


Abstract
The WNT/β-catenin signaling pathway has been identified as an important endogenous regulator of hepatic cytochrome P450 (P450) expression in mouse liver. In particular, it is involved in the regulation of P450 expression in response to exposure to xenobiotic agonists of the nuclear receptors constitutive androstane receptor (CAR), aryl hydrocarbon receptor (AhR), and Nrf2. To systematically elucidate the effect of the WNT/β-catenin pathway on the regulation and inducibility of major human P450 enzymes, HepaRG cells were treated with either the WNT/β-catenin signaling pathway agonist, WNT3a, or with small interfering RNA directed against β-catenin, alone or in combination with a panel of activating ligands for AhR [2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)], CAR [6-(4-chlorophenyl)imidazo[2,1-b][1,3]thiazole-5-carbaldehyde-O-(3,4-dichlorobenzyl)oxime (CITCO)], pregnane X receptor (PXR) [rifampicin], and peroxisome proliferator–activated receptor (PPAR) α [4-chloro-6-(2,3-xylidino)-2-pyrimidinylthioacetic acid (WY14,643)]. Assessment of P450 gene expression and enzymatic activity after downregulation or activation of the WNT/β-catenin pathway revealed a requirement of β-catenin in the AhR-, CAR-, and PXR-mediated induction of CYP1A, CYP2B6 and CYP3A4 (for CAR and PXR), and CYP2C8 (for PXR) gene expression. By contrast, activation of the WNT/β-catenin pathway prevented PPARα-mediated induction of CYP1A, CYP2C8, CYP3A4, and CYP4A11 genes, suggesting a dominant-negative role of β-catenin in PPARα-mediated regulation of these genes. Our data indicate a significant effect of the WNT/β-catenin pathway on the regulation of P450 enzymes in human hepatocytes and reveal a novel crosstalk between β-catenin and PPARα signaling pathways in the regulation of P450 expression.
Introduction
The WNT/β-catenin signaling pathway, traditionally well known as a regulator of embryonic development, tissue homeostasis, and carcinogenesis, was recently identified as an important endogenous regulator of hepatic drug metabolism in mouse liver, where the activity of this particular pathway determines hepatic zonation of cytochrome P450 (P450) expression (Loeppen et al., 2005; Hailfinger et al., 2006; Sekine et al., 2006; Braeuning and Schwarz, 2010; Braeuning et al., 2011a; Schreiber et al., 2011) and is also critically involved in the regulation of P450 transcription in response to exposure to xenobiotic agonists of a number of nuclear receptors (Braeuning et al., 2009; Ganzenberg et al., 2013; Vaas et al., 2014). Lipid- and xenobiotic-sensing nuclear receptors pregnane X receptor (PXR, NR1I2), constitutive androstane receptor (CAR, NR1I3), and aryl hydrocarbon receptor (AhR), together with the liver-enriched transcription factor peroxisome proliferator–activated receptor (PPAR) α (NR1C1) form networks of transcription factors that coordinately regulate hepatic expression of the majority of drug-metabolizing P450s, phase II enzymes, and transporters in response to xenobiotic exposure (Handschin and Meyer, 2005; Pascussi et al., 2008; Pelkonen et al., 2008; Thomas et al., 2013). Recent research elucidated extensive crosstalk between these receptors and other nuclear receptors and transcription factors, linking xenobiotic metabolism to the homeostasis of lipids, bile acids, glucose, and other endogenous processes (Moreau et al., 2008; Gao and Xie, 2012). For WNT/β-catenin signaling in general, it has been observed that activation of the pathway in murine liver cells stimulates the expression of most P450s and augments their inducibility by xenobiotic agonists of nuclear receptors in vivo and in vitro. Further studies link the β-catenin pathway to elevated expression of some P450s in human hepatoblastoma (Schmidt et al., 2011) and primary human hepatocytes (PHHs), underlining the relevance of these findings also for humans. In particular, it was found that the WNT/β-catenin pathway activation induces the expression of CYP2E1, CYP1A2, and AhR, but not of CYP3A4, hepatocyte nuclear factor-4α, or PXR in PHHs (Gerbal-Chaloin et al., 2014). However, thus far there are no systematic data available on the effect of the WNT/β-catenin pathway on the regulation of basal or xenobiotic-induced P450 gene expression and activities in human cells.
Furthermore, although the interactions of signal transduction through β-catenin and the drug-sensing receptors PXR, CAR, and AhR have been studied previously, a potential interaction of β-catenin with PPARα remains to be established. This nuclear receptor acts as a lipid sensor to control the expression of gene networks involved in lipid and energy homeostasis, adipocyte differentiation, and inflammatory responses (Lalloyer and Staels, 2010; Wahli and Michalik, 2012). We recently demonstrated that PPARα directly regulates the transcription of CYP3A4 (Klein et al., 2012; Thomas et al., 2013). Transcriptional activation of other drug biotransformation genes by PPARα has already been reported for the fatty acid ω–hydroxylase, CYP4A11 (Johnson et al., 2002; Wanders et al., 2011), several UDP-glucuronosyltransferases (Barbier et al., 2003; Senekeo-Effenberger et al., 2007), and some drug transporters (Cheng et al., 2005; Moffit et al., 2006). Our comparison of ligand-induced gene expression changes mediated by PPARα and the two prototypic xenosensors PXR and CAR further emphasizes the similar target profiles of these three nuclear receptors with respect to the regulation of drug biotransformation genes in the liver (Thomas et al., 2013). Thus, PPARα is a relatively novel player in the regulation of drug-metabolizing enzymes, and PPARα activity in linking regulatory pathways of intermediary and xenobiotic metabolism warrants further study.
This study was therefore undertaken to analyze the relevance of β-catenin activation or inhibition on basal and inducible expression of a broad spectrum of P450 isoforms in the HepaRG human hepatocarcinoma cell line. When following a differentiation protocol including two weeks of cultivation in medium containing a high amount of dimethylsulfoxide, HepaRG cells differentiate into a mixed population of hepatocyte- and bile duct epithelium-like cells. Differentiated HepaRG cells possess several hepatocyte-specific functions and show exceptional expression and activity of many P450s relevant for the metabolism of drugs and xenobiotics (Aninat et al., 2006; Guillouzo et al., 2007; Klein et al., 2015). Our data suggest a significant effect of WNT/β-catenin signaling on basal as well as inducible expression of the major P450 genes. Moreover, perturbation of the WNT/β-catenin pathway revealed the ancillary role of β-catenin in AhR-, PXR-, and CAR-mediated induction but an inhibitory role of β-catenin in PPARα-mediated induction of target P450 enzymes (CYP1A, CYP2, CYP3, and CYP4 isoforms). We suggest intricate interactions between WNT/β-catenin signaling, lipid homeostasis, and drug biotransformation capacity that may be of clinical relevance under diseased conditions such as obesity, hepatic steatosis, and cancer.
Materials and Methods
Cell Culture and Treatment.
Cryopreserved differentiated HepaRG cells (Biopredic, Rennes, France) were seeded in 12-well plates at a density of 0.8 × 106 cells/well according to the supplier’s instructions. HepaRG cells were cultured in HepaRG maintenance/metabolism medium (Biopredic) for 4 days before treatment. At day 4 after seeding, cells were treated with 20 ng/ml WNT3a (dissolved in 0.1% bovine serum albumin; Sigma, Taufkirchen, Germany) in HepaRG serum-free induction medium (Biopredic), and WNT3a treatment was repeated without additional medium change at days 5 and 6. In addition, cells were exposed to the following known enzyme inducers at day 6 for an additional 24 hours, alone or in combination with WNT3a treatment and dissolved in dimethylsulfoxide: 10 nM 2,3,7,8-tetrachlorodibenzop-[p]-dioxin (TCDD; Ökometric, Bayreuth, Germany), 10 µM rifampicin (Sigma), 5 µM 6-(4-chlorophenyl)imidazo[2,1-b][1,3]thiazole-5-carbaldehyde-O-(3,4-dichlorobenzyl)-oxime (CITCO; Enzo Life Sciences, Loerrach, Germany), or 100 µM 4-chloro-6-(2,3-xylidino)-2-pyrimidinylthioacetic acid (WY14,643; gift from Dr. C. Gembardt, Ludwigshafen, Germany).
HuH7 cells were cultured at 37°C with 5% CO2 concentration and passaged every 3 to 4 days by the Trypsin/EDTA method. HuH7 cells were cultured in Dulbecco’s modified Eagle’s medium with 10% fetal calf serum gold (PAA Laboratories GmbH, Pasching, Austria), 1% penicillin/ streptomycin, and 1% pyruvate.
Transfections with Small Interfering RNAs.
For the RNA interference experiments, HepaRG cells were transfected with 20 nM small interfering RNAs (siRNAs) using 10 pmol Lipofectamine RNAiMAX Transfection Reagent (Life Technologies, Carlsbad, Germany) in 24-well plates with serum-free medium. The siRNA targeting β-catenin and a nontargeting siRNA as a negative control (Lo GC Duplex 2) were also obtained from Life Technologies. One-hundred microliters of the transfection cocktail was added per well to the cells containing 100 µl culture medium. Upon 20 minutes of complex formation, the liposomes were given to the cells. Twenty-four hours after the transfection, cells were treated for an additional 48 hours with the above-mentioned agents.
Quantitative Parallel Real-Time Polymerase Chain Reaction Using Fluidigm Biomark.
RNA integrity and quantity were analyzed with the Agilent 2100 Bioanalyzer using the RNA 6000 Nano Kit (Agilent Technologies, Waldbronn, Germany). Synthesis of cDNA was performed with 500 ng RNA using TaqMan Reverse Transcription Reagents (Applera GmbH, Darmstadt, Germany). Quantification of expression of 11 P450 genes was performed with Fluidigm’s BioMark HD high-throughput quantitative chip platform (Fluidigm Corporation, San Francisco, CA) following the manufacturer’s instructions (Spurgeon et al., 2008). All corresponding TaqMan assays were purchased from Life Technologies (Darmstadt, Germany). For the validation of siRNA, the expression of downstream genes of WNT signaling was determined using the following assays from Life Technologies: CTNNB1 and Axin 2. The mRNA expression levels were normalized to glyceraldehyde 3-phosphate dehydrogenase mRNA expression.
Measurement of P450 Enzyme Activity.
P450 enzyme activities were determined in HepaRG cell culture supernatants using a liquid chromatography with tandem mass spectrometry–based substrate cocktail assay, as described previously (Feidt et al., 2010).
Western Blot Analysis.
For the protein measurements, 50 µg total cell lysates was electrophoretically separated on a 10% SDS-polyacrylamide gel and subsequently transferred to a nitrocellulose membrane using a Trans-blot semi-dry Fastblot 44 transfer chamber (Biometra, Göttingen, Germany). After blocking with 5% skim milk in Tris-buffered saline/Tween 20, membranes were incubated with the primary antibodies in 1% skim milk solution in Tris-buffered saline/Tween 20. For the detection of the levels of proteins of interest, we used a mouse anti-human β-catenin antibody from Becton and Dickinson (1:500 dilution; Franklin Lakes, NJ). CYP3A4 protein was detected as described earlier (Wolbold et al., 2003). For normalization, mouse anti–β-actin (1:500; Sigma-Aldrich, St. Louis, MO) was used and goat anti–rabbit-IRD800 (1:10,000; Li-COR, Bad Homburg, Germany) and goat anti–mouse-IRD650 (1:10,000; Li-COR) were used as fluorescent-labeled secondary antibodies. Detection was performed with a Li-COR Odyssey CLx fluorescence reader.
Luciferase Reporter Gene Assays.
Human hepatocarcinoma HuH7 cells were transfected with 300 ng reporter pGL3 basic plasmid containing 8.8 kb of the CYP3A4 upstream promoter region (Tegude et al., 2007) using Turbofect (Fermentas, St. Leon-Rot, Germany) as previously reported (Thomas et al., 2013). As an internal control of transfection efficiency, 25 ng pGL3-TK-Renilla (Promega, Mannheim, Germany) was used. Cells were harvested in reporter “passive” lysis buffer (Promega) after 48 hours of incubation, and cell extracts were analyzed for firefly and Renilla luciferase activities using the Enspire Multimode Plate-Reader (PerkinElmer, Rodgau, Germany) and the Luciferase Assay System (P.J.K. GmbH, Kleinblittersdorf, Germany). Firefly luciferase activities were normalized to Renilla luciferase activities.
Statistical Methods.
For comparisons of two groups, the two-sample t test (two tailed) was applied. To identify statistically significant differences between more than two groups, one-way analysis of variance with the Student–Newman–Keuls post test was performed. Measurements of experiments performed in triplicate were averaged before analyses. Statistical significance was defined as P < 0.05. All calculations were performed using GraphPad Prism software (GraphPad Software Inc., San Diego, CA). Densitometric analyses were performed using the ImageJ software available online from the National Institutes of Health (Bethesda, MD).
Results
Modulation of WNT/β-Catenin Signaling in HepaRG Cells.
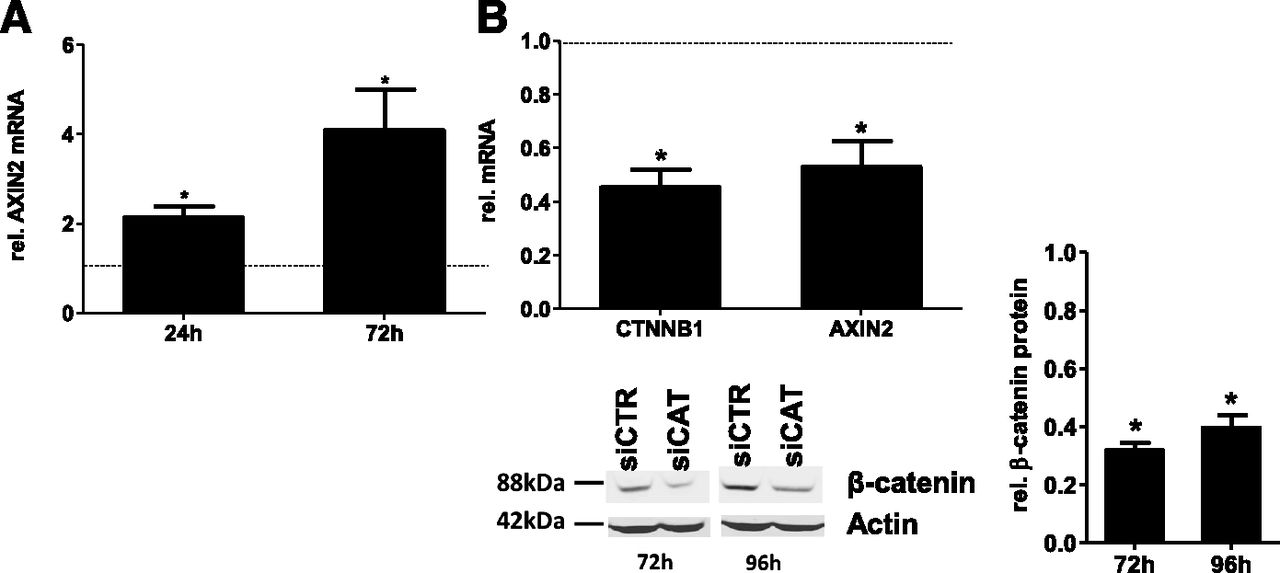
To follow the consequences of WNT pathway activation or inhibition on basal and inducible expression of a broad spectrum of P450 isoforms in the HepaRG human hepatocarcinoma cell line, we applied two available strategies to either activate or inactivate the pathway (i.e., using stimulation with the canonical natural WNT ligand, WNT3a, and depletion of β-catenin using siRNA). As shown in Fig. 1A, treatment with 20 ng/ml WNT3a significantly induced the expression of AXIN2 (conductin), a prototypical target of β-catenin signaling (Lustig et al., 2002), at both indicated time points, demonstrating successful activation of the pathway. Correspondingly, siRNA-mediated knockdown of β-catenin by β-catenin–targeting small interfering RNA (siCAT) resulted in approximately 50% decreased mRNA expression not only of CTNNB1 mRNA (CTNNB1 is the name of the gene encoding β-catenin) but also of its downstream target gene, AXIN2, as assessed 72 hours after siRNA transfection (Fig. 1B). Measurement of corresponding protein levels of β-catenin revealed more than 60% of protein loss 72 hours and 96 hours after siRNA transfection compared with the nontargeting control siRNA, as assessed using densitometric analysis of the bands (Fig. 1B, bottom right).

Validation of WNT3a-mediated activation and siRNA-mediated inhibition of WNT/β-catenin activity. (A) Activation of WNT/β-catenin signaling pathways in HepaRG cells by treatment with WNT3a for 24 and 72 hours, as monitored by mRNA levels of the model β-catenin target gene AXIN2. (B) Specific knockdown of the β-catenin transcript in HepaRG cells using specific siCAT. Levels of mRNA expression of the β-catenin gene, CTNNB1, and of its downstream model target gene, AXIN2, were determined by using specific TaqMan assays compared with nontargeting siRNA (siCTR). Mean values of at least three independent experiments ± S.D. are depicted. Statistical significance (P < 0.05) is indicated by asterisks. rel., relative.
Effect of WNT/β-Catenin Pathway Modulation on Basal P450 Expression.
Changes in mRNA expression of major P450 enzymes upon WNT/β-catenin pathway stimulation or β-catenin knockdown were investigated in three independent cultures of HepaRG cells (Fig. 2). Interestingly, expression levels of only two genes, CYP2E1 (2.6-fold) and CYP1A1 (1.8-fold), were significantly upregulated 72 hours after activation of β-catenin, confirming previous observations (Gerbal-Chaloin et al., 2014). Accordingly, CYP2E1, but not CYP1A1, was more than 50% downregulated by siCAT, suggesting involvement of β-catenin especially in the basal regulation of CYP2E1. By contrast, basal expression of CYP2B6 was significantly increased upon knockdown of β-catenin. However, the observed minor decrease in CYP2B6 expression upon WNT3a treatment was not statistically significant.

Effect of WNT/β-catenin signaling on basal P450 expression. For β-catenin activation, HepaRG cells were treated for 72 hours with 20 ng/ml WNT3a (white bars) or with siRNA directed against β-catenin (dotted bars). The levels of the indicated P450s were measured by quantitative real-time polymerase chain reaction. Vehicle controls (0.1% bovine serum albumin and nontargeting control siRNA, respectively) were set to 1. Data represent means ± S.D. of three independent experiments measured in triplicate. Statistical significance (P < 0.05, paired t test) is indicated by asterisks. rel., relative.
Activation of WNT/β-Catenin Pathway Increases AhR-, CAR-, and PXR- Mediated Induction but Inhibits PPARα-Mediated Induction of P450 Genes.
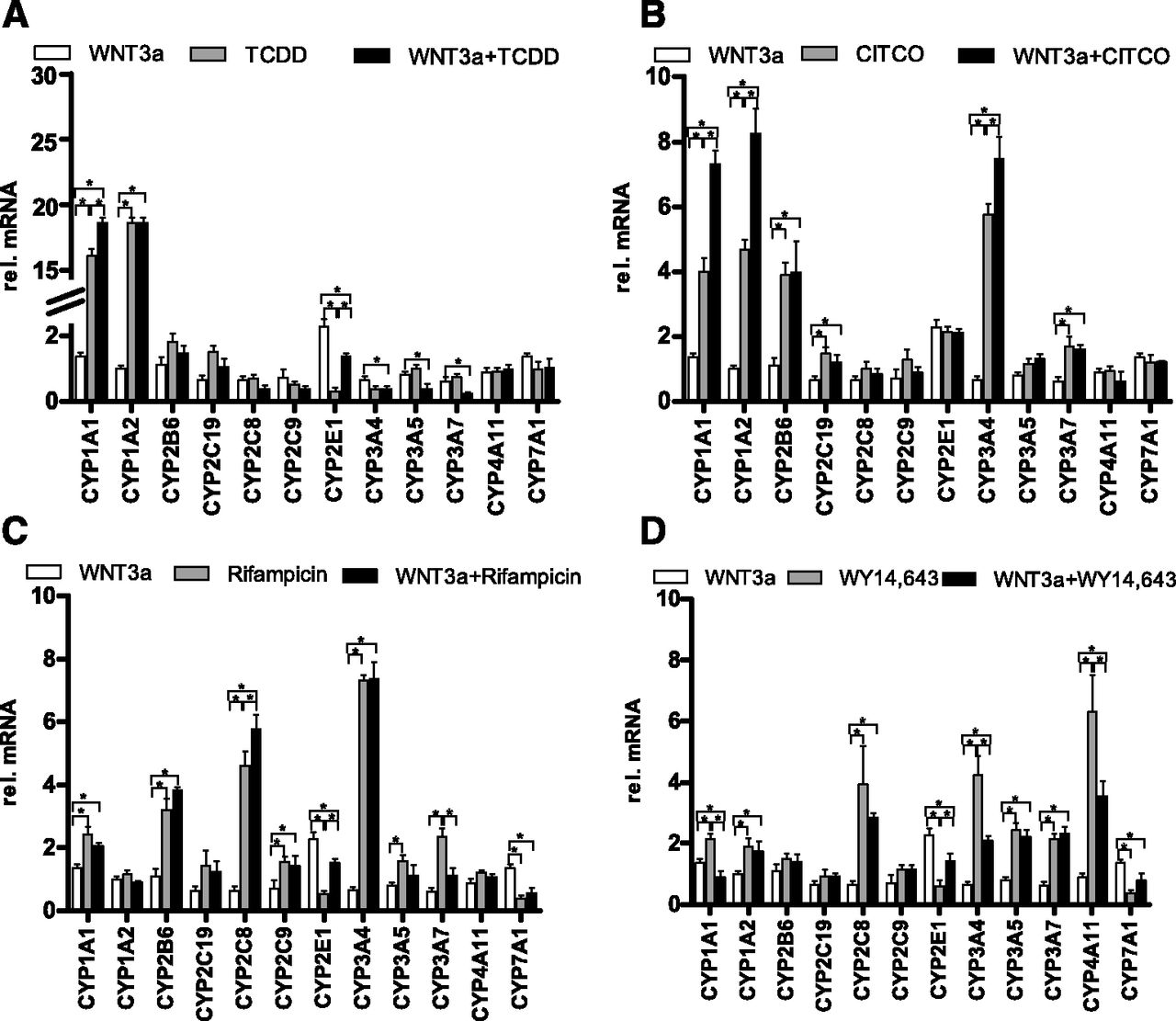
We next asked whether the WNT/β-catenin pathway interferes with the inducibility of P450 enzymes via nuclear receptors during ligand-mediated activation by their known agonists. HepaRG cells were treated with the indicated agonists of AhR (Fig. 3A), CAR (Fig. 3B), PXR (Fig. 3C), and PPARα (Fig. 3D) in combination with WNT3a treatment. Activation of the AhR with TCDD alone resulted in a more than 4-fold activation of the AhR-target genes CYP1A1 and CYP1A2 (Fig. 3A, light-gray bars). Interestingly, paralleled activation of the WNT/β-catenin pathway led to a significantly higher increase of CYP1A1, but not of CYP1A2, expression (Fig. 3A, black bars). In addition, activation of the AhR together with β-catenin restored expression of CYP2E1, which was significantly downregulated by TCDD alone. In addition to CYP1A genes, activation of CAR and PXR also resulted in the induction of CYP2B6, CYP2C19, CYP3A4, and CYP3A7 gene expression (Fig. 3, B and C, light-gray bars). Induction of WNT/β-catenin signaling in combination with CAR-ligand treatment resulted in synergistic increases of CYP1A1 and CYP1A2 and in a moderate increase of CYP3A4 (Fig. 3B, black bars).The expression of the same genes, together with CYP2C8, was similarly additionally increased after activation of PXR (Fig. 3C, black bars). Surprisingly, cotreatment of HepaRG cells with WNT3a and the PPARα agonist WY14,643 antagonized the WY14,643-mediated upregulation of CYP1A1, CYP1A2, CYP2C8, CYP3A4, and CYP4A11 gene expression (Fig. 3D, light-gray and black bars). Of note, ligand-mediated modulation of P450 gene expression in combination with WNT/β-catenin pathway activation was correspondingly reflected in the changes of their enzyme activity, as assessed by mass spectrometry–based analyses of the metabolism of a cocktail of specific substrates for the major P450s (Supplemental Fig. 1). No effects of WNT3a treatment on CYP7A1 expression were observed, whereas downregulation of CYP7A1 mRNA by PXR or PPARα activation was in line with previously published reports (Marrapodi and Chiang, 2000; Li and Chiang, 2005).

Expression profiling of P450s during ligand-activated induction of nuclear receptors in combination with WNT pathway activation. Expression levels of indicated P450s were analyzed by real-time quantitative reverse-transcription polymerase chain reaction after treatment of HepaRG cells for 72 hours with 20 ng/ml WNT3a alone (white bars in every panel) or with 10 nM of the AhR agonist TCDD alone (A), 5 µM of the CAR agonist CITCO (B), 10 µM of the PXR agonist rifampicin (C), or 100 µM of the PPARα agonist WY14,643 (D), as indicated for each treatment with light-gray bars in every panel. Combined treatment with WNT3a together with each agonist is shown with black bars in every panel. The bars indicate means ± S.D. relative to vehicle control set at 1. Asterisks indicate significant changes in the expression of target genes (P < 0.05, paired t test). rel., relative.
β-Catenin Is Required for P450 Induction via AhR, PXR, and CAR and Inhibits PPARα-Mediated Induction of P450 Gene Expression.
Activation of the WNT/β-catenin pathway via WNT3a triggers a downstream cascade, which includes post-translational protein modifications, cytoplasm-nucleus shuttling, and other secondary events that in part might not be directly related to the transcriptional activity of β-catenin. To further elucidate whether β-catenin itself interferes with the ligand-activated induction of P450 enzymes via nuclear receptors, we combined nuclear receptor agonist treatments with transfection of an siRNA (siCAT) targeting β-catenin. As shown in Fig. 4A, AhR-mediated induction of CYP1A genes was completely abolished after knockdown of β-catenin, demonstrating its requirement for TCDD-mediated upregulation. Similarly, loss of induction was also observed for CAR-mediated regulation of CYP1A1, CYP1A2, CYP2B6, and CYP3A4 (Fig. 4B) as well as PXR-mediated regulation of CYP1A1, CYP2B6, CYP2C8, and CYP3A4 (Fig. 4C). These results confirmed our previous observations using WNT3a treatment (Fig. 3, B and C) and further indicated an involvement of β-catenin in the inducibility of these genes via PXR and CAR (Fig. 3, B and C). By contrast, the combination of WY14,643 treatment with β-catenin knockdown resulted in the significantly higher induction of CYP1A1, CYP2C8, CYP2E1, and CYP3A genes (Fig. 4D), which is in line with the WNT3a-mediated effects on PPARα-mediated induction of these P450s (Fig. 3D). Interestingly, downregulation of β-catenin did not lead to an increased inducibility of CYP4A11 by PPARα, presumably due to already maximally induced status. Assessment of P450 enzyme activity after treatments with the agonists alone or in the combination with β-catenin knockdown confirmed the results of gene expression alterations (Supplemental Fig. 2).
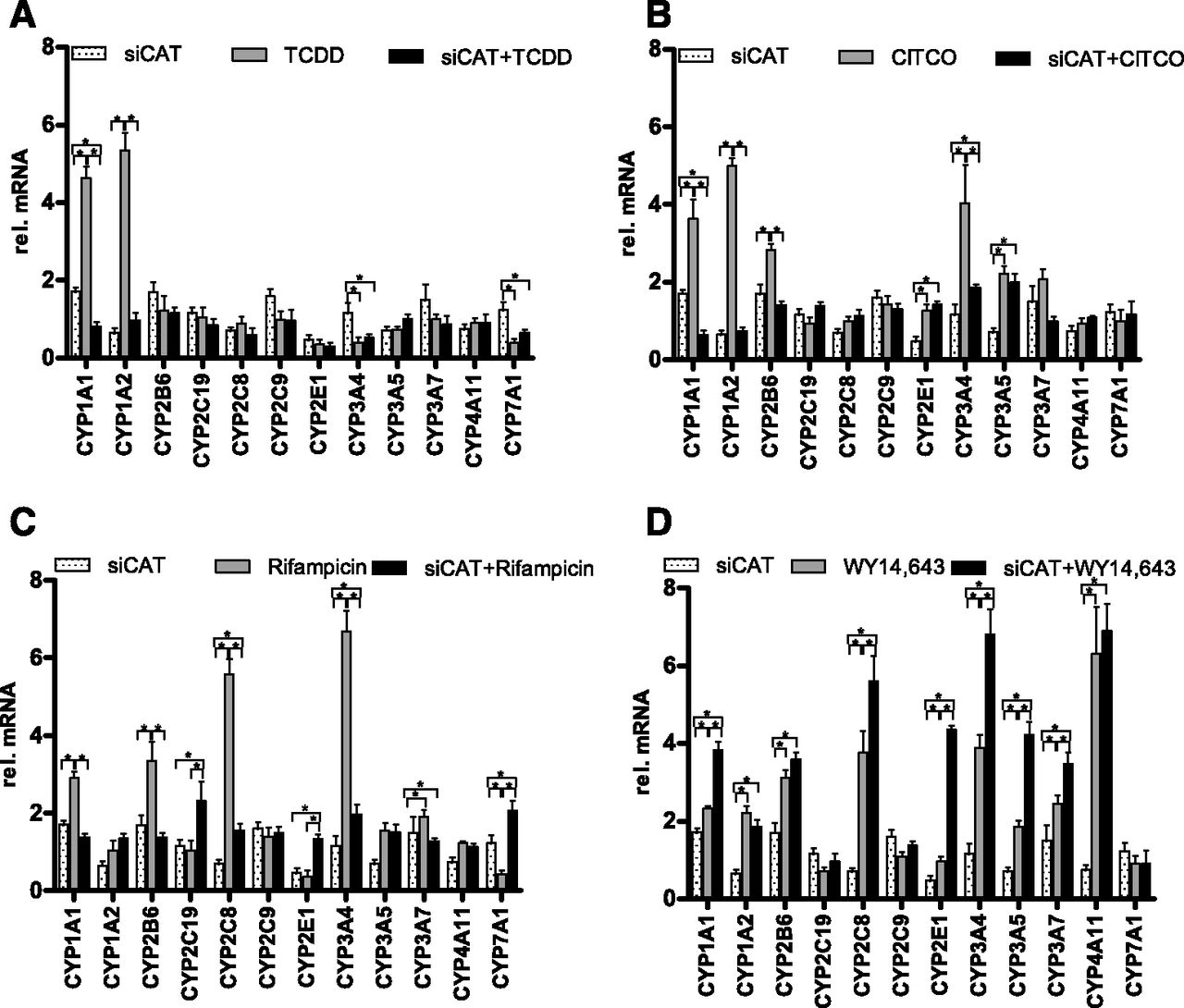
Expression profiling of P450 enzymes during ligand-activated induction of nuclear receptors in combination with β-catenin knockdown. Expression levels of indicated P450s were analyzed in HepaRG cells by real-time quantitative reverse-transcription polymerase chain reaction after β-catenin knockdown alone (white bars in every panel, siCAT), or with 10 nM of the AhR agonist TCDD alone (A), 5 µM of the CAR agonist CITCO (B), 10 µM of the PXR agonist rifampicin (C), or 100 µM of the PPARα agonist WY14,643 (D), as indicated for each treatment with light-gray bars in every panel. Combined treatment of β-catenin siRNA together with each agonist is shown with black bars in every panel. The bars indicate means ± S.D. relative to noncoding siRNA (siCTR) set at 1. Asterisks indicate significant changes in the expression of target genes (P < 0.05, paired t test). rel., relative.
Activating and Inhibitory Roles of β-Catenin in the Ligand-Mediated Induction of CYP3A4 by PXR and PPARα.
The crosstalk between the PPARα and WNT/β-catenin pathways has not been previously studied in human hepatocytes. We thus further focused on our novel observation of the opposite influence of β-catenin on the induction of P450 enzymes by PPARα versus other receptors. We thus analyzed protein expression of CYP3A4 by Western blotting after rifampicin-mediated activation of PXR, a well characterized regulator of CYP3A4 expression, and compared this with CYP3A4 protein expression under PPARα activation by WY14,643, either in the presence of WNT3a (Fig. 5A) or siCAT (Fig. 5B). In line with the mRNA expression profiles, coinduction of PPARα with β-catenin resulted in decreased CYP3A4 protein expression compared with WY14,643 treatment alone (Fig. 5A, gray bars). Activation of PXR together with β-catenin, however, led to an increased fold induction of CYP3A4 protein compared with the effect of rifampicin alone, confirming the effects at the mRNA level (Fig. 5A, black bars). Correspondingly, knockdown of β-catenin led to increased CYP3A4 protein expression by activated PPARα (Fig. 5B, gray bars) and to more than 50% reduced levels of CYP3A4 protein after treatment with rifampicin (Fig. 5B, black bars). We could further confirm the opposite roles of β-catenin in the ligand-mediated CYP3A4 induction by PXR and PPARα using a luciferase reporter construct containing 8.8 kb of an adjacent CYP3A4 promoter transfected in HuH7 cells (Fig. 5, C and D). Thus, we could show that β-catenin increased PXR-mediated induction but inhibited PPARα-mediated induction of CYP3A4 gene and protein expression.
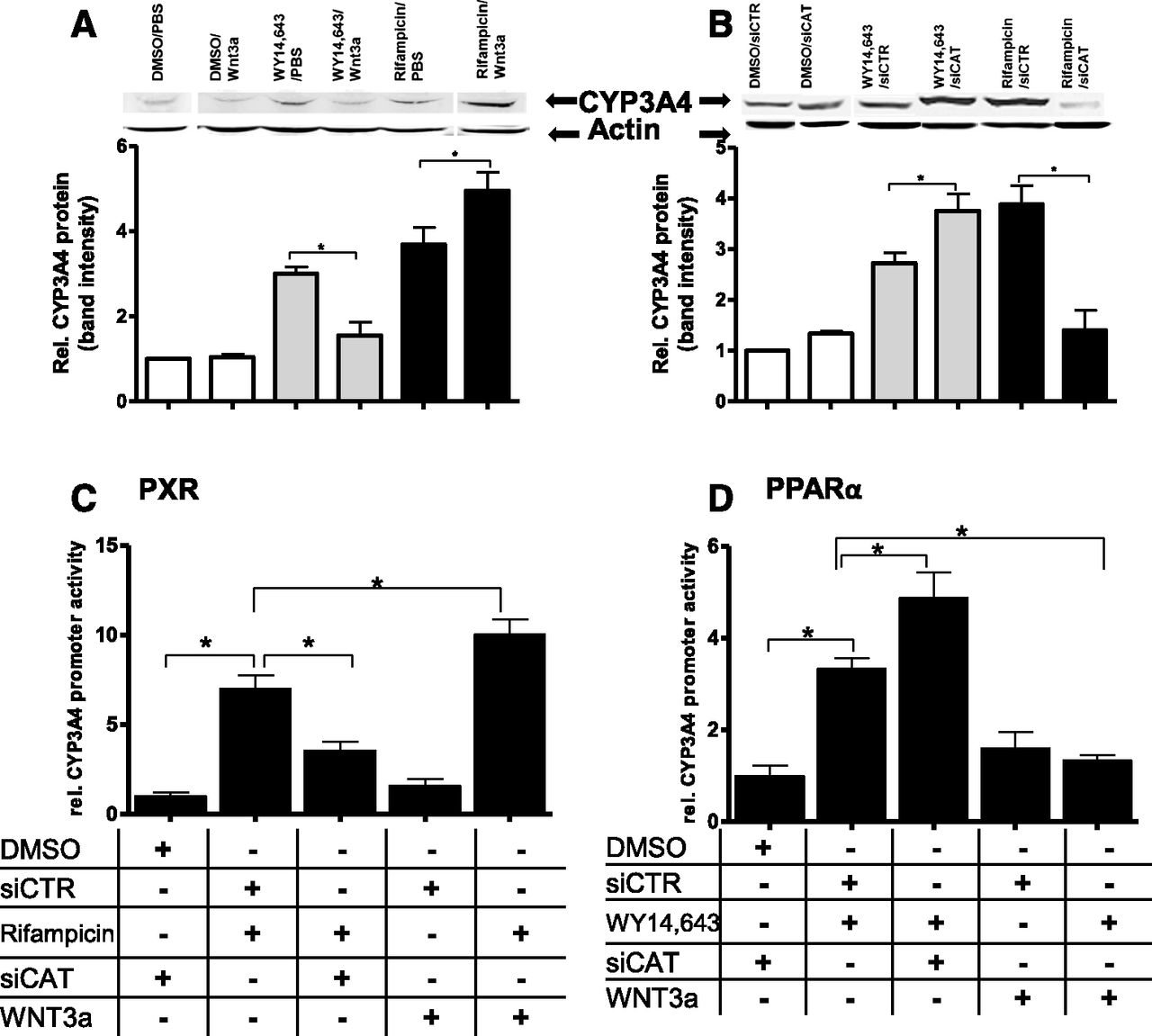
Activating and inhibitory roles of β-catenin in the ligand-mediated induction of CYP3A4 by PXR and PPARα. Protein expression levels of CYP3A4 were analyzed by Western blotting after activation of PPARα with 100 µM of WY14,643 (gray bars in both panels) or 10 µM of the PXR agonist rifampicin (black bars in both panels), in the presence of WNT3a (A) or siCAT (B). Western blot analysis with a previously characterized antibody against CYP3A4 (Wolbold et al., 2003) was performed in three biologic replicates; one representative is shown. Corresponding densitometric analyses of three replicates are shown under each blot, relative to PBS control (A) or noncoding siRNA (siCTR) (B), set to 1. The error bars indicate standard deviations between three independent experiments. Statistical significance is indicated by asterisks (P < 0.05). For further analyses at the level of gene transcription, a luciferase reporter gene construct containing 8.8 kb of the CYP3A4 promoter was transfected into HuH7 cells. Cells were treated with 10 µM rifampicin (C) or 100 µM WY14,643 (D) for 48 hours (vehicle control: DMSO), in the presence or absence of nontargeting siRNA (siCTR), siCAT, or WNT3a. Firefly luciferase activities were normalized to Renilla luciferase activities. Data are means ± S.D. of three independent experiments, each performed in triplicate. Statistical significance is indicated by asterisks (P < 0.05). DMSO, dimethylsulfoxide; PBS, phosphate-buffered saline. rel., relative.
Discussion
Although the involvement of the WNT/β-catenin pathway in the regulation of several P450 enzymes has been well characterized and described for mice, less evidence exists thus far for β-catenin–mediated modulation of P450 gene expression in humans. One previous study links the expression of some P450 isoforms in hepatoblastomas, pediatric human liver tumors, to the mutational activation of β-catenin signaling (Schmidt et al., 2011). In another very recent publication, a group of authors from France demonstrated the regulation of basal CYP2E1 and CYP1A2 expression by β-catenin in PHHs (Gerbal-Chaloin et al., 2014). Thus, we were interested in providing a systematic analysis of P450 gene expression modulation by β-catenin in HepaRG cells, not only at the basal level but also including P450 induction via nuclear receptor agonists. The latter phenomenon has, to the best of our knowledge, not been previously analyzed in human cells. HepaRG cells were chosen because of their high relevance as an in vitro model for human hepatocytes, which show less interindividual variability compared with cultures of PHHs (Klein et al., 2015). By two different, complementary approaches for modulating the WNT pathway (namely by using the canonical ligand WNT3a as well as by specific siRNA-mediated β-catenin downregulation), our data provide evidence for an intricate crosstalk between the WNT/β-catenin pathway and various receptors involved in the regulation of P450 gene expression. In particular, we could demonstrate that β-catenin is required for the full induction of a number of downstream P450 genes by AhR, PXR, and CAR. By contrast, β-catenin prevents the induction of CYP1A1, CYP2C8, CYP3A4, and CYP4A11 by PPARα. We could further show that the differential modulation of PXR- or PPARα-mediated CYP3A4 induction by β-catenin occurs within an 8.8-kb upstream region of the CYP3A4 promoter.
Our data on the influence of β-catenin on basal P450 expression are in accordance with a recent study, which demonstrated basal regulation of CYP1A1 and CYP2E1 by WNT/β-catenin (Gerbal-Chaloin et al., 2014). A similar connection between β-catenin and the murine Cyp2e1 (Braeuning et al., 2009; Gougelet et al., 2014) and Cyp1a1/Cyp1a2 mRNAs (Braeuning et al., 2009, 2011b; Vaas et al., 2014) has also been reported. Interestingly, the basal level of the mouse homolog of the human CYP2B6 gene, Cyp2b10, was upregulated in Ctnnb1 knockout mice (Braeuning et al., 2009; Ganzenberg et al., 2013), thereby confirming our findings on increased basal expression of CYP2B6 after siRNA-mediated β-catenin knockdown. The lack of major effects of β-catenin on the expression of CYP3A enzymes is also in line with previous results (Gerbal-Chaloin et al., 2014).
Synergistic activation of target gene expression by β-catenin and the nuclear receptors AhR and CAR has been previously studied in mice as well as in murine cell lines (Braeuning et al., 2009, 2011a; Ganzenberg et al., 2013; Vaas et al., 2014). Data presented in this article show, for the first time, synergistic effects of the WNT/β-catenin pathway and the receptors AhR, CAR, and PXR in human liver cells. By showing a synergism of AhR or CAR with β-catenin, our data are in line with previous data obtained in murine liver cells. Cooperative effects of β-catenin and PXR, as now observed in human liver cells, were not detectable in Ctnnb1 knockout mice treated with the PXR inducer pregnenolone-α-carbonitrile (Braeuning et al., 2009). This indicates that the regulation of some aspects of drug metabolism differ between rodent and human cells, thus necessitating studies with cells of human origin.
An important novel aspect presented in this study is the crosstalk between β-catenin and PPARα in the regulation of several P450s, but especially of CYP3A4, the most important P450 isoforms in humans. The role of PPARα in the regulation of drug-metabolizing enzymes from P450 families 1–3 has long been underestimated. However, there are a number of previous reports that demonstrate involvement of PPARα in the regulation of P450 enzymes mediating detoxification of exogenous compounds, including CYP3A4, not only in vitro (Schröder et al., 2011; Klein et al., 2012; Thomas et al., 2013, 2014) but also in vivo (de Keyser et al., 2013; Tsamandouras et al., 2014). Contrasting the situation with the other receptors, β-catenin antagonizes the effects of the activated PPARα on the expression of target genes. This effect occurs at the transcriptional level at the CYP3A4 promoter and is especially interesting, because the expression of the same gene is synergistically influenced by β-catenin and PXR.
Classically, active β-catenin binds to transcription factors of the T cell factor (TCF) family to initiate the transcription of target genes, acting as a transcriptional coactivator of the TCF proteins. Synergistic interactions between binding sites for TCF/β-catenin and binding sites for nuclear receptors (e.g., by easier access of the ligand-activated nuclear receptor to a gene promoter that has already bound TCF/β-catenin and therefore changed its chromatin structure) may play a role in the observed interactions between β-catenin and the receptors AhR, CAR, and PXR. Of note, an interaction between a TCF/β-catenin binding site and an AhR binding site in the human CYP1A1 promoter has been reported (Braeuning et al., 2011b). Thus far, no unequivocal consensus TCF binding sites were identified within the CYP3A4 promoter, which means that other possible mechanisms should be taken into account. Activatory effects of β-catenin on certain genes might also be mediated by its interaction with other transcription factors. For example, it was published that β-catenin physically interacts with the AhR and modulates the activity of the ligand-activated AhR at its DNA binding sites (Braeuning et al., 2011b). This effect is thought to be exerted either by β-catenin itself, acting as a transcriptional coactivator of the AhR, or by effects of β-catenin on the recruitment of other proteins involved in the transcription process. The AhR is also a direct transcriptional target of β-catenin in human cells (Chesire et al., 2004; Gerbal-Chaloin et al., 2014). Moreover, recent data point toward a crosstalk of β-catenin and hepatocyte nuclear factor 4α in the regulation of expression of liver-specific genes in vitro and in vivo, suggesting a further possible mechanism of P450 modulation (Colletti et al., 2009; Gougelet et al., 2014). Hepatocyte nuclear factor 4α plays an important role in the regulation of various P450s that are also regulated by different xenosensing nuclear receptors.
It is more difficult to find mechanistic explanations for the observed inhibitory action of β-catenin on PPARα-dependent effects. An interference via glycogen synthase 3β (GSK) seems possible, because GSK, a key component in canonical WNT signaling, might be also involved in the regulation of PPARα (Pawlak et al., 2015). However, this scenario would only apply to effects of WNT3a, not for siRNA-mediated silencing of β-catenin, since the latter approach interferes with the WNT pathway downstream of GSK. Moreover, the extent to which the overall activity of GSK is modulated by WNT signaling is not known, since a major part of the regulation during WNT3a-mediated activation of the pathway seems to be related to the dissociation of the GSK-containing multiprotein complex, rather than to direct inhibition of the GSK enzyme. The interaction between β-catenin and the AhR has been studied in detail and therefore might provide hints for mechanisms possibly also relevant for the interaction of β-catenin with PPARα: β-catenin acts as a ubiquitin ligase for the AhR in murine intestinal cells (Kawajiri et al., 2009). Taken together, these findings indicate that β-catenin can exert much more diverse cellular effects than dimerization with TCF proteins. Further studies will reveal in-depth mechanistic details of the novel β-catenin/PPARα interaction. In summary, our findings show that β-catenin is an important modulator of basal as well as nuclear receptor-mediated P450 expression triggered by xenobiotics in human liver cells. The identification of the antagonistic connection of β-catenin and PPARα adds a new layer of crosstalk to the complex network of β-catenin and nuclear receptors and substantiates the role of the WNT/β-catenin signaling pathway as a master regulator of drug and xenobiotic metabolism in the human liver.
Acknowledgments
The authors thank Igor Liebermann, Britta Klumpp, and Kyoko Momoi (Dr. Margarete Fischer-Bosch Institute of Clinical Pharmacology, Stuttgart, Germany) for expert technical assistance, as well as Christophe Chesne and Biopredic for help and support with HepaRG cells.
Authorship Contributions
Participated in research design: Thomas, Bayha, Vetter, Braeuning.
Conducted experiments: Thomas, Bayha, Vetter, Hofmann, Braeuning.
Performed data analysis: Thomas, Bayha, Vetter, Hofmann, Braeuning.
Wrote or contributed to the writing of the manuscript: Thomas, Schwarz, Zanger, Braeuning.
Footnotes
- Received December 12, 2014.
- Accepted March 30, 2015.
This research was supported by the German Federal Ministry of Education and Research Virtual Liver Network [Grant 0315755] and the Robert Bosch Foundation, Stuttgart, Germany.
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- AhR
- aryl hydrocarbon receptor
- CAR
- constitutive androstane receptor
- CITCO
- 6-(4-chlorophenyl)imidazo[2,1-b][1,3]thiazole-5-carbaldehyde-O-(3,4-dichlorobenzyl)oxime
- GSK
- glycogen synthase 3β
- P450
- cytochrome P450
- PHH
- primary human hepatocyte
- PPAR
- peroxisome proliferator–activated receptor
- PXR
- pregnane X receptor
- siCAT
- β-catenin–targeting small interfering RNA
- siRNA
- small interfering RNA
- TCDD
- 2,3,7,8-tetrachlorodibenzo-p-dioxin
- TCF
- T cell factor
- WY14,643
- 4-chloro-6-(2,3-xylidino)-2-pyrimidinylthioacetic acid
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}