


Abstract
In this study, we report the effect of methanol, dimethyl sulfoxide (DMSO), and acetonitrile on the cytochrome P450 (P450)-mediated metabolism of several substrates in human liver microsomes: phenacetinO-deethylation for P4501A2, coumarin 7-hydroxylation for P4502A6, tolbutamide hydroxylation for P4502C8/2C9,S-mephenytoin 4′-hydroxylation for P4502C19, dextromethorphan O-demethylation for P4502D6, chlorzoxazone 6-hydroxylation for P4502E1, and testosterone 6β-hydroxylation for P4503A4. DMSO was found to inhibit several P450-mediated reactions (2C8/2C9, 2C19, 2E1, and 3A4) even at low concentrations (0.2%). There was no measurable effect on the catalytic activity of the various P450s when methanol was present at levels ≤1%, except for P4502C8/9 and 2E1. Acetonitrile did not noticeably change the catalytic activity of the P4502C8/2C9, 2C19, 2D6, and 2E1 enzymes at concentrations ≤1%. It was found that the content level of the organic solvents should be kept lower than 1% because, for all three solvents, a concentration of 5% strongly affected the metabolism of the various probes. These findings should be taken into consideration when designing in vitrometabolism studies of new chemical entities.
It has become increasingly common to use in vitro metabolism studies in liver microsomes, slices, and hepatocytes to predict in vivobiotransformations of new chemical entities. These studies include metabolite identification as well as determination of kinetic parameters such as Michaelis-Menton parameters (Km , Vmax) and inhibition constants (Ki ). These studies are used for many reasons including prediction of metabolic pathwaysin vivo, identification of enzymes responsible for metabolism, calculation of in vivo intrinsic clearance fromin vitro intrinsic clearance, and prediction of drug-drug interactions.
Many new chemical entities are highly lipophilic and require water-miscible organic solvents for effective solubilization. It is known that some organic solvents can affect the activity of enzymes involved in the biotransformations of exogenous compounds. These effects can be associated with solvation properties or competitive metabolism of the solvents for the enzyme in question. For example, it is known that dimethyl sulfoxide (DMSO)1 is a substrate for NADPH-dependent microsomal metabolizing enzymes (Gerhards and Gibian, 1967) and that methanol and ethanol are substrates of the alcohol-metabolizing enzyme systems (Teschke et al., 1975). Several years ago, a general study on the effect of solvents on the drug metabolism of commonly employed substrates in the Ames test was published. It was shown that the activity of the various substrates in the rat subcellular fractions could be inhibited or stimulated depending on the substrate and solvent used (Kawalek and Andrews, 1980).
Cytochrome P450 (P450) enzymes play an important role in the metabolism of exogenous compounds. It is possible to find studies that pinpoint the effect of organic solvents on the activity of some known P450 enzymes. For example, it has been reported, using specific cytochrome P450 substrates, that DMSO can inhibit the activity of P4502E1 and that several organic solvents can affect the activity of P4502A6 (Yooet al., 1987; Draper and Parkinson, 1997). However, to our knowledge, no systematic study clearly showing the effect of common organic solvents on the activity of the important P450 enzymes has been published. Such data would be valuable for the interpretation ofin vitro studies in which organic solvents are essential for effective solubilization of the chemicals.
In this study, we report the effect of methanol, DMSO, and acetonitrile on the cytochrome P450-mediated hepatic activity of several substrates in human microsomes: phenacetin O-deethylation for P4501A2, coumarin 7-hydroxylation for P4502A6, tolbutamide hydroxylation for P4502C8/2C9, S-mephenytoin 4′-hydroxylation for P4502C19, dextromethorphan O-demethylation for P4502D6, chlorzoxazone 6-hydroxylation for P4502E1, and testosterone 6β-hydroxylation for P4503A4. The particular P450s studied were selected on the basis that they constitute the major P450s present in human liver (Shimadaet al., 1994).
Materials and Methods
Chemicals.
β-Nicotinamide adenine dinucleotide phosphate, EDTA,d-glucose-6-phosphate, glucose 6-phosphate dehydrogenase, acetaminophen, chlorzoxazone, coumarin, dextromethorphan, phenacetin, tolbutamide, 7-hydroxycoumarin, and 6-hydroxychlorzoxazone were purchased from Sigma. 6β-Hydroxytestosterone and testosterone were obtained from Steraloids (Wilton, NH). Bovine serum albumin, Folin & Ciocalteu’s Phenol Reagent, and the modified Lowry protein assay reagent were purchased from Pierce. Dextrorphan d-tartrate and 4-hydroxytolbutamide were purchased from RBI (Natick, MA). (±)-4′-Hydroxymephenytoin and S(+)-mephenytoin were obtained from Ultrafine Chemicals (Manchester, UK). All other reagents were of highest purity commercially available or HPLC grade.
Tissue and Microsomes.
Human tissues were obtained from various sources (F. Guengerich, Vanderbilt University School of Medicine, Nashville, TN; IIAM, Exton, PA; Québec Transplant, Montréal, Canada). Microsomes were prepared from frozen (−80°C) tissue as described in the literature (Lu and Levin, 1972). Protein concentrations of the microsomal fractions were determined by the method of Lowry et al.(1951) using bovine serum albumin as a standard. For this study, a human microsome mixture prepared from six different livers was used.
Enzymatic Assay.
The microsomal incubation conditions used to study the metabolism of the phenacetin, coumarin, tolbutamide, S-mephenytoin, dextromethorphan, chlorzoxazone, and testosterone have been reported in a previous paper (Chauret et al., 1997). Briefly, each incubation was performed with microsomal protein in a 100 mM phosphate buffer at pH 7.4 (except for coumarin incubations, which were conducted in 50 mM Tris) containing 20 mM glucose 6-phosphate, 2.0 mM NADP, 2.0 mM magnesium chloride, and 2.0 units of glucose 6-phosphate dehydrogenase in a total volume of 500 μl. There was a 2-min preincubation step at 37°C, before the reaction was started by the addition of the specific substrate.
When the substrate was water soluble, the stock solution was prepared in an aqueous solution (water for dextromethorphan and 60 mM potassium hydroxide for chlorzoxazone), and proper organic solvent volumes were added to the incubations to get the desired amount of organic solvent. When the substrate was not water soluble (phenacetin, tolbutamide, coumarin, S-mephenytoin, testosterone), the stock solutions were prepared in a DMSO, acetonitrile, or methanol solution to give a final solvent concentration of 0.2% in the incubation mixture. Then, an appropriate volume of solvent was added to get the desired organic solvent content. For these insoluble substrates, the control incubations containing no organic solvent were done using a methanolic stock solution where the methanol was allowed to evaporate at room temperature. Then, the preincubated oxidative buffer containing the microsomes was added to the tube, and the suspension was mixed by vortexing. It was assumed, unless specified, that the prewarmed microsomal solution would redissolve the compound. Each experiment was conducted in triplicate. After a specific period of time, the reactions were quenched by adding the appropriate chemical to precipitate the proteins as described elsewhere (Chauret et al., 1997). The incubation mixtures were then centrifuged for 10 min at 13,000 rpm in an Eppendorf Centrifuge 5415C. An aliquot of the supernatant fraction (injection volume of 50 μl) was analyzed by HPLC.
HPLC Analysis.
The HPLC system consisted of a Waters 600S controller, a Waters 717 plus Autosampler, a Waters 996 photodiode array detector, and a Shimadzu RF-551 fluorescence HPLC monitor; the data were collected and processed by Millennium version 2.15 software. The HPLC methods for the analysis of the phenacetin-O-deethylase, coumarin-7-hydroxylase, tolbutamide hydroxylase,S-mephenytoin 4′-hydroxylase, dextromethorphan-O-demethylase, chlorzoxazone 6-hydroxylase, and testosterone 6β-hydroxylase activities have been reported previously (Chauret et al., 1997).
Data Analysis.
Results are obtained from three different incubations and are represented as a mean ± standard deviation. In general, there was considered to be a solvent effect when the activity in the incubations containing solvents does not overlap with the activity in the controls, taking into consideration the standard deviation. The extent of inhibition or enhancement in the activity is the ratio of the average activity in the incubations containing organic solvent over the average activity in the controls containing no organic solvent, expressed in percentage.
Results and Discussion
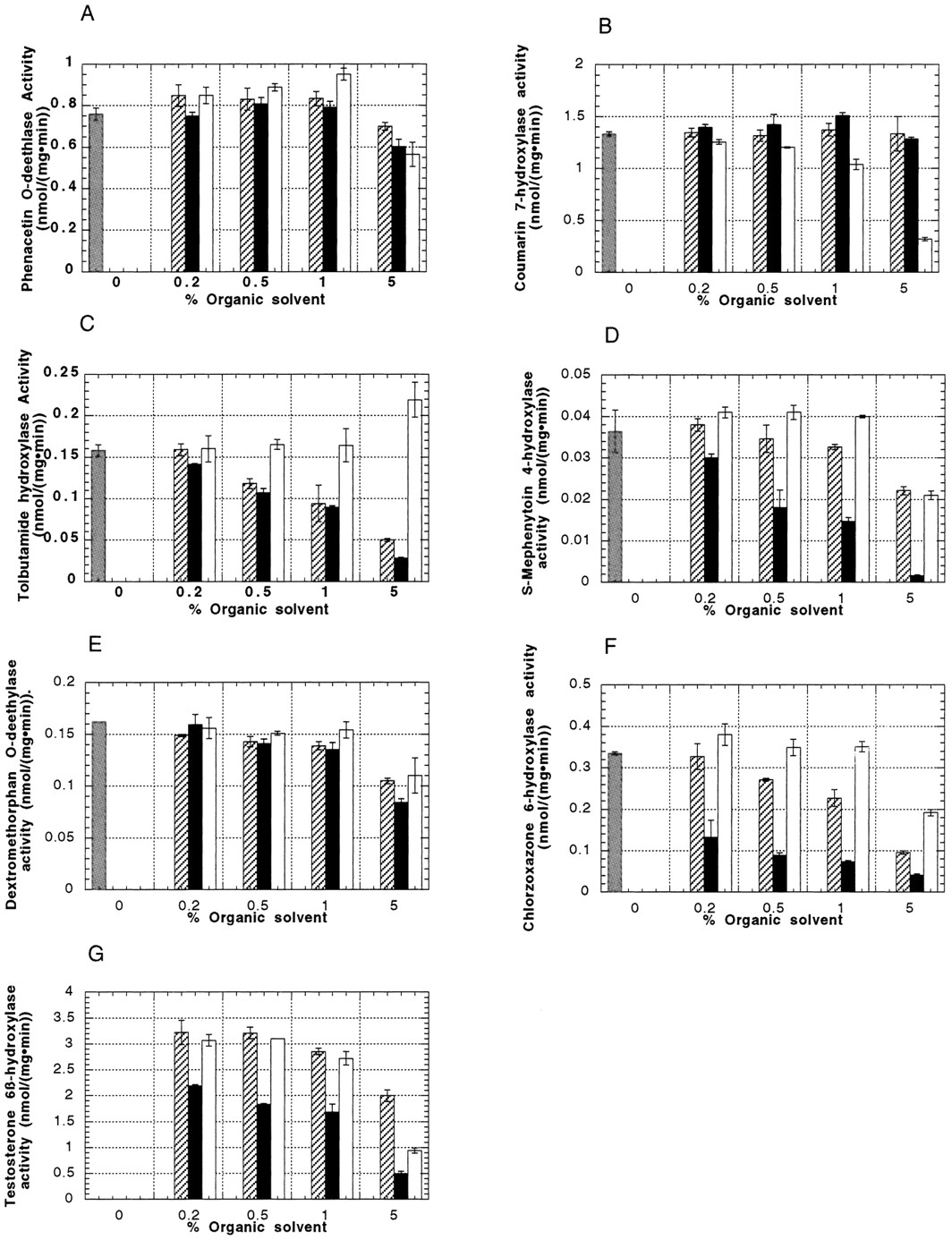
The results of the effect of methanol, DMSO, and acetonitrile on the catalytic activity of seven cytochromes P450 are summarized in fig.1. In all cases, there was no systematic effect of a particular solvent on the various P450s, and the various solvents did not affect the metabolism of a specific probe to the same extent.

Effect of organic solvent on cytochrome P450-mediated activities in human liver microsomes.
A, phenacetin-O-deethylase activity for P4501A2; B, coumarin-7-hydroxylase activity for P4502A6;C, tolbutamide hydroxylase activity for P4502C8/2C9;D, S-mephenytoin-4-hydroxylase activity for P4502C19; E, dextromethorphan-O-demethylase activity for P4502D6;F, chlorzoxazone-6-hydroxylase activity for P4502E1;G, testosterone 6β-hydroxylase activity for P4503A4. Results are an average of N = 3.Gray bars, control; stippled bars, methanol; black bars, DMSO; white bars, acetonitrile.
Overall, the phenacetin-O-deethylase P4501A2 activity (fig.1A) was not affected by methanol or DMSO at levels ≤1%. In the presence of acetonitrile at levels ≤1%, there was a detectable increase in the activity observed. However, at a solvent concentration of 5%, the P4501A2 activity was decreased in all cases (12, 20, and 27% inhibition for methanol, DMSO, and acetonitrile, respectively, as compared with the control incubations).
In general, the coumarin-7-hydroxylase P4502A6 activity (fig.1B) was not changed in the presence of methanol or DMSO in the concentration range studied. This is in line with the findings ofDraper and Parkinson (1997). Inhibition of the activity was observed with increasing content of acetonitrile (5–75% inhibition when increasing the acetonitrile content from 0.2 to 5%) .
The tolbutamide hydroxylase P4502C8/2C9 activity (fig. 1C) was not noticeably affected in the presence of 0.2% of methanol, DMSO, or acetonitrile. However, at concentrations ≥0.5%, the activity was strongly inhibited in the presence of methanol and DMSO. When acetonitrile was present at levels up to 1%, there was no effect on the P4502C8/9 activity, but surprisingly, the activity was enhanced in the presence of 5% acetonitrile (139% of control). To confirm this finding, another P4502C9 substrate, diclofenac, was used, and an increase of activity (125% of control) was also observed in presence of 5% acetonitrile. There are other literature examples reported for the activation of monooxygenase reactions in liver microsomes by chemicals (Kawalek and Andrews, 1980). For example, acetone is known to stimulate the oxidation of acetaminophen (Moldéus and Gergely, 1980).
The S-mephenytoin 4-hydroxylase P4502C19 activity (fig.1D) was not noticeably affected in the presence of ≤1% methanol or acetonitrile. However, it was inhibited in the presence of DMSO even at a low concentrations. At a 5% level of organic solvent, the activity was attenuated to varying degrees (39, 95, and 42% inhibition for methanol, DMSO, and acetonitrile, respectively).
Dextromethorphan-O-demethylase P4502D6 activity (fig.1E) was not changed in the presence of ≤1% acetonitrile. However, it was slightly diminished in the presence of ≤1% methanol or DMSO. At a concentration of 5%, all three solvents significantly reduced the catalytic activity of P4502D6 (35, 49, and 35% inhibition for methanol, DMSO, and acetonitrile).
Chlorzoxazone 6-hydroxylase P4502E1 activity (fig. 1F) was slightly increased by 0.2% acetonitrile (13% enhancement as compared with control) and significantly inhibited at 5% acetonitrile (40% inhibition). Methanol did not affect the P4502E1 activity at 0.2%, but inhibition of the activity was seen at concentrations ≥0.5%. Finally, DMSO strongly inhibited the activity, even at levels as low as 0.2% (60% inhibition). This is consistent with the fact that DMSO is a P4502E1 inhibitor (Yoo et al., 1987).
Testosterone 6β-hydroxylase P4503A4 activity (fig. 1G) could not be evaluated in the absence of organic solvent. The results obtained with the control incubations containing no organic solvent were irreproducible and indicated an extent of metabolism that was very low [<0.5 nmol/(mg·min)]. A possible explanation is that testosterone does not resolubilize upon addition of the preincubated mixture containing the microsomes and the incubation buffer. Based on the results obtained with 0.2% methanol (highest activity seen), it seems that the P4503A4 activity was not significantly modified by the presence of methanol and acetonitrile when present at levels ≤0.5%. However, the presence of DMSO, even at 0.2%, strongly reduced the activity (32% inhibition). In all cases, the 5% organic solvent content strongly diminished the activity (30, 85, and 71% inhibition for methanol, DMSO, and acetonitrile, respectively).
As demonstrated here and by others, the presence of organic solvents can greatly affect the in vitro metabolism of a variety of enzymatic activities (Kawalek and Andrews, 1980; Yoo et al., 1987; Draper and Parkinson, 1997). Although DMSO is a good universal organic solvent, it is not an optimal solvent because it can inhibit the activity of several P450s (2C19, 3A4, 2E1, and 2C8/9) even at low levels (0.2%). In general, methanol and acetonitrile represent better alternatives as long as the content is kept at a relatively low level. It was found that the content should be kept lower than 1%, as, in general, with all the three solvents studied, a concentration of 5% strongly affected the metabolism of the various probes studied. It is important to mention that the effects observed could vary with the experimental conditions such as the protein content in the microsomal incubations, especially if the inhibition is due to competitive metabolism of the solvents.
The presence of an organic solvent can strongly affect the reliability and interpretation of in vitro data. For example, if the solvent selected for in vitro studies inhibits particular P450s, one might miss an important metabolic pathway of a new compound. Also, if the rate of metabolism is affected by the solvent, the kinetic data generated by in vitro studies will be erroneous and prediction of in vivo intrinsic clearance will not be accurate. If, for an inhibition study, the organic solvents used to solubilize the substrate and the inhibitor are not kept minimal and constant, this may result in a nonspecific and/or a nonsensitive inhibition assay. This study has shown that the selection and amount of solvent for effective solubilization of chemicals is a critical parameter to consider when designing in vitro metabolism studies of new chemical entities.
Footnotes
-
Send reprint requests to: Nathalie Chauret, Merck Frosst Centre for Therapeutic Research, P.O. Box 1005, Pointe-Claire Dorval, Quebec H9R 4P8, Canada.
- Abbreviations used are::
- P450
- cytochrome P450
- DMSO
- dimethyl sulfoxide
- Received August 28, 1997.
- Accepted October 14, 1997.
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}