


Abstract
A human UDP-glucuronosyltransferase (UGT) catalyzing the glucuronidation of morphine has been identified. A full length cDNA was isolated from a human liver cDNA library and found to be identical to the UGT2B7 form having a tyrosine at position 268. This cDNA was transfected into HK 293 cells, and stable expression was achieved. Cell homogenates and membrane preparations from HK 293 cells expressing UGT2B7 catalyzed the glucuronidation of morphine and other clinically significant opioid agonists, antagonists, and partial agonists. UGT2B7 catalyzed morphine glucuronidation at the 3- and 6-hydroxy positions and also mediated the formation of codeine-6-glucuronide from codeine. This represents the first demonstration of a UGT capable of catalyzing the glucuronidation of both the 3- and 6-positions of opioids. Since humans excrete morphine-3-glucuronide and morphine-6-glucuronide after morphine administration, it is likely that UGT2B7 is a major isoform in humans responsible for the metabolism of this important drug and its surrogates.
Glucuronide conjugation is an important metabolic process in which xenobiotics and endobiotics are converted to hydrophilic metabolites through the mediation of UGTs1 (E.C. 2.4.1.17). UGTs are intrinsic membrane proteins of the endoplasmic reticulum and nuclear envelope of cells of liver and other organs, and are encoded by multiple genes of at least two families (1). After glucuronidation, conjugates are readily excreted by renal and hepatic mechanisms leading to rapid elimination of the aglycone from the body. At this time, >30 individual UGT isoforms have been identified either by cloning and expression or by purification procedures from hepatic microsomes. UGTs that are members of the UGT1 gene complex share common second through fifth exons, with at least seven separate exons encoding for specific proteins with unique N-terminal domains (2). In contrast, gene products of the UGT2 family are transcribed from unique genes. At least eight isozymes of the human UGT2 family have been identified (3), and the genes UGT2B4, UGT2B9, andUGT2B15 have been mapped to human chromosome 4 (4, 5).
Xenobiotics such as those of the opioid drug class are, in general, metabolized and excreted largely as glucuronides by the liver and kidney. Morphine is the best example of an opioid drug that is extensively glucuronidated. Morphine possesses hydroxy groups on the 3- and 6- positions of the molecule and both morphine-3-glucuronide and morphine-6-glucuronide are found in the urine of humans (6, 7), whereas codeine with a methoxy group on the 3-position is converted only to the 6-glucuronide.
A principal interest in this laboratory for many years has been the isolation and identification of the UGTs that catalyze the glucuronidation of opioids in rats and humans (8-10). We have recently shown that stably expressed human UGT1.1, often termed the major “bilirubin” UGT (11), reacts with high affinity and efficiency with the opioid partial agonist/antagonist, buprenorphine (12). Previous investigations by Miners et al.(13) and Wahlström et al.(14) suggested, that several isozymes mediate hepatic opioid glucuronidation in humans; however, no expressed human UGTs (other than UGT1.1) have been reported to participate in opioid glucuronidation.
This communication describes the cloning and expression of a cDNA encoding for a human liver microsomal UGT that catalyzes the glucuronidation of morphine and other opioids. Sequence analysis showed that the cDNA was identical to a human UGT, termed UGT2B7 variant (15), which had been shown previously to code for a UGT mediating glucuronidation of NSAIDs and certain forms of carboxylic acid-containing drugs. This UGT, UGT2B7, is likely to be the major isoform responsible for morphine glucuronidation in humans.
Materials and Methods
Chemicals.
UDP-GlcUA (319 mCi/mmol) was purchased from ICN Pharmaceuticals (Irvine, CA). Morphine sulfate was acquired from Merck (Rahway, NJ). Morphine-3-glucuronide, morphine-6-glucuronide, and codeine-6-glucuronide were acquired from Research Biochemicals (Natick, MA). All aglycones and l-α-phosphatidylcholine (type XVI-E from egg yolk), dithiothreitol, and UDP-GlcUA were purchased from Sigma Chemical Co. (St. Louis, MO). Protein assay reagents were from Bio-Rad (Richmond, CA).
Screening of the cDNA Library.
A human liver cDNA library in Bluescript SK (Stratagene, La Jolla, CA) was plated in XL1-Blue MRF′ cells. The library was screened by plaque hybridization with a full length rat liver cDNA, UGT2B1 (kindly supplied by Dr. Peter Mackenzie, Flinders Medical Centre, Bedford Park, South Australia, Australia). The probe was labeled with32P-dCTP by the nick-translation method following the protocol of the supplier (Boehringer Mannheim Biochemica, Indianapolis, IN), and hybridization was performed at 42°C in 50% formamide, 4.8 × standard saline citrate, 1 × Denhardt’s solution, 10% dextran, and 0.1% SDS containing salmon testes DNA at 0.1 mg/ml. Bacteriophages that hybridized to the probe were isolated and purified.
Restriction enzyme analysis showed that one clone (HM1) contained a 2.2 kb pair insert. This clone was further characterized.
DNA Sequence Analysis.
Analysis was performed by the DNA Facility, University of Iowa, using an Applied Biosystems 373 A Automatic DNA Sequencer. The HM1 insert was initially sequenced in the Bluescript plasmid using primers to the T3 and T7 promoters. Subsequently, oligonucleotides were synthesized (Genosys Biotechnologies, The Woodlands, TX) and used as primers to generate additional and overlapping DNA sequences until both strands were sequenced.
Expression of HM1 in Human Embryonic Kidney (HK 293) Cells.
The HM1 insert was removed from Bluescript SK and was ligated into theBamH1/Xho1 site of the mammalian expression vector pcDNA3.1 (Invitrogen, San Diego, CA). This expression vector construct is referred to as pcDNA3-2B. The HK 293 cells were transfected using the calcium phosphate transfection method (16) and were grown in media containing geneticin (G418) (17). Cells were harvested after 1–4 passages of stable transfection for analysis of glucuronidation activity.
Glucuronidation Assays.
Cells expressing pcDNA3-2B protein were resuspended in TBS containing 0.5 mM dithiothreitol and disrupted by freeze-thawing 3 times in liquid nitrogen before homogenization. Glucuronidation activity toward opioids [5 mM morphine (pH 8.4), 5 mM codeine (pH 7.7), 2 mM nalorphine (pH 8.4), 2 mM naloxone (pH 7.7), 2 mM naltrexone (pH 7.0), and 0.5 mM buprenorphine (pH 7.0)] was assayed using the method described by Puig and Tephly (9). Determination of the kinetic parameters for glucuronidation of opioids was performed on membrane preparations from cells expressing pcDNA3-2B protein, prepared as described by Battagliaet al. (18). Membrane preparations were frozen in aliquots. Membranes were resuspended in TBS containing 0.5 mM dithiothreitol and homogenized by hand in a Potter-Elvehjem homogenizer. The glucuronides of morphine-3, morphine-6, and codeine-6 were identified and quantified using a modified HPLC method as described by Yue et al. (19). For HPLC analysis of morphine-3-glucuronide, morphine-6-glucuronide, and codeine-6-glucuronide conjugates, the samples were eluted from the Sep-Pak C18 cartridges with 2 ml of 15% acetonitrile in phosphate buffer at pH 2.1. Morphine-3-glucuronide and morphine-6-glucuronide or codeine-6-glucuronide standards were added to 400 μl of the eluent that was injected into the HPLC system and eluted with 25% acetonitrile in phosphate buffer (pH 2.1) and 1 mM SDS. The radioactivity eluted with the glucuronide standards was measured. Analysis of glucuronidation of nonopioids was conducted and analyzed as described by Green et al. (17).
Results
Screening and Nucleotide Sequence of HM1.
Pritchard et al. (20) has previous reported that rat liver UGT2B1 exhibited glucuronidation activity toward morphine. Therefore, a human liver cDNA library was screened with the full-length cDNA, UGT2B1. Of 27 cDNA clones identified, two were full-length clones. One of these clones (HM1) was characterized and found to be identical to the human UGT2B7 variant as reported by Jin et al. (15). The deduced amino acid sequence has a tyrosine in position 268 instead of a histidine, as reported by Ritter et al. (21). Because there is no knowledge of the relative abundance of the two forms, there seems to be no reason to term either form as a variant at this time. Thus, we have designated HM1 as UGT2B7(Y) and the histidine form as UGT2B7(H).
Expression of UGT2B7(Y) in HK293 Cells.
Stably expressed UGT2B7(Y) catalyzes the glucuronidation of opioids (table 1), whereas opioid glucuronidation by vector-transfected and nontransfected HK293 cells was not detected. Of the opioids tested with stably expressed UGY2B7(Y), the glucuronidation rates for morphine, naloxone, and nalorphine were similar, whereas the rates for naltrexone and buprenorphine were <10% of that determined for morphine. The rates of codeine-6-glucuronidation were only ∼ 1% of the rate of morphine glucuronidation.
Glucuronidation of opioids by human UGT2B7(Y) expressed in HK 293 cells
Expressed UGT2B7(Y) had been shown previously to glucuronidate 4-hydroxyestrone and 4-hydroxyestradiol, and carboxylic acid-containing NSAIDs, such as ibuprofen and naproxen (15). Glucuronidation activities toward the four mentioned substrates were observed when the UGT2B7(Y) expressed in HK293 cells was studied using homogenate preparation (data not shown).
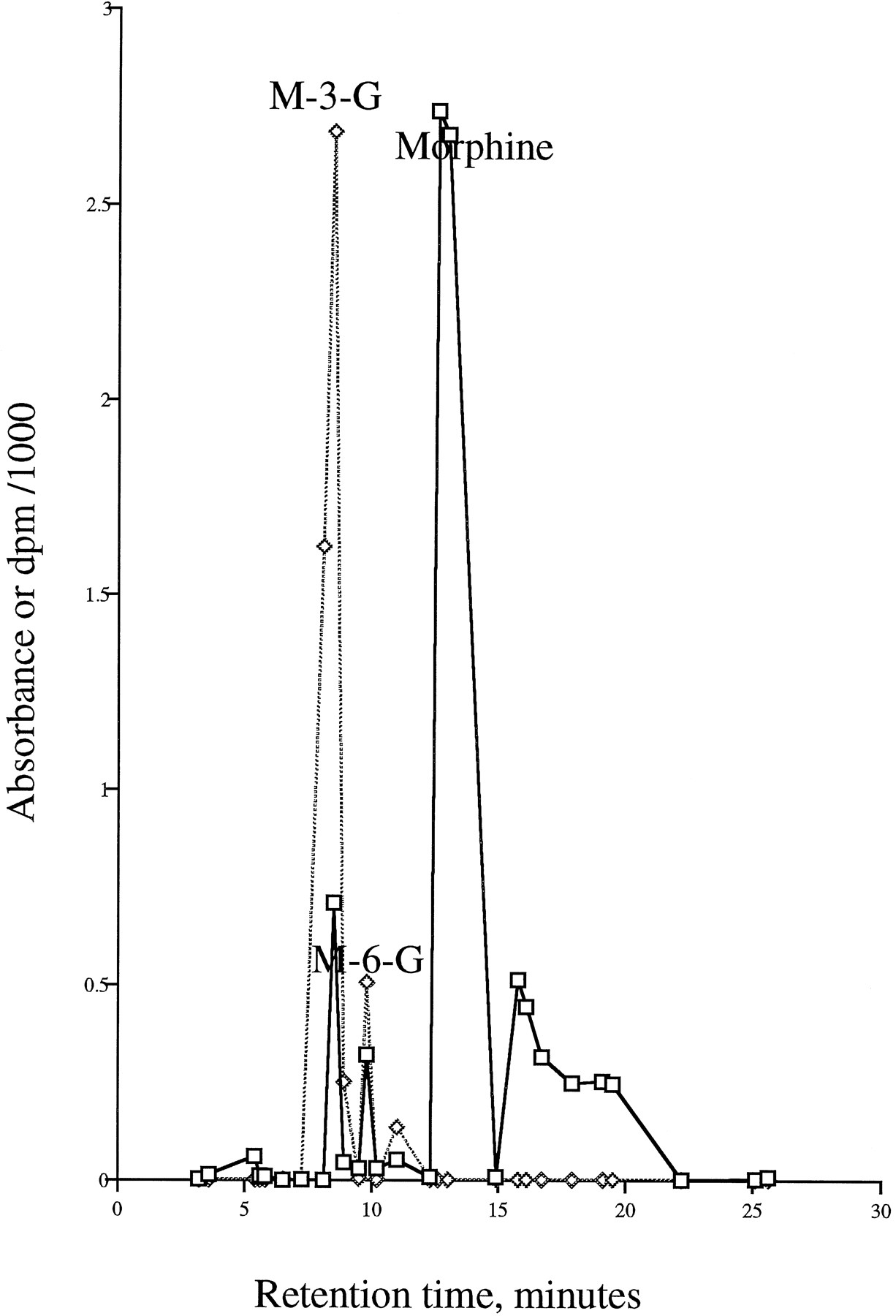
The HPLC chromatogram of the products from the glucuronidation of morphine by the stably expressed UGT2B7(Y) using [14C]UDP-GlcUA is shown in fig. 1. Carrier morphine-3-glucuronide and morphine-6-glucuronide were added to the incubation samples before HPLC analysis, and radioactivity coeluting with standards was measured. Both morphine-3-glucuronide and morphine-6-glucuronide were formed in reactions catalyzed by UGT2B7(Y). Preliminary results have also shown that expressed UGT2B7(H) also catalyzes morphine-3-glucuronide and morphine-6-glucuronidation (data not shown).2 Likewise, the HPLC analysis of the glucuronidation of codeine showed that radioactivity eluted along with the codeine-6-glucuronide standard (data not shown). This is the first demonstration of a UGT catalyzing the formation of an opioid-6-glucuronide.

HPLC chromatogram of a preparation from an incubation of morphine with a homogenate of HK 293 cells stably expressing UGT2B7(Y).
Experimental procedures are given in Materials and Methods.□, Absorbance at 210 nm; ◊, radioactivity dpm/1000.
Kinetic parameters were determined for the glucuronidation of morphine, codeine, nalorphine, and buprenorphine using membrane preparations from the HK293 cells expressing UGT2B7(Y) (table 2). Nalorphine and buprenorphine were found to produce substrate inhibition above 1.2 mM and 0.3 mM substrate, respectively. Buprenorphine and nalorphine are glucuronidated with higher efficiency than morphine, which is probably due to the increased length of the N-alkyl side chain of these opioids. The efficiency of codeine glucuronidation was 8 times less than that of morphine-6-glucuronidation.
Kinetic parameters of glucuronide formation in a membrane preparation from HK 293 cells stably expressing UGT2B7(Y)
Discussion
Morphine is metabolized in humans to morphine-3-glucuronide and morphine-6-glucuronide; but, until now, no human UGT has been identified to catalyze the formation of morphine-3-glucuronide and morphine-6-glucuronide with significant efficiency. UGT1.1, which also catalyzes opioid glucuronidation (12), does not catalyze the formation of morphine-6-glucuronide (unpublished data). The ratio ofVmax for the formation of morphine-3-glucuronidevs. morphine-6-glucuronide by the expressed UGT2B7(Y) was found to be similar to ratios reported for human liver microsomes (∼ 7:1) (19, 22). These data suggest that the UGT2B7(Y) represents the major human morphine UGT catalyzing the glucuronidation of the 3-OH and the 6-OH positions in opioids. Morphine-6-glucuronide has been shown to be a potent analgesic in clinical studies in human subjects, and the analgesic properties of morphine in humans are enhanced by the action of morphine-6-glucuronide (23). Morphine-3-glucuronide, in contrast, has no analgesic effect but shows enhanced excitatory activity by a nonopioid mechanism (24). It has been suggested that certain adverse effects of morphine in humans is due to the morphine-3-glucuronide formed. Thus, glucuronidation processes in the case of opioid pharmacology (e.g. morphine) may lead to potentially enhanced drug action or adverse effects not usually considered in the process of glucuronidation of most drugs.
1,4-Benzodiazepine derivatives, such as diazepam and halazepam, are converted in vivo to oxazepam, an active metabolite that is subsequently excreted as the pharmacologically inactive diasterometric oxazepam glucuronide. Based on inhibition studies, Patel et al. (25) have suggested that the S and Rforms of oxazepam are glucuronidated by different UGT isozymes, and that UGT2B7 is the enzyme catalyzing the formation ofS-oxazepam glucuronide. Patel et al. (26) also have found that 10% of a population had a significantly lower ratio ofS- to R-glucuronide metabolites. This result correlated with the results of an in vitro study using microsomes from human livers, where 10% of the livers displayed an abnormal high KM for the formation of theS-glucuronide, but no differences were found forKM for the R-glucuronide formation. These investigators speculated that the differences could be caused by a polymorphism in the UGT2B7 gene, yet the only difference in the proteins of UGT2B7(H) and UGT2B7(Y) is a histidine instead of a tyrosine at the 268 position. This amino acid difference does not seem to alter the glucuronidation ratio of morphine-3-glucuronide to morphine-6-glucuronide.2 Whether the difference in one amino acid affects the glucuronidation of substrates other than morphine is unknown at the present time. In addition, the question remains as to which UGT2B7 form is predominant in the human population.
Buprenorphine and naltrexone are antagonists that are under investigation as potential drugs for the treatment of opioid (27) and alcohol (28) abuse, respectively. Stably expressed human UGT1.1 also catalyzes the glucuronidation of opioids, in addition to bilirubin. This isoform reacts with high affinity toward buprenorphine, but exhibits a low efficiency of glucuronidation toward naltrexone and nalorphine (12), which may explain why liver microsomes from Crigler-Najjar type I patients, who do not express an active UGT1.1 due to a defect in the first exon of the UGT1 gene, glucuronidate buprenorphine at low rates (12). Naltrexone is glucuronidated at normal rates in Crigler-Najjar patients (12) most likely because UGT2B7 is the major isoform involved in its glucuronidation. However, the use of naltrexone or buprenorphine in patients with perturbation of liver function, as in alcohol addiction, might pose some problems dependent on the adequacy of the UGT2B7, UGT1.1, and UDP-GlcUA levels in such patients. Moreover, possible drug–endobiotic interaction must be considered when morphine is administered with such substrates that also interact with UGT2B7 (e.g. catechol estrogens). Drug–drug interactions should likewise be considered, because UGT2B7 is a major isoform catalyzing NSAID and benzodiazepine glucuronidation. Further studies on the substrate specificity and kinetic analysis for UGT2B7(Y) and UGT2B7(H) need to be investigated to identify possible drug–endobiotic and/or drug–drug interactions.
Footnotes
-
Send reprint requests to: Dr. Thomas R. Tephly, 2-459 Bowen Science Building, Department of Pharmacology, University of Iowa, Iowa City, IA 52242.
-
This research was supported by the National Institutes of Health Grant GM26221.
-
↵2 T. R. Tephly et al., unpublished data.
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- NSAID
- nonsteroidal anti-inflammatory drug
- UDP-GlcUA
- UDP-glucuronic acid
- SDS
- sodium dodecyl sulfate
- TBS
- 10mM Tris, 0.9% NaCl, pH 7.4
- OH
- hydroxy
- Received September 19, 1996.
- Accepted October 28, 1996.
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}