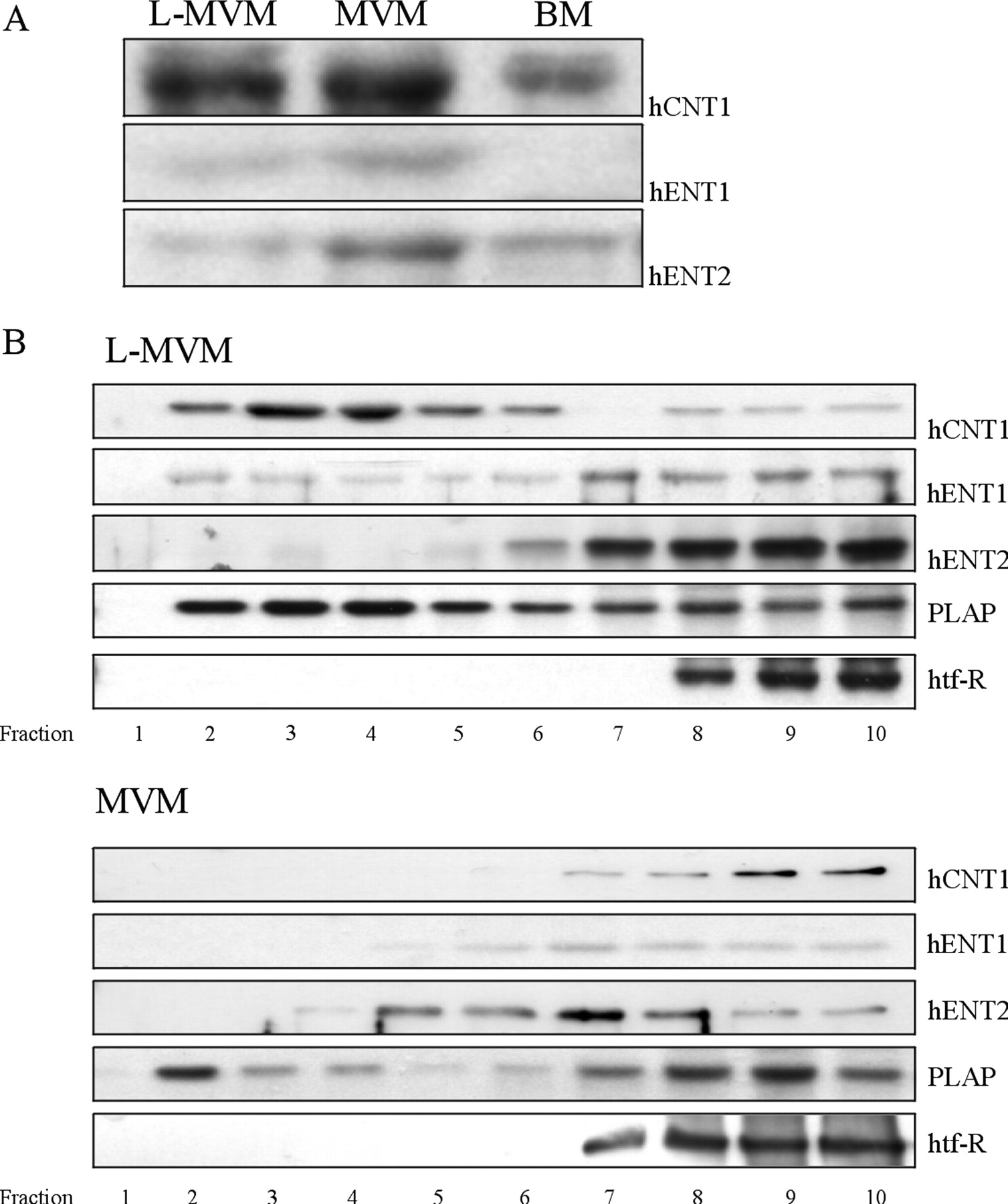
Distribution of nucleoside transporters in raft and nonraft fractions isolated from LMVM and MVM preparations. Lipid rafts were isolated from apical membrane vesicles, prepared from normal-term human placentae, by solubilizing in ice-cold 1% Triton X-100 and fractionating on a sucrose gradient. A, immunoblot analysis shows that both hCNT1 and hENT2 are present in apical and basal purified membrane-derived vesicles, whereas hENT1 is present only in apical membrane vesicles. B, LMVM and MVM sucrose gradient fractions were separated by SDS-polyacrylamide gel electrophoresis and analyzed by Western blotting using antibodies against hCNT1, hENT1, and hENT2, with placental alkaline phosphatase (PLAP) and human transferrin receptor (htf-R) used as raft and nonraft markers, respectively. Top panel, a representative Western blot of LMVM samples showing the distribution of hCNT1 and hENT1 into both lipid raft and nonraft fractions and exclusive distribution of hENT2 to nonraft fractions. Bottom panel, Western blot of MVM samples showing localization of hCNT1 and hENT1 to nonraft fractions and distribution of hENT2 to some raft and all nonraft fractions.
To determine whether hCNT1, hENT1, and hENT2 are localized to membrane rafts, we isolated a detergent-resistant membrane fraction by sucrose density centrifugation, as described recently (Godoy and Riquelme, 2008). In Western blots of LMVM-containing gradient fractions (Fig. 5B), hCNT1 and hENT1 immunoreactivity was detected in both detergent-resistant (lipid raft) and -nonresistant fractions, although hCNT1 mainly partitioned into lipid raft fractions, and hENT1 was predominantly associated with nonlipid raft fractions. In contrast, hENT2 was totally absent from the detergent-resistant membranes of the LMVM flotation fractions, indicating that hENT2 does not localize to lipid rafts in this fraction. Western blotting of gradient fractions enriched for MVM showed a completely different distribution of both hCNT1 and hENT1 transporter proteins. Here, both transporters were absent from the low sucrose density fractions and were found exclusively in nonraft fractions. hENT2 immunoreactivity was found mainly in nonraft fractions, although it could be detected in some, but not all, lipid raft fractions (Fig. 5B).
Discussion
Maternofetal transfer of nutrients such as glucose and amino acids is mostly determined by the activity of specific transporter proteins expressed in the placental syncytiotrophoblast plasma membranes. Glucose transport from the mother to the fetus seems to be influenced by the maternal blood glucose concentration across the placenta and is mediated by members of the facilitative glucose transporters (Baumann et al., 2002), although evidence for SLGT1-mediated Na+-coupled transport in the syncytiotrophoblast has also been provided (Kevorkova et al., 2007). The maternofetal transfer of most amino acids is dependent on the coupling of secondary active amino acid transporters localized in the microvillous apical membrane of the syncytiotrophoblast (in direct contact with the maternal blood) and facilitative transporters present in the basal membrane (facing the fetal circulation) (Grillo et al., 2008). This model of nutrient vectorial flux is to some extent similar to the one described for nucleosides in (re)absorptive epithelia (intestinal and renal epithelial cells), also involving the asymmetric distribution of concentrative (CNT-type) and equilibrative (ENT-type) nucleoside transporter proteins in apical and basolateral domains, respectively, although some ENT1 expression at the apical side has also been reported (Errasti-Murugarren et al., 2007).
Of interest, the “absorptive model” for the vectorial flux of nucleosides is not the only one that could be applied to the syncytiotrophoblast epithelium. Previous evidence supported apical and basal localization of equilibrative nucleoside transport activities and transporters in the syncytiotrophoblast (Barros et al., 1995; Govindarajan et al., 2007). This result is further reinforced in this contribution by combining functional data and hENT1 and hENT2 immunodetection. hENT2 was present in both domains (apical and basal) when determined by Western blot, although this pattern was not exactly reproduced when hENT2 expression was investigated with immunohistochemistry. Putative post-translational modifications of the protein, perhaps related to its polarized sorting, might interfere with staining procedures. Regarding hENT1, its apical localization was corroborated by Western blot experiments. However, in agreement with previous observations (Barros et al., 1995; Govindarajan et al., 2007), a hENT1-like activity identified at the basal plasma membrane vesicles was not associated with a detectable hENT1 protein, as measured by either Western blot or immunohistochemical analysis. The possibility of two distinct isoforms of the hENT1 protein in the syncytiotrophoblast apical and basal membranes should be further evaluated. In fact, splicing isoforms of the SLC29A1 gene could affect both protein immunodetection and/or subcellular localization in polarized epithelia as previously reported for the SLC28A3 gene product (Errasti-Murugarren et al., 2009). As an alternative, there is the possibility that immunoreactivity using antibodies raised against the intracellular loop between transmembrane domains 6 and 7 might be impaired if significant post-translational modification occurs in the loop (i.e., phosphorylation). In any case, it seems that transport activities consistent with the occurrence of both types of transporter proteins (hENT1 and hENT2) are present at both poles of this epithelium.
Nevertheless, and more importantly, this study also demonstrates that a concentrative nucleoside transport activity, mostly, if not exclusively, linked to the expression of the pyrimidine-preferring nucleoside transporter hCNT1 is found at both the microvillous and basal membranes of the syncytiotrophoblast. This represents a unique distribution of CNT-type proteins in a polarized epithelium. The occurrence of hCNT1 is based on the selectivity profile of the uptake of natural nucleosides and the cis inhibition of uridine uptake by nucleosides into apical and basal plasma membrane vesicles. In this case, the functional evidence is strongly supported by hCNT1 immunodetection. In fact, immunohistochemical analysis of placental sections further identified hCNT1 at both poles of the syncytiotrophoblast. The low staining for hCNT1 at the basal side is probably due to its comparatively low expression in this membrane domain, as confirmed by the activity measurements and the Western blot analysis of the subcellular membrane fractions.
All these observations taken together would be consistent with the model of NT protein distribution in the syncytiotrophoblast shown in Fig. 6, a model that rules out the occurrence of hCNT2/3 functional expression, at least in placentae at term. Moreover, residual adenosine uptake found in vesicles would be consistent with previous evidence showing that adenosine is a high-affinity hCNT1 inhibitor (with a Ki value in the low micromolar range) but a poor permeant (Larráyoz et al., 2004; Smith et al., 2004), suggesting that endogenous maternal adenosine levels could reduce interaction of hCNT1 with nucleoside-derived drugs, reducing in that way their fetal toxicity. In agreement with previous observations (Yamamoto et al., 2007), all five NT-related mRNAs could be found in placenta. This observation is consistent with either CNT2/3 transporters being expressed in other cell types or being developmentally regulated in the syncytiotrophoblast or both. Nevertheless, the occurrence of hCNT1 as the unique concentrative nucleoside transporter in this epithelium has a physiological rationale. The expression levels (mRNA transcript amounts) of the enzymes implicated in both pyrimidine and purine nucleoside metabolism in placenta have been searched in the Unigene database (Sayers et al., 2009). The enzyme machinery expressed in placenta is consistent with high purine but low pyrimidine nucleotide de novo synthesis. Of interest, the enzyme machinery implicated in salvage pathways shows the opposite pattern: the enzymes responsible for pyrimidine recycling are highly expressed. This scenario is consistent with the need for a plasma membrane pyrimidine-preferring concentrative nucleoside transporter (hCNT1) to mediate pyrimidine but not purine salvage.

Model of nucleoside transporter protein distribution in the syncytiotrophoblast. Integrated model of nucleoside, amino acid, and glucose transporter distribution in human placenta together with expression levels of nucleoside metabolism key enzymes. PRPS1/2, phosphoribosyl pyrophosphate synthase 1/2; CAD, carbamoyl phosphate synthetase II/aspartate transcarbamoylase/dihydro-orotase; DD, dihydroorotate dehydrogenase; UP, uridine phosphorylase 1; UCK, uridine-cytidine kinase; TK, thymidine kinase; TP, thymidine phosphorylase; PyN, pyrimidine nucleoside; PN, purine nucleoside; NB, nucleobase; NP, nucleoside phosphate; AAT, amino acid transporter.
It is not evident whether this NT distribution pattern will in turn confer vectoriality to nucleoside transport across the placenta. The fetus is highly dependent on glucose and amino acids, which are the building blocks of purine and pyrimidine nucleotides and de novo synthesis is highly prevalent in the growing fetus (Alexiou and Leese, 1992; Boza et al., 1996). In this context, it is tempting to speculate that both apical and basal hCNT1 are implicated in the pyrimidine nucleoside supply to the placenta, from both maternal and fetal sides. This speculation would be consistent also with the expression pattern of hENT2, present at both poles, being the only NT protein known to mediate transport of nucleobases, which could also be used for pyrimidine salvage purposes (Fig. 6). As discussed above, this model is based on results obtained in term placenta. A previous report showed slight variations in thymidine uptake (a hCNT1 substrate) along different gestational stages (Sooranna et al., 1999). In any case, this was total uptake and does not rule out the possibility that the contribution of particular nucleoside transporter proteins to nucleoside uptake changes during placental growth. This may be of interest in the understanding of placental physiology but difficult to address in human studies.
As introduced above, hCNT1 is a high-affinity transporter for selected anticancer drugs, such as gemcitabine and 5-azacytidine (García-Manteiga et al., 2003; Rius et al., 2009). These drugs have been reported to induce a variety of harmful effects in rodent placenta and developmental toxicity as well (Eudaly et al., 1993; Serman et al., 2007). Moreover, inhibition of hCNT1 by selected drugs may compromise pyrimidine salvage by the syncytiotrophoblast, thus contributing to impaired placental function. Conclusions drawn from the use of cell lines expressing exclusively low-affinity ENT-type transporters should be made cautiously and reevaluated (Chishu et al., 2008).
Finally, the occurrence of selected hNT proteins in raft microdomains might be relevant also in functional terms. We have recently shown that localization of hCNT3 in lipid rafts might determine its biological activity (Errasti-Murugarren et al., 2010), and, interestingly, CNT1 protein had also been identified in a caveolin-enriched plasma membrane fraction isolated from quiescent rat liver (Duflot et al., 2002). On the basis of this previous evidence, the analysis of the localization of hNT proteins in lipid rafts was performed. As described above, Western blotting of the LMVM gradient showed a bimodal distribution for hCNT1 and hENT1 immunoreactivities, being detected both in lipid raft and nonlipid raft fractions, whereas hENT2 was totally absent in raft fractions. Of interest, the heavy apical MVM showed a completely different distribution for both hCNT1 and hENT1 transporter proteins, being found exclusively in nonraft fractions. The heavy MVM and LMVM fractions used in this study correspond to the finger-like projections and the bottom part between the microvilli, respectively (Godoy and Riquelme, 2008), which also show a heterogeneous population of raft domains (Braccia et al., 2003; Godoy and Riquelme, 2008). The expression of particular cytoskeletal proteins in finger-like projection regions of the microvilli (MVM) that has been proposed to be tightly associated with plasma membrane lipidic microdomains could explain the presence of lipid raft markers in nonraft fractions (Godoy and Riquelme, 2008). In this regard, both hCNT1 and hENT1 transporters in MVM are located mainly in nonraft fractions, which are clearly defined by the human transferrin receptor, whereas distribution of hENT2 protein is less clear. All these observations, taken together, suggest that nucleoside transporter proteins show heterogeneous distribution not only in polarized epithelia but also at the subcellular level in particular apical domains. Although the elucidation of the physiological rationale for this distribution and the effect on nucleoside transport activity requires further analysis, it is interesting to point out that preliminary data from our laboratories (E. Errasti-Murugarren, unpublished observations) indicate that in some pathological conditions, such as pre-eclampsia, disruption of lipid raft domains will result in altered distribution of hNT proteins, thus probably affecting their biological function.
In summary, this study provides the first comprehensive analysis of nucleoside transporter expression in the human syncytiotrophoblast. The only concentrative nucleoside transporter protein expressed at significant levels in this epithelium at term is hCNT1, a pyrimidine-preferring, sodium-coupled concentrative nucleoside transporter. Its occurrence at both poles of the epithelium, basal and apical, is consistent with the fact that the placenta has the ability to synthesize purine nucleotides de novo but seems to be dependent on pyrimidine nucleoside salvage. Thus, hCNT1 would be a key player in this process. Considering that some anticancer nucleoside-derived drugs and xenobiotics such as nicotine and caffeine are either translocated by or bound to hCNT1 with high affinity, we suggest that most of the harmful effects triggered by these drugs can be mediated, at least in part, by their ability to interact with hCNT1, thus impairing the pyrimidine nucleoside salvage process.
Authorship Contributions
Participated in research design: Errasti-Murugarren, Riquelme, and Pastor-Anglada.
Conducted experiments: Errasti-Murugarren, Díaz, and Godoy.
Performed data analysis: Errasti-Murugarren, Díaz, Riquelme, and Pastor-Anglada.
Wrote or contributed to the writing of the manuscript: Errasti-Murugarren, Díaz, Riquelme, and Pastor-Anglada.
Acknowledgements
We express our gratitude to Dr. M. Pérez and the staff of the San José Hospital Maternity Unit (Santiago, Chile) for their assistance in obtaining biological material and the Confocal Microscopy Facility of Serveis Cientificotècnics (Universitat de Barcelona-Institut d'Investigacions Biomèdiques August Pi i Sunyer) for their support and advice with confocal techniques. We also thank C. Vallejos for continuous support in the preparation of plasma membranes, A. Valdebenito for technical assistance, and Dr. Felipe Barros (Centro de Estudios Científicos, Valdivia, Chile) for helpful discussions.
Footnotes
This research was supported in part by Fondecyt-Chile [Grants 1070695 and SAF2008-00577] (the former to G.R.); Centro de Investigación Biomédica en Red (an initiative of Instituto de Salud Carlos III); Generalitat de Catalunya [Grant 2009SGR00624] (to M.P.-A.); and the Ministerio de Ciencia e Innovación [Grant AP2003–3938] (to E.E.-M.).
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.111.071837.
-
ABBREVIATIONS:
- h
- human
- CNT
- concentrative nucleoside transporter
- ENT
- equilibrative nucleoside transporter
- NT
- nucleoside transporter protein
- RT
- reverse transcriptase
- PCR
- polymerase chain reaction
- bp
- base pair(s)
- LMVM
- light microvillous membrane
- MVM
- microvillous membrane
- BM
- basement membrane
- NBTI
- nitrobenzylthioinosine
- MES
- morpholinoethanesulfonic acid
- PBS
- phosphate-buffered saline.
- Received February 18, 2011.
- Accepted August 8, 2011.
- Copyright © 2011 The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}