


Abstract
Azelastine, an antiallergy and antiasthmatic drug, has been reported to be mainly N-demethylated to desmethylazelastine in humans. In the present study, Eadie-Hofstee plots of azelastine N-demethylation in human liver microsomes were biphasic. In microsomes from human B-lymphoblast cells, recombinant cytochrome P-450 (CYP)2D6 and CYP1A1 exhibited higher azelastine N-demethylase activity than did other CYP enzymes. On the other hand, recombinant CYP3A4 and CYP1A2 as well as CYP1A1 and CYP2D6 in microsomes from baculovirus-infected insect cells were active in azelastine N-demethylation. TheKM value of the recombinant CYP2D6 (2.1 μM) from baculovirus-infected insect cells was similar to theKM value of the high-affinity (2.4 ± 1.3 μM) component in human liver microsomes. On the other hand, theKM values of the recombinant CYP3A4 (51.1 μM) and CYP1A2 (125.4 μM) from baculovirus-infected insect cells were similar to the KM value of the low-affinity (79.7 ± 12.8 μM) component in human liver microsomes. Bufuralol inhibited the high-affinity component, making the Eadie-Hofstee plot in human liver microsomes monophasic. AzelastineN-demethylase activity in human liver microsomes (5 μM azelastine) was inhibited by ketoconazole, erythromycin, and fluvoxamine (IC50 = 0.08, 18.2, and 17.2 μM, respectively). Azelastine N-demethylase activity in microsomes from twelve human livers was significantly correlated with testosterone 6β-hydroxylase activity (r = 0.849,p < .0005). The percent contributions of CYP1A2, CYP2D6, and CYP3A4 in human livers were predicted using several approaches based on the concept of correction with CYP contents or relative activity factors (RAFs). Our data suggested that the approach using RAFCL (RAF as the ratio of clearance) is most predictive of the N-demethylation clearance of azelastine because it best reflects the observedN-demethylation clearance in human liver microsomes. Summarizing the results, azelastine N-demethylation in humans liver microsomes is catalyzed mainly by CYP3A4 and CYP2D6, and CYP1A2 to a small extent (in average, 76.6, 21.8, and 3.9%, respectively), although the percent contribution of each isoform varied among individuals.
Cytochrome P-450 (CYP)1 comprises a superfamily of enzymes that have long been recognized as the primary enzymes responsible for human drug metabolism. Although the number of individual enzymes that have been identified and characterized is increasing (Nelson et al., 1996), the metabolism of xenobiotics in humans is handled mainly by enzymes from three families: CYP1, CYP2, and CYP3 (Spatzenegger and Jaeger, 1995). Drug interactions can cause severe complications from medications. Clinically relevant drug interactions are often the result of the effects on CYP enzymes involved in biotransformation (Muck, 1994).
Azelastine is a long-acting antiallergy and antiasthmatic drug. Azelastine possesses properties beyond histamine H1 receptor-blocking activity. These include antagonism of the chemical mediators adenosine, LTC4, LTD4, endothelin-1, and platelet activation factor, and inhibition of the generation and/or release of histamine, interleukin-1β, leukotrienes, and superoxide free radicals (Perhach et al., 1989; Szelenyi, 1989). Azelastine may also prevent Ca2+-mediated cellular events through a reversible inhibition of inward current and the voltage-dependent L-type Ca2+ channels (Hazama et al., 1994). Azelastine was reported to be metabolized byN-demethylation to desmethylazelastine (Fig.1) in mammals (Tatsumi et al., 1984). It has also been established that desmethylazelastine has pharmacologic activity equivalent to the parent drug (Perhach et al., 1989; Szelenyi, 1989). It has been reported that azelastine and desmethylazelastine inhibited the CYP2C19 and CYP2D6 activities in human liver microsomes (Morganroth et al., 1997). However, the CYP enzymes responsible for azelastine N-demethylation in human livers are unknown. Therefore, in the present study, we identified the CYP enzymes involved in azelastine N-demethylation in human liver microsomes to evaluate the possibility of drug interactions with respect to this metabolic pathway.

Azelastine N-demethylation pathway in humans.
Recently, several prediction methods for assessing the contribution of multiple CYPs to certain metabolic reactions in human liver microsomes have been reported (Crespi, 1995; Iwatsubo et al., 1997; Becquemont et al., 1998). We determined the percent contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation in human liver microsomes using recombinant CYPs expressed in human B-lymphoblast cells or baculovirus-infected insect cells based on these prediction methods. This is the first report in which these prediction methods, including relative activity factor (RAF), were evaluated in terms of different expression systems to estimate the contribution of each CYP in certain types of drug metabolism.
Materials and Methods
Chemicals.
Azelastine hydrochloride, 4-(p-chlorobenzyl)-2-(hexahydro-1-methyl-1H-azepin-4-yl)-1(2H)-phthalazinone hydrochloride, was provided by Eisai (Tokyo, Japan). Desmethylazelastine hydrobromide, 4-(p-chlorobenzyl)-2-(hexahydro-1H-azepin-4-yl)-1(2H)-phthalazinone hydrobromide, was provided by Degussa Japan (Tokyo, Japan). NADP+, glucose 6-phosphate, and glucose 6-phosphate dehydrogenase were purchased from Oriental Yeast (Tokyo, Japan). (±)-Bufuralol hydrochloride, 1′-hydroxybufuralol maleate, sulfaphenazole, S-mephenytoin, and ketoconazole were obtained from Ultrafine chemicals (Manchester, UK). 7-Ethoxyresorufin and resorufin were purchased from Sigma (St. Louis, MO). Testosterone, 6β-hydroxytestosterone, and 11β-hydroxytestosterone were obtained from Steraloids (Wilton, NH). Furafylline and fluvoxamine maleate were obtained from Funakoshi (Tokyo, Japan) and Tocris Cookson (Ballwin, MO), respectively. Other chemicals were of the highest grade commercially available.
Enzyme Preparations.
Microsomes from human B-lymphoblast cells expressing CYP1A1, CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9(Arg), CYP2C19, CYP2D6(Val), CYP2E1, and CYP3A4 were obtained from Gentest (Woburn, MA). These were coexpressed with NADPH-CYP oxidoreductase (OR), except for CYP1A2, CYP2B6, or CYP2C19. Microsomes from baculovirus-infected insect cells expressing CYP1A1, CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9(Arg), CYP2C19, CYP2D6(Val), CYP2E1, CYP3A4, and CYP3A5 were also obtained from Gentest. All enzymes were coexpressed with OR. CYP2E1 or CYP3A4 were also coexpressed with cytochrome b5(b5).
Human liver samples (HL 1–HL 5) were obtained from autopsy. The use of human livers for this study was approved by the Institutional Committee of Tokyo Medical Examiner's Office, Japan. Liver tissues were rapidly frozen in liquid nitrogen immediately after excision and were stored at −80°C. Microsomes from human liver were prepared as described previously (Kamataki and Kitagawa, 1974) and were stored at −80°C until use. The protein concentrations were measured according to the method of Lowry et al. (1951). The CYP content was estimated by the method of Johannesen and Depierre (1978). Human liver microsomes (HLG1–HLG12) and pooled human liver microsomes (lot #2, comprised of 20% each of specimens HLG1, HLG3, HLG13, HLG14, and 10% each of specimens HLG4 and HLG6) were also purchased from Gentest. The immunochemically determined CYP contents except for CYP2C and specific catalytic activities of each CYP isoform in those microsomes were provided in the data sheets by the manufacturer.
Azelastine N-Demethylase Activity.
The azelastine N-demethylase activities in human liver microsomes or microsomes from two different expression systems were determined by HPLC. A typical incubation mixture (0.2 ml of total volume) contained 100 mM potassium phosphate buffer (pH 7.4), an NADPH-generating system (0.5 mM NADP+, 5 mM glucose 6-phosphate, 5 mM MgCl2, 1 U/ml glucose 6-phosphate dehydrogenase), azelastine, and 0.5 mg/ml microsomal protein of human livers. The reaction was initiated by the addition of the NADPH-generating system after a 2-min preincubation at 37°C. The reaction mixtures were incubated for 30 min and reactions were terminated by the addition of 0.1 ml of ice-cold acetonitrile. After removal of the protein by centrifugation at 10,000 rpm for 5 min, a 100-μl portion of the supernatant was subjected to HPLC. For recombinant CYPs, the incubation mixture was of the same composition as mentioned above, except for 1.0 mg/ml of microsomal protein for microsomes from B-lymphoblast cells and 10 pmol/ml of CYP for microsomes from baculovirus-infected insect cells. The azelastine concentration used was 5 μM. The mixture was incubated for 60 or 30 min at 37°C for microsomes from B-lymphoblast cells or baculovirus-infected insect cells, respectively.
HPLC analyses were performed according to the method of Pivonka et al. (1987) with slight modifications. The HPLC equipment was a L-6000 pump (Hitachi, Tokyo, Japan), 712 WISP intelligent sample processor (Waters, Tokyo, Japan), Chromatopak C-R5A (Shimadzu, Kyoto, Japan), CTO-6A column oven (Shimadzu) with a Capcell Pak C18 UG120 (4.6 × 250 mm; 4 μm) column (Shiseido, Tokyo, Japan). The eluent was monitored fluorometrically (excitation, 213 nm; emission, 360 nm) using a F-1080 fluorescence detector (Hitachi). The mobile phase was 20% tetrahydrofuran containing 0.3% triethylamine (pH 2.4, adjusted with phosphoric acid). The flow rate was 1.0 ml/min and the column temperature was 35°C.
Kinetic Analysis.
The kinetic studies were performed using microsomes from five different human livers (HL 1–HL 5), from B-lymphoblast cells, or from baculovirus-infected insect cells expressing human CYP. In determining the kinetic parameters, the azelastine concentration ranged from 1 to 200 μM. Eadie-Hofstee plots were constructed to determine whether kinetics were mono- or biphasic. Kinetic parameters were estimated from the fitted curves using a computer program (KaleidaGraph; Synergy Software, Reading, PA) designed for nonlinear regression analysis. The following equations were applied for assuming one or two independent enzymatic activities:
Inhibition Analysis.
CYP specific inhibitors were screened for their effects on azelastineN-demethylation in pooled human liver microsomes at an azelastine concentration of 5 μM. The inhibitors studied were fluvoxamine (Pastrakuljic et al., 1997), furafylline (Tassaneeyakul et al., 1994), coumarin (Yun et al., 1991), sulfaphenazole (Baldwin et al., 1995), S-mephenytoin (Chiba et al., 1993), quinidine (Broly et al., 1989), bufuralol (Yamazaki et al., 1994), chlorzoxazone (Peter et al., 1990), erythromycin (Watkins et al., 1985), and ketoconazole (Baldwin et al., 1995). The range of concentration used was 0.1 to 100 μM. With the exception of quinidine, which was dissolved in dimethyl sulfoxide, the inhibitors were dissolved in methanol such that the final concentration of solvent in the incubation mixture was <0.1%. The incubation mixture including chemical inhibitors was preincubated for 2 min before the reaction was initiated by the addition of an NADPH-generating system. AzelastineN-demethylation was determined as described above.
Ethoxyresorufin O-Deethylase Activity.
The activity in human liver microsomes was determined fluorometrically as described previously (Sinjari et al., 1993) with slight modifications. The incubation mixture (0.5 ml of total volume) contained 100 mM Tris-HCl buffer (pH 7.4), 1.6 mg/ml BSA, 0.25 mM NADPH, and 0.2 mg/ml microsomal protein. The reaction was initiated by the addition of 7-ethoxyresorufin (final concentration, 0.1–2 μM) dissolved in dimethyl sulfoxide after a 3-min preincubation at 37°C. After the incubation for 15 min at 37°C, the reaction was terminated by the addition of 2 ml of ice-cold methanol. The incubation mixtures were centrifuged at 1000g for 10 min, and the concentration of the resorufin formed in the supernatant was determined fluorometrically (excitation, 574 nm; emission, 596 nm) with an F-4500 fluorescence spectrophotometer (Hitachi).
Bufuralol 1′-Hydroxylase Activity.
The activity in human liver microsomes was determined as described previously (Kronbach et al., 1987) with slight modifications. The incubation mixture (0.2 ml of total volume) contained 50 mM potassium phosphate buffer (pH 7.4), an NADPH-generating system, 0.2 mg/ml microsomal protein, and 1 to 10 μM bufuralol as the substrate. The reaction was initiated by the addition of an NADPH-generating system after a 2-min preincubation at 37°C. After incubation for 10 min at 37°C, the reaction was terminated by the addition of 20 μl of 60% perchloric acid. The incubation mixtures were centrifuged at 10,000 rpm for 5 min, and a 20-μl portion of the supernatant was subjected to HPLC. The HPLC equipment was the same as described above. The analytical column was Capcell Pak C18 UG120 (4.6 × 250 mm, 4 μm, Shiseido) and the formed 1′-hydroxybufuralol was detected fluorometrically (excitation, 252 nm; emission, 302 nm). The mobile phase was 20% acetonitrile containing 1 mM perchloric acid. The flow rate was 1.0 ml/min and the column temperature was 35°C.
Testosterone 6β-Hydroxylase Activity.
The activity in human liver microsomes was determined as described previously (Arlotto et al., 1991) with slight modifications. The incubation mixture (0.2 ml of total volume) contained 100 mM potassium phosphate buffer (pH 7.4), an NADPH-generating system, 1.0 mg/ml microsomal protein, and 5 to 200 μM testosterone as the substrate. The reaction was initiated by the addition of an NADPH-generating system after a 2-min preincubation at 37°C. After incubation for 5 min at 37°C, the reaction was terminated by the addition of 0.1 ml of ice-cold acetonitrile. 11β-Hydroxytestosterone (50 ng) was added as an internal standard. The incubation mixtures were centrifuged at 10,000 rpm for 5 min and a 100-μl portion of the supernatant was subjected to HPLC. HPLC analyses were performed using an L-6200 intelligent pump (Hitachi), AS-8010 autosampler (Tosoh, Tokyo, Japan), D-2000 chromato-integrator (Hitachi), and 865-CO column oven (Jasco, Tokyo, Japan), equipped with a Capcell Pak C18 UG120 (4.6 × 250 mm; 4 μm) column (Shiseido). The eluent was monitored at 240 nm using an L-4200 UV/VIS detector (Hitachi). The mobile phase was solvent A (methanol/water/acetonitrile, 39:60:1) and solvent B (methanol/water/acetonitrile, 80:18:2). Typical conditions for elution were as follows: 25% B (0–25 min); 25 to 80% B (25–28 min); 80 to 25% B (28–33 min). A linear gradient was used for all solvent changes. The flow rate was 1.0 ml/min and the column temperature was 35°C.
Contributions of CYP1A2, CYP2D6, and CYP3A4 to AzelastineN-Demethylase Activity in Human Liver Microsomes.
The contributions of each CYP to the azelastineN-demethylase activity in human liver microsomes were estimated by five different prediction methods.
Prediction Method 1.
The percent contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastineN-demethylase activity (1 μM azelastine concentration) were estimated based on the contents of these CYP proteins in human liver microsomes determined by immunoblotting, based on the following equations (Becquemont et al., 1998):
Prediction Method 2.
The percent contributions of these CYPs to azelastineN-demethylase activity (1 μM azelastine) were estimated based on the kinetic parameters of recombinant CYP1A2, CYP2D6, and CYP3A4, and the contents of these CYP proteins in human liver microsomes estimated by immunoblotting (Yamazaki et al., 1997). The following equations were used for the prediction of the expected activities (Iwatsubo et al., 1997):
Prediction Method 3.
The percent contributions of these CYPs to azelastineN-demethylation were estimated as described previously (Nakajima et al., 1998) by applying RAF values as proposed by Crespi (1995) using the values of the activities (RAFv). The RAFv of CYP1A2 (RAFv, CYP1A2) was determined as the ratio of ethoxyresorufinO-deethylase activity (at a substrate concentration of 2 μM), a specific metabolic reaction mediated by CYP1A2 (Sinjari et al., 1993), in human liver microsomes to the activity for recombinant CYP1A2. The RAFv of CYP2D6 (RAFv, CYP2D6) was determined as the ratio of the activity of bufuralol 1′-hydroxylation (at a substrate concentration of 1 μM), a specific metabolic reaction mediated by CYP2D6 (Kronbach et al., 1987), in human liver microsomes to the activity for recombinant CYP2D6. The RAFv of CYP3A4 (RAFv, CYP3A4) was determined as the ratio of the activity of testosterone 6β-hydroxylation (at a substrate concentration of 100 μM), a specific metabolic reaction mediated by CYP3A4 (Waxman et al., 1988), in human liver microsomes to the activity for recombinant CYP3A4. Using RAFv, theN-demethylation clearance of azelastine by CYP1A2, CYP2D6, and CYP3A4 in human liver microsomes (CLCYP1A2, CLCYP2D6, and CLCYP3A4, respectively) were expressed as follows:
The contributions of CYP1A2, CYP2D6, and CYP3A4 to the clearance of azelastine N-demethylation by human liver microsomes were calculated using the following equations:
Prediction Method 4.
The percent contributions were estimated by the application of RAFs using Vmax values (RAFVmax), instead of RAFvin eqs. 12 to 14. The RAFVmax values of CYP1A2, CYP2D6, and CYP3A4 were determined as the ratio of theVmax of ethoxyresorufinO-deethylase activity, bufuralol 1′-hydroxylase activity, and testosterone 6β-hydroxylase activity, respectively, in human liver microsomes to the Vmax values for recombinant CYPs. The contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation by human liver microsomes were calculated using eqs. 15 to 17, respectively.
Prediction Method 5.
The percent contributions were estimated by the application of the RAF using CL values (RAFCL). The RAFCL values for CYP1A2, CYP2D6, and CYP3A4 were determined as the ratios of the CL of ethoxyresorufinO-deethylase activity, bufuralol 1′-hydroxylase activity, and testosterone 6β-hydroxylase activity, respectively, in human liver microsomes to the values for recombinant CYPs. The contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation by human liver microsomes were calculated using eqs. 15 to 17, respectively.
Statistical Analysis.
Correlations between the azelastine N-demethylase activity and ethoxyresorufin O-deethylase activity, bufuralol 1′-hydroxylase activity, or testosterone 6β-hydroxylase activity in microsomes from twelve human livers were determined using linear regression analyses. Results are expressed as means ± S.D.
Results
Azelastine N-Demethylation in Human Liver Microsomes.
Azelastine and desmethylazelastine were well separated from the endogenous components with HPLC (Fig. 2). Incubation of azelastine with human liver microsomes yielded only one peak of which the retention time was identical with that of authentic desmethylazelastine. The formation of desmethylazelastine at 10 μM azelastine increased linearly for up to 60 min of incubation time. Unless specified, an incubation time of 30 min was used to ensure initial rate conditions for the formation of desmethylazelastine. Figure 3 shows a typical Eadie-Hofstee plot for azelastine N-demethylase activity in human liver microsomes (HL 2). The plot indicated that multiple enzymes were responsible for the biotransformation of azelastine to desmethylazelastine. For the high-affinity component in microsomes from four different human livers, the mean values ± S.D. ofKM1 and Vmax1were 2.4 ± 1.3 μM and 13.2 ± 10.1 pmol/min/mg, respectively. For the low-affinity component, the mean values ± S.D. of KM2 andVmax2 were 79.7 ± 12.8 μM and 178.9 ± 54.3 pmol/min/mg, respectively (Table1).

Representative HPLC chromatograms of azelastine N-demethylation in human liver microsomes.
The standard mixture spiked with desmethylazelastine under the same conditions described for samples (A); the incubation mixture of human liver microsomes without (B) and with (C) azelastine. Peak: 1, desmethylazelastine; 2, azelastine.

Typical Eadie-Hofstee plot for azelastine N-demethylase activity in human liver microsomes.
Data are the mean of duplicate determinations (HL 2).
Kinetic parameters of azelastine N-demethylase activity in human liver microsomes
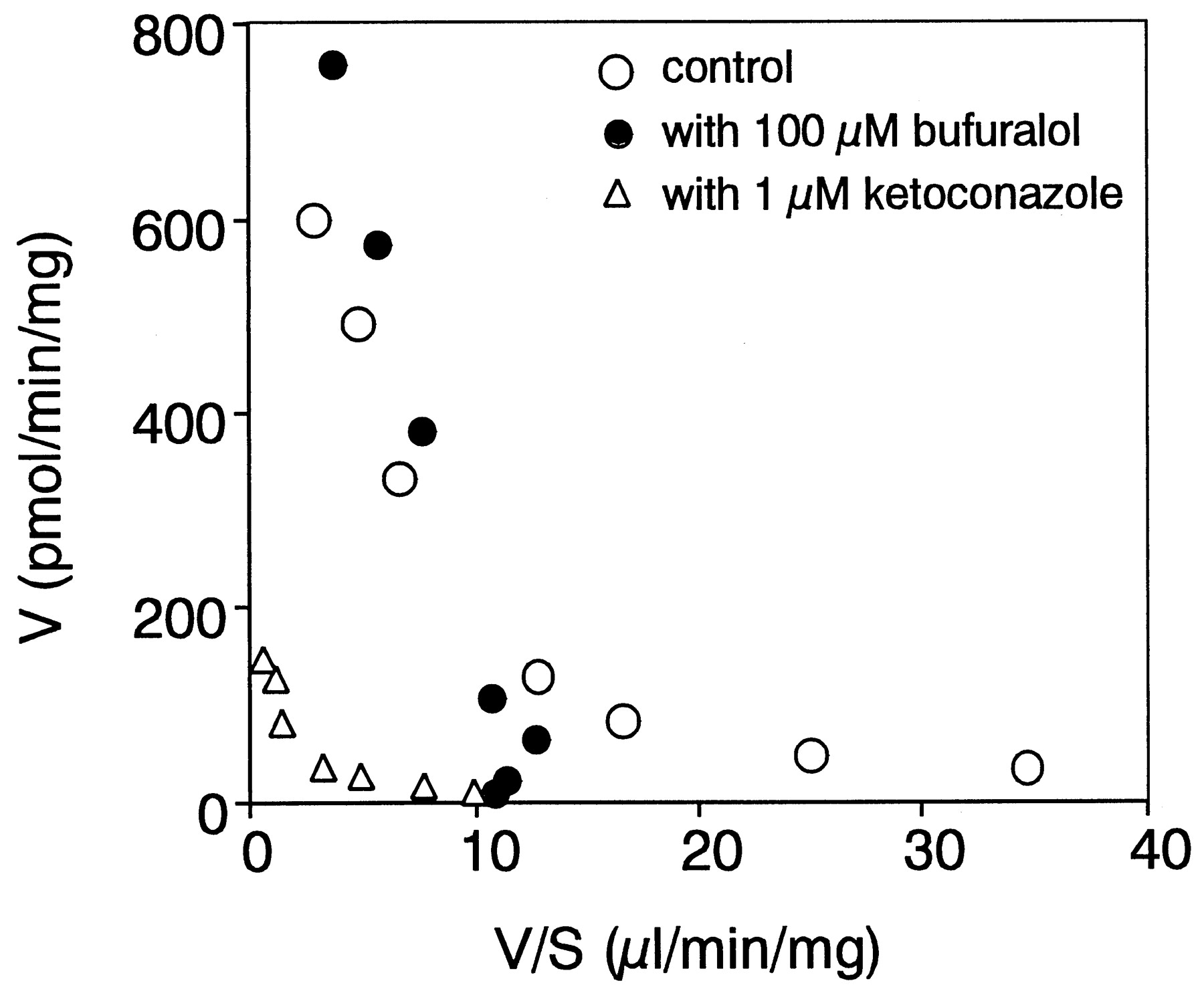
The kinetic parameters for azelastine N-demethylation in pooled human liver microsomes (Gentest, lot #2) were also determined (KM1 = 0.6 μM,Vmax1 = 40.4 pmol/min/mg,KM2 = 79.0 μM andVmax2 = 785.3 pmol/min/mg). As shown in Fig. 4, 100 μM bufuralol inhibited the high-affinity component in the human liver microsomes causing the Eadie-Hofstee plot to become monophasic (KM= 78.9 μM, Vmax = 975.9 pmol/min/mg). On the other hand, 1 μM ketoconazole inhibited both components, and the Eadie-Hofstee plot remained biphasic (KM1 = 0.8 μM, Vmax1 = 15.2 pmol/min/mg,KM2 = 81.8 μM andVmax2 = 188.3 pmol/min/mg).
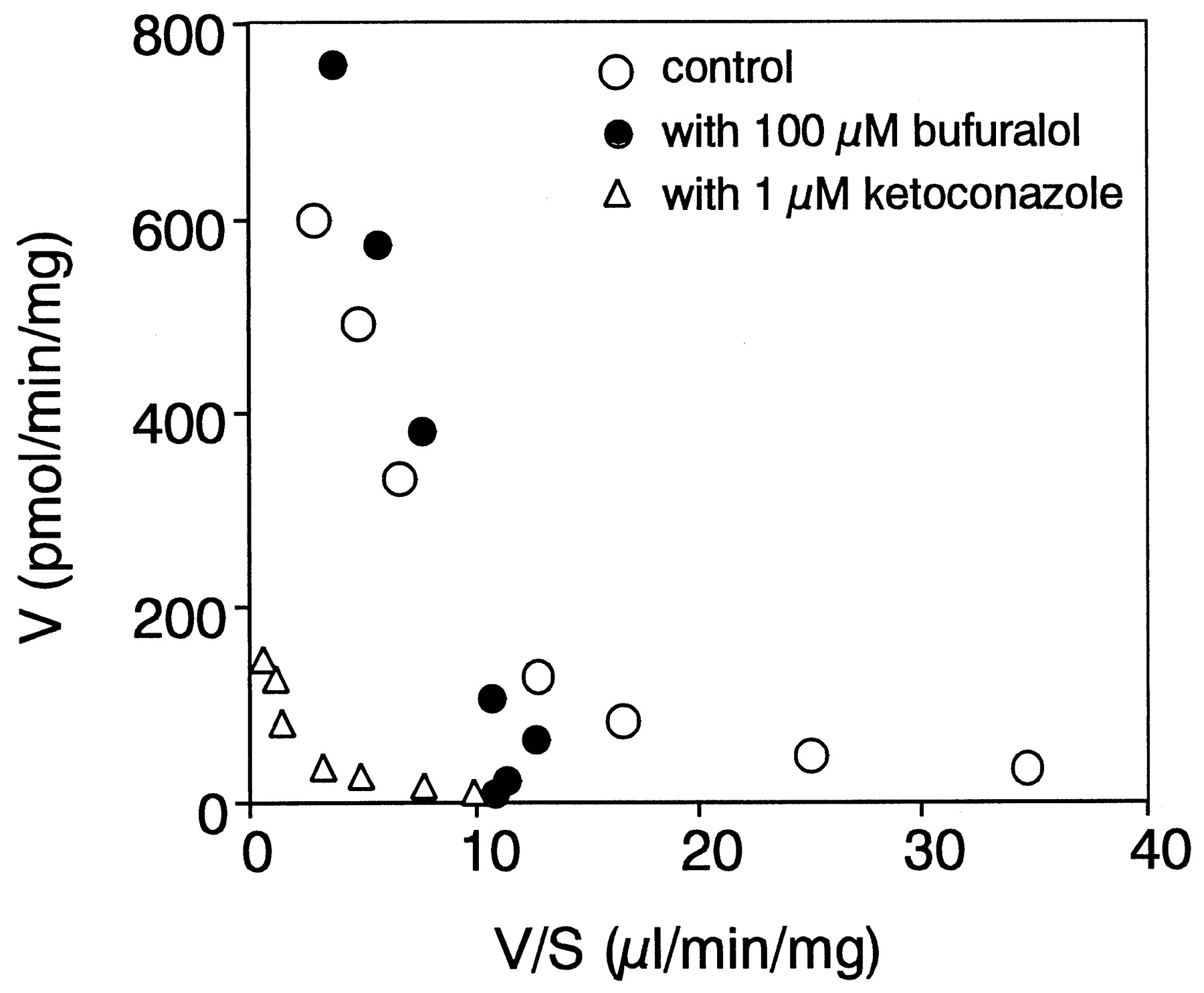
Eadie-Hofstee plot for azelastine N-demethylase activity in pooled human liver microsomes.
Azelastine N-demethylase activity in pooled human liver microsomes was determined with 100 μM bufuralol (●), 1 μM ketoconazole (▵), and without inhibitor (○) at 1 to 200 μM azelastine. Data are the mean of duplicate determinations.
Inhibition Analysis.
CYP-specific inhibitors were screened for their effects on azelastineN-demethylase activity in pooled human liver microsomes (Fig. 5) at an azelastine concentration of 5 μM (∼KM value of the high-affinity component). Ketoconazole, a specific inhibitor of CYP3A4, essentially abolished the conversion of azelastine to desmethylazelastine (IC50 = 0.08 μM). Erythromycin, a substrate of CYP3A4, also inhibited azelastine N-demethylation (IC50 = 18.2 μM). Fluvoxamine, a potent inhibitor of CYP1A2, inhibited azelastine N-demethylation (IC50 = 17.2 μM). Bufuralol (a substrate of CYP2D6), quinidine (an inhibitor of CYP2D6), and sulfaphenazole (an inhibitor of CYP2C9) inhibited weakly azelastineN-demethylation (IC50 = 84.4, 85.4, and 91.0 μM, respectively). The effects of furafylline (CYP1A2), coumarin (CYP2A6), S-mephenytoin (CYP2C19), and chlorzoxazone (CYP2E1) on the azelastineN-demethylase activities were small up to a concentration of 100 μM inhibitor.

Effects of CYP inhibitors on azelastine N-demethylase activity in human liver microsomes.
Azelastine N-demethylase activity in pooled human liver microsomes was determined at an azelastine concentration of 5 μM. The inhibitor concentration ranged from 0.1 to 100 μM. Fluvoxamine and furafylline (CYP1A2), coumarin (CYP2A6), sulfaphenazole (CYP2C9),S-mephenytoin (CYP2C19), bufuralol and quinidine (CYP2D6), chlorzoxazone (CYP2E1), and ketoconazole and erythromycin (CYP3A4) were used. The control activity was 83.0 pmol/min/mg. Each data point represents the mean of duplicate determinations.
Azelastine N-Demethylase Activity in Microsomes from B-Lymphoblast Cells or Baculovirus-Infected Insect Cells Expressing Human CYP.
The azelastine N-demethylase activity of recombinant CYPs was determined at a concentration of 5 μM azelastine (∼KM value of the high-affinity component). In recombinant CYPs that exhibited azelastineN-demethylase activity, the formation of desmethylazelastine at 5 μM azelastine increased linearly with incubation time up to 60 min. Unless specified, an incubation time of 30 min was used to ensure the initial rate condition for the formation of desmethylazelastine. Microsomes from two different expression systems, human B-lymphoblast cells (Fig. 6A) or baculovirus-infected insect cells (Fig. 6B), were compared. In the microsomes from B-lymphoblast cells, CYP2D6, CYP1A1, CYP3A4, CYP2C19, CYP2C9, and CYP1A2 exhibited azelastine N-demethylase activities of 0.324, 0.266, 0.037, 0.033, 0.019, and 0.010 pmol/min/pmol CYP, respectively. In the microsomes from baculovirus-infected insect cells, CYP2D6, CYP1A1, CYP1A2, CYP3A4 with b5, CYP3A4 without b5, CYP3A5, CYP2C19, CYP2C9, and CYP2C8 exhibited the azelastine N-demethylase activities of 1.018, 1.013, 0.425, 0.395, 0.366, 0.215, 0.199, 0.051, and 0.030 pmol/min/pmol CYP, respectively. In all CYP isoforms that exhibited detectable azelastine N-demethylase activities, the activities were higher in microsomes from baculovirus-infected insect cells than from B-lymphoblast cells (CYP1A1, 3.8-fold; CYP1A2, 42.5-fold; CYP2C9, 2.7-fold; CYP2C19, 6.0-fold; CYP2D6, 3.1-fold; CYP3A4, 9.9-fold).

Azelastine N-demethylase activity by recombinant CYPs.
Microsomes from human B-lymphoblast cells (A) or baculovirus-infected insect cells (B) were incubated with 5 μM azelastine. Each column represents the mean of duplicate determinations.
Kinetic analyses were performed using microsomes from B-lymphoblast cells or baculovirus-infected insect cells expressing human CYP (Table2). In the microsomes from B-lymphoblast cells or baculovirus-infected insect cells, CYP1A1 exhibited similarKM (15.9 versus 19.4 μM) and higherVmax in baculovirus-infected insect cells (1.13 versus 4.32 pmol/min/pmol CYP). In baculovirus-infected insect cells, the kinetic parameters of CYP1A2 were as follows:KM = 125.4 μM,Vmax = 16.45 pmol/min/pmol CYP. Because of the low activity, the kinetic parameters could not be calculated for recombinant CYP1A2 from B-lymphoblast cells. The microsomes from B-lymphoblast cells or baculovirus-infected insect cells exhibited different values for KM (36.1 versus 8.5 μM) and for Vmax (0.35 versus 0.60 pmol/min/pmol CYP) for CYP2C19, similar values forKM (1.4 versus 2.1 μM) and different values of Vmax (0.44 versus 1.60 pmol/min/pmol CYP) for CYP2D6, and different values forKM (74.1 versus 43.7 μM) and forVmax values (0.59 versus 2.67 pmol/min/pmol CYP) for CYP3A4. CYP3A4 coexpressed with or withoutb5 in the microsomes from baculovirus-infected insect cells exhibited similar values forKM (51.1 versus 43.7 μM). TheVmax values were higher for CYP3A4 coexpressed with b5 (4.45 versus 2.67 pmol/min/pmol CYP). For all CYP isoforms that showed azelastineN-demethylase activity, the intrinsic clearance (CL,Vmax/KM) values were higher in the microsomes from baculovirus-infected insect cells than in the microsomes from B-lymphoblast cells.
Kinetic parameters of azelastine N-demethylase activity in microsomes expressing human CYP in human B-lymphoblast cells or baculovirus-infected insect cells
Correlation between Azelastine N-Demethylase Activity and CYP1A2, CYP2D6, or CYP3A4 Activities in Human Liver Microsomes.
The azelastine N-demethylase activity (5 μM azelastine) in microsomes from twelve human livers (HLG1–HLG12) was compared with the CYP1A2-, CYP2D6-, and CYP3A4-mediated activities. EthoxyresorufinO-deethylase activity (2 μM ethoxyresorufin), bufuralol 1′-hydroxylase activity (1 μM bufuralol), and testosterone 6β-hydroxylase activity (100 μM testosterone) in human liver microsomes were determined as probes of CYP1A2, CYP2D6, and CYP3A4 activity, respectively. As shown in Fig.7, azelastine N-demethylase activity was significantly correlated with testosterone 6β-hydroxylase activity (r = 0.849, p< .0005). The azelastine N-demethylase activity at 1 μM azelastine in microsomes from six human livers (HLG1, 4, 6, 7, 10, and 11) was also significantly correlated with testosterone 6β-hydroxylase activity (r = 0.897, p< .05, data not shown). In contrast, the azelastineN-demethylase activities at both 1 and 5 μM azelastine were not correlated with either ethoxyresorufin O-deethylase activity or bufuralol 1′-hydroxylase activity.
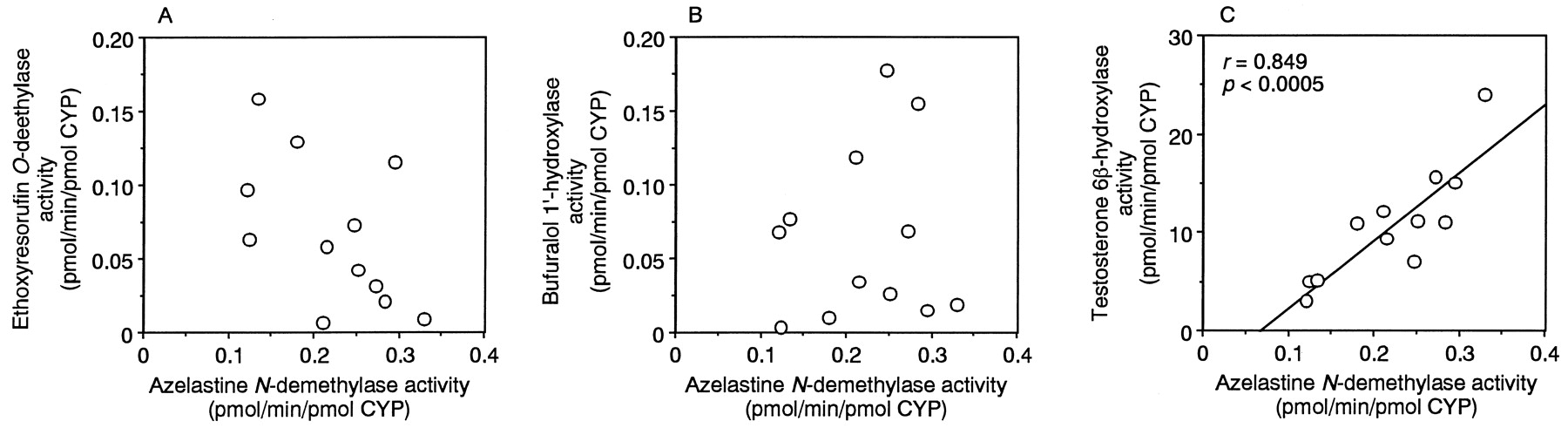
Relationship between azelastine N-demethylase activity and ethoxyresorufin O-deethylase, bufuralol 1′-hydroxylase, and testosterone 6β-hydroxylase activities in microsomes from twelve human livers.
Azelastine N-demethylase activity (5 μM) and ethoxyresorufin O-deethylase activity (2 μM) (A); azelastine N-demethylase activity and bufuralol 1′-hydroxylase activity (1 μM) (B); azelastineN-demethylase activity and testosterone 6β-hydroxylase activity (100 μM) (C).
Contributions of CYP1A2, CYP2D6, and CYP3A4 to AzelastineN-Demethylase Activity in Human Liver Microsomes.
The contributions of CYP1A2, CYP2D6, and CYP3A4 to the azelastineN-demethylase activity in microsomes from six human livers (HLG1, 4, 6, 7, 10, and 11) were estimated using the following prediction methods:
Prediction Method 1.
The azelastine N-demethylase activity in microsomes from six human livers at an azelastine concentration of 1 μM ranged from 12.2 to 53.9 pmol/min/mg. The immunochemically determined CYP contents (pmol/mg) supplied by Gentest are shown in Table3. Because it has been reported that the CYP3A content is principally CYP3A4, the CYP3A content was assumed to represent the CYP3A4 content. The vrec-CYP1A2, vrec-CYP2D6, and vrec-CYP3A4 from B-lymphoblast cells were 0, 0.206, and 0.009 pmol/min/pmol CYP, respectively. The vrec-CYP1A2, vrec-CYP2D6, vrec-CYP3A4 withoutb5, and vrec-CYP3A4with b5 from baculovirus-infected insect cells were 0.094, 0.657, 0.100, and 0.077 pmol/min/pmol CYP, respectively. When the activity of recombinant CYPs from B-lymphoblast cells was used, the sum of vCYP1A2, vCYP2D6, and vCYP3A4 did not reach the observed vHL at all. On the other hand, when the activity of recombinant CYPs from baculovirus-infected insect cells was used, the contributions of CYP1A2, CYP2D6, and CYP3A4 to the hepatic microsomal azelastine N-demethylase activity as estimated by eqs. 3 to 8 were 2.3 to 39.7, 0 to 32.3, and 20.9 to 68.7% (CYP3A4 without b5), and 16.1 to 52.9% (CYP3A4 with b5), respectively (Fig.8A). HLG4 did not have immunodetectable CYP2D6 protein or measurable bufuralol 1′-hydroxylase activity. Accordingly, vCYP2D6 in HLG4 was presumed to be zero.
Contribution of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation in human liver microsomes by prediction methods 1 and 2

The percent contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation in human liver microsomes.
Extrapolation was determined by prediction methods 1 (A), 2 (B), 3 (C), 4 (D), and 5 (E) using microsomes from six human livers (HLG1, 4, 6, 7, 10, and 11) and recombinant CYP1A2, CYP2D6, and CYP3A4 +b5 from baculovirus-infected insect cells. Prediction methods 1 and 2 were based on each CYP content in human liver microsomes and prediction methods 3 to 5 were based on three different RAFs, as described in Materials and Methods.
Prediction Method 2.
The azelastine N-demethylase activity in human liver microsomes and immunochemically determined CYP contents was the same as used for method 1 (Table 3). The kinetic parameters for recombinant CYP2D6 and CYP3A4 from B-lymphoblast cells were as follows;KM, CYP2D6 = 1.4 μM,Vmax, CYP2D6 = 0.44 pmol/min/pmol CYP,KM, CYP3A4 = 74.1 μM, andVmax, CYP3A4 = 0.59 pmol/min/pmol CYP. The kinetic parameters for recombinant CYP1A2, CYP2D6, and CYP3A4 from baculovirus-infected insect cells were as follows; KM, CYP1A2 = 125.4 μM, Vmax, CYP1A2 = 16.45 pmol/min/pmol CYP, KM, CYP2D6 = 2.1 μM, Vmax, CYP2D6= 1.6 pmol/min/pmol CYP, KM, CYP3A4 = 43.7 (without b5) and 51.1 μM (withb5), and Vmax, CYP3A4 = 2.67 (without b5) and 4.45 pmol/min/pmol CYP (with b5). When the kinetic parameters for recombinant CYPs from B-lymphoblast cells were used, the sum of vCYP1A2, vCYP2D6, and vCYP3A4 did not reach the observed azelastine N-demethylase activity (vHL) at all. On the other hand, when the kinetic parameters for recombinant CYPs from baculovirus-infected insect cells were used, the contributions of CYP1A2, CYP2D6, and CYP3A4 to the hepatic microsomal azelastine N-demethylase activity were estimated to be 3.2 to 54.9, 0 to 25.4, 12.5 to 41.0 (CYP3A4 withoutb5), and 17.9 to 58.7% (CYP3A4 withb5), respectively (Fig. 8B).
Prediction Method 3.
For prediction methods 3 to 5, data from recombinant CYP1A2, CYP2D6, and CYP3A4 coexpressed with b5 from baculovirus-infected insect cells were applied because the clearance of azelastine N-demethylation of recombinant CYP1A2 from B-lymphoblast cells was not obtained. The N-demethylation clearance of azelastine (CLHL,Vmax/KM) in microsomes from six human livers ranged from 38.0 to 91.2 μl/min/nmol CYP (Table 4). CLrec-CYP1A2, CLrec-CYP2D6, and CLrec-CYP3A4 were 131.2, 761.9, and 87.1 μl/min/nmol CYP, respectively. The ethoxyresorufinO-deethylase activity (2 μM ethoxyresorufin) in microsomes from the six human livers ranged from 6.3 to 157.7 fmol/min/pmol CYP, and for recombinant CYP1A2 it was 1.332 pmol/min/pmol CYP. Thus, RAFv, CYP1A2 was estimated to range from 0.005 to 0.118. The bufuralol 1′-hydroxylase activity (1 μM bufuralol) in microsomes from six human livers ranged from 0.010 to 0.118 pmol/min/pmol CYP, and for recombinant CYP2D6 it was 5.886 pmol/min/pmol CYP. In our study, bufuralol 1′-hydroxylase activity was detectable in HLG4. Thus, RAFv, CYP2D6 was estimated to range from 0.002 to 0.020. The testosterone 6β-hydroxylase activity (100 μM testosterone) in microsomes from six human livers ranged from 1.897 to 9.338 pmol/min/pmol CYP, and for recombinant CYP3A4 it was 35.092 pmol/min/pmol CYP. Thus, RAFv, CYP3A4 was estimated to range from 0.054 to 0.266. Therefore, the contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation clearance in human liver microsomes were estimated to be 0.9 to 36.6, 1.9 to 23.4, and 12.4 to 33.5%, respectively (Fig. 8C).
Contribution of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation in human liver microsomes by prediction methods 3 to 5
Prediction Method 4.
The Vmax values of ethoxyresorufinOdeethylase activity in microsomes from six human livers ranged from 9.3 to 261.3 fmol/min/pmol CYP, and for recombinant CYP1A2 it was 1.98 pmol/min/pmol CYP. Thus, RAFVmax, CYP1A2 was estimated to range from 0.005 to 0.132 (Table4). The Vmax values of bufuralol 1′-hydroxylase activity in microsomes from six human livers ranged from 0.116 to 0.531 pmol/min/pmol CYP, and for recombinant CYP2D6 it was 6.382 pmol/min/pmol CYP. Thus, RAFVmax, CYP2D6was estimated to range from 0.018 to 0.083. TheVmax values of testosterone 6β-hydroxylase activity in microsomes from six human livers ranged from 8.39 to 37.33 pmol/min/pmol CYP, and for recombinant CYP3A4 it was 34.89 pmol/min/pmol CYP. Thus, RAF Vmax, CYP3A4was estimated to range from 0.240 to 1.070. Therefore, the contributions of CYP1A2, CYP2D6, and CYP3A4 toN-demethylation clearance in human liver microsomes were estimated to be 0.9 to 40.8, 15.0 to 166.4, and 55.0 to 117.7%, respectively (Fig. 8D).
Prediction Method 5.
The KM values of the ethoxyresorufinOdeethylase activity in microsomes from six human livers and for recombinant CYP1A2 were 0.27 ± 0.19 and 0.05 μM, respectively. The CL values of the ethoxyresorufinO-deethylase activity in the microsomes ranged from 0.045 to 0.919 μl/min/fmol CYP, and for recombinant CYP1A2 it was 38.002 μl/min/fmol CYP. Thus, RAFCL, CYP1A2 was estimated to range from 0.001 to 0.024 (Table 4). TheKM values of the bufuralol 1′-hydroxylase activity in microsomes from five human livers, except HLG4, and for recombinant CYP2D6 were 6.0 ± 2.8 and 1.5 μM, respectively. TheKM value of the bufuralol 1′-hydroxylase activity in HLG4 was 30.1 μM. The CL values of bufuralol 1′-hydroxylase activity in microsomes from six human livers ranged from 5.3 to 150.2 μl/min/nmol CYP, and for recombinant CYP2D6 it was 4.35 μl/min/pmol CYP. Thus, RAFCL, CYP2D6 was estimated to range from 0.001 to 0.035. TheKM values of the testosterone 6β-hydroxylase activity in microsomes from six human livers and for recombinant CYP3A4 were 40.6 ± 11.1 and 37.0 μM, respectively. The CL values for testosterone 6β-hydroxylase activity in microsomes from six human livers ranged from 0.225 to 0.927 μl/min/pmol CYP, and for recombinant CYP3A4 it was 0.943 μl/min/pmol CYP. Thus, RAFCL, CYP3A4 was estimated to range from 0.239 to 0.983. Therefore, the contributions of CYP1A2, CYP2D6, and CYP3A4 to azelastine N-demethylation clearance in human liver microsomes were estimated to be 0.2 to 8.4, 1.4 to 45.1, and 48.9 to 101.6%, respectively (Fig. 8E). The sum of the contributions of CYP1A2, CYP2D6, and CYP3A4 to the N-demethylation clearance was almost 100% for each of the six human liver microsomes.
Discussion
Azelastine has been reported to be metabolized to desmethylazelastine, 6-hydroxyazelatine, 7-oxoazelastine, 4-[(4-chlorophenyl)methyl]-2-(5-methylamino-1-carboxy-2-pentyl)-1(2H)-phthalazinone, and 4-[(4-chlorophenyl)methyl]-2-(5-methylamino-1-carboxy-3-pentyl)-1(2H)-phthalazinone in experimental animals (Tatsumi et al., 1984; Yang et al., 1992;Adusumalli et al., 1992). Desmethylazelastine has been also detected in human plasma as a metabolite of azelastine after oral administration (Pivonka et al., 1987). In the present study, azelastine was metabolized by human liver microsomes to only one metabolite, desmethylazelastine (Fig. 2). The conversion of azelastine to desmethylazelastine was dependent on NADPH and active protein in human liver microsomes. The biphasic Eadie-Hofstee plots for azelastineN-demethylase activity in human liver microsomes indicated that multiple enzymes were responsible for the reaction.
The inhibitory effects of ketoconazole, erythromycin, and fluvoxamine on azelastine N-demethylase activity in human liver microsomes suggested that CYP3A4 and CYP1A2 contribute to azelastineN-demethylation. Furthermore, the weak inhibition by bufuralol and quinidine suggested that CYP2D6 also participates in azelastine N-demethylation. In the kinetic studies, the inhibition of the high-affinity component of azelastineN-demethylation in human liver microsomes by bufuralol suggested that CYP2D6 is the high-affinity enzyme. In addition, the inhibition by 1 μM ketoconazole indicated that CYP3A4 contributes to both (high- and low-affinity) components of azelastineN-demethylation in human liver microsomes. The azelastineNdemethylase activity at 5 μM azelastine was significantly (r = 0.849, p < .0005) correlated with testosterone 6β-hydroxylase activity in microsomes from twelve human livers, supporting a major role for CYP3A4. It has been reported that the Cmax value of azelastine in humans after repetitive oral administration of azelastine hydrochloride (4.4 mg every 12 h) was 16.5 nM (Morganroth et al., 1997). However, azelastine has been reported to accumulate in the liver to a level almost 50-fold higher than the plasma concentration in experimental animals (Tatsumi et al., 1980, 1984). Accordingly, the concentration of azelastine in human livers could be projected to accumulate to up to 0.8 μM. Then, the correlation study was performed with azelastine N-demethylase activity at 1 μM azelastine in microsomes from six human livers. The significant correlation with testosterone 6β-hydroxylase activity suggested that CYP3A4 might be a principal enzyme that is responsible for azelastineN-demethylation in human liver microsomes even at a low concentration.
The azelastine N-demethylase activity for recombinant CYPs from baculovirus-infected insect cells was generally higher than that from B-lymphoblast cells. OR was not coexpressed only in recombinant CYP1A2 and CYP2C19 from B-lymphoblast cells, and that might affect the difference of activity between two systems. In addition, theKM values of recombinant CYP2C19 and CYP3A4 in the two expression systems were also different from each other. These differences between the two expression systems might be due to the differences in OR expression levels and/or lipid composition. In the baculovirus-infected insect cells, the Vmaxvalue of the azelastine N-demethylase activity of CYP3A4 coexpressed with b5 was 1.7-fold higher than that of CYP3A4 without b5. The effects of b5 appears to depend on the particular CYP3A substrate (Gillam et al., 1995; Yamazaki et al., 1996a, b). Coexpressed b5 did not affect theKM value for azelastineN-demethylation, although it increased theVmax value. The addition ofb5 in a CYP reconstitution system has been reported to decrease the KM value (Kamataki et al., 1983; Yamazaki et al., 1996b). Differences in effects ofb5 on KM values might be due to difference in coexpression or exogenous supplementation of b5.
The KM values of recombinant CYP2D6 and CYP3A4 for azelastine N-demethylation were similar to those of the high- and low-affinity component in human liver microsomes, respectively. It has been reported that CYP1A1 is not expressed in human livers and that CYP2C19 accounts for only 1% of the total CYP (McManus et al., 1990; Shimada et al., 1994; Inoue et al., 1997). Accordingly, it was also expected that the contribution of CYP1A1 and CYP2C19 to azelastine N-demethylation in human livers would be negligible or minor. Therefore, the contributions of CYP1A2, CYP2D6, and CYP3A4 to the activity in individual microsomes from different human livers were investigated in detail.
Extrapolation from in vitro data to intrinsic clearance of drug in humans has received increasing attention recently because it is important for predicting drug-drug interactions in vivo. The best prediction methods for such extrapolation have not, however, agreed. The first prediction approach was based on the simplest interpretation of the capacity of a particular CYP to metabolize a drug and the content of the CYP protein in human liver microsomes (prediction method 1). Then, the second approach considered the affinity (KM) and capacity (Vmax) of enzymes, the substrate concentration and content of the CYP protein, to estimate the contributions of each CYP isoform (prediction method 2). To reflect the concentration of azelastine in human livers in clinical use, the percent contributions of each CYP isoform at 1 μM azelastine were estimated. When the activity or kinetic parameters for the azelastineN-demethylation of recombinant CYP1A2, CYP2D6, and CYP3A4 from B-lymphoblast cells were used, the sum of vCYP1A2, vCYP2D6, and vCYP3A4 did not reach the observed vHL, owing to the relatively low azelastineN-demethylase activity in the CYPs from B-lymphoblast cells. These results would suggest that the use of B-lymphoblast cells is not appropriate for prediction, at least in the case of azelastineN-demethylation. These results were consistent with a previous report by Yamazaki et al. (1997) in which recombinant CYPs expressed in baculovirus-infected insect cells exhibited a proper prediction of omeprazole oxidation, but those expressed in B-lymphoblast cells or yeast did not. One limitation to these extrapolation methods is that the CYP contents were measured by immunoblot analysis. There are isoforms that can not be distinguished from other isoforms by immunoblot analysis because of high cross-reactivity. In addition, active and inactive protein cannot be distinguishable by immunoblot analysis. Furthermore, distinct enzymes may have different affinities for the coenzymes necessary for catalytic activity, which serves to unlink the CYP content and its catalytic activity. Therefore, an approach using the catalytic activity for extrapolation was advocated as described below.
Crespi (1995) first proposed the concept of RAF to extrapolate the data obtained from recombinant CYPs to those from human liver microsomes. We applied RAF to estimate the relative contributions of CYP1A2, CYP2D6, and CYP3A4 to the N-demethylation clearance of azelastine in individual human liver microsomes. RAFVmax was proposed first (Crespi, 1995), and then RAFv was also applied for prediction (Kobayashi et al., 1997; Nakajima et al., 1998). It has been suggested that the in vitro CL (Vmax/KM) is most appropriate for the calculation of absolute scaling factor values because the enzymes usually will not become saturated in vivo, and the absolute drug concentration at the site of metabolism cannot be measured (Crespi, 1995). Therefore, we evaluated the application of three different RAFs to predict the contribution of each CYP isoform.
In prediction method 3, the ethoxyresorufin O-deethylase activity in human liver microsomes and recombinant CYPs were determined at a substrate concentration of 2 μM (∼40 times theKM value for recombinant CYP1A2). Accordingly, RAFv, CYP1A2 should be close to RAFVmax, CYP1A2 in prediction method 4. The maximum difference in KM values between human liver microsomes and recombinant CYP1A2 was 10-fold. Thus, the difference in KM values would reflect the difference between RAFVmax, CYP1A2 and RAFCL, CYP1A2 in prediction methods 4 and 5. The HLG4 microsomes might be from a poor metabolizer of CYP2D6. TheKM value of bufuralol 1′-hydroxylase activity in HLG4 was higher than those in the other human liver microsomes. As the KM value of bufuralol 1′-hydroxylase activity of CYP1A2 expressed in baculovirus-infected insect cells was 38.1 μM, it is suggested that the bufuralol 1′-hydroxylase activity detected in HLG4 might be catalyzed by CYP1A2. In prediction method 3, because the bufuralol 1′-hydroxylase activity was determined at a substrate concentration of 1 μM (∼KM value of recombinant CYP2D6), there would be little contribution of CYP1A2 to the bufuralol 1′-hydroxylase activity. In these conditions, RAFv, CYP2D6 would be applicable for the extrapolation. In prediction method 3, when testosterone 6β-hydroxylase activity was determined at a substrate concentration of 100 μM (∼2.7-times theKM value for recombinant CYP3A4), RAFv, CYP3A4 was smaller than the RAFCL, CYP3A4 calculated in prediction method 5. It is suggested that RAF as the ratio of activity in a nonsaturating substrate condition might result in underestimation. The difference in the KM values in human liver microsomes and recombinant CYP3A4 was 1.5-fold at most. Thus, the percent contributions of CYP3A4 were similar for prediction methods 4 and 5.
Eadie-Hofstee plots of azelastine N-demethylation for microsomes from HLG1, 4, and 6 were monophasic, but were biphasic for microsomes from HLG7, 10, and 11. Therefore, demethylation clearance of azelastine in human liver microsomes (CLHL) was estimated as the rate of metabolism under nonsaturating conditions, because it is considered to be comparable to the clearance in vivo. The monophasic Eadie-Hofstee plot in microsomes from HLG4 agreed with the presumption of estimation (Fig. 8) that HLG4 lacks CYP2D6 and that CYP3A4 mainly catalyzes azelastine N-demethylation. On the other hand, it is unclear why the Eadie-Hofstee plots in microsomes from HLG1 and 6 showed monophasic pattern. However, monophasic Eadie-Hofstee plot do not necessarily indicate that only one enzyme is involved in the reaction. It is suspected that the plots appeared to be monophasic; nevertheless, multiple enzymes contribute to azelastineN-demethylation.
We conclude that the using RAF as the ratio of CL and the data for recombinant CYPs from baculovirus-infected insect cells appears to be the most appropriate approach for estimating the contributions of CYPs involved in certain types of drug metabolism because the total of the predicted clearances showed the best reflection of the observed clearance. However, several problems remain to be solved: 1) Does the RAFCL provide the most appropriate prediction for any types of drug metabolism? 2) Is the RAF of each CYP isoform independent of the kind of marker activity? 3) Are the data using recombinant CYPs from baculovirus-infected cells most appropriate for any types of drug metabolism? 4) Does coexpression of OR orb5 in the expression system affect the prediction? In addition, when an in vitro metabolic study is performed using recombinant CYPs, it is important to keep the following points in mind: 1) In the expression system, the content of OR andb5 will be different from that in human livers. 2) The effects of OR and b5 will depend on the chemical(s) involved. 3) It is not known whether interindividual differences in the b5content or NADH-b5 reductase contribute to CYP activities in vivo. Additional study is now underway in our laboratory to solve these problems.
Inhibitors used at high concentration also inhibit other CYP isoforms nonspecifically, even if the inhibitor is known as a specific inhibitor for certain CYP isoform. In fact, when inhibitors were used at 100 μM, the sum of the total inhibition was far greater than 100%. However, the percent inhibitions using lower concentrations of inhibitor gave more rational results. For example, using 10 μM ketoconazole, bufuralol, and furafylline, the inhibition values were 80% for CYP3A4, 10% for CYP2D6, and 10% for CYP1A2, respectively (see Fig. 4). Thus, the results of the inhibition studies supported the percent contribution obtained from recombinant CYP approach.
In conclusion, data from the present study show evidence that azelastine N-demethylation is catalyzed mainly by CYP3A4 and CYP2D6, and CYP1A2 to a small extent, in humans (in average, 76.6, 21.8, and 3.9%, respectively). The percent contributions of these CYPs differ in individuals. It is suggested that there would be some possibility of drug interactions via CYP3A4 and CYP2D6, when drugs that affect those CYPs or are metabolized by them are coadministered with the azelastine.
Acknowledgment
We thank Brent Bell for reviewing the manuscript.
Footnotes
-
Send reprint requests to: Miki Nakajima, Ph.D., Division of Drug Metabolism, Faculty of Pharmaceutical Sciences, Kanazawa University, Takara-machi 13–1, Kanazawa 920-0934, Japan. E-mail:nmiki{at}kenroku.kanazawa-u.ac.jp
- Abbreviations used are::
- CYP
- cytochrome P-450
- b5
- cytochromeb5
- OR
- NADPH-CYP oxidoreductase
- RAF
- relative activity factor
- RAFv
- RAF as the ratio of activity
- RAFVmax
- RAF as the ratio ofVmax value
- RAFCL
- RAF as the ratio of clearance
- Received March 1, 1999.
- Accepted July 12, 1999.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}