


Abstract
Famciclovir, a 9-substituted guanine derivative, is a new antiviral agent which undergoes rapid hydrolysis and oxidation in man to yield the active antiherpes agent, penciclovir. Studies with human liver cytosol have indicated that the oxidation of the penultimate metabolite, 6-deoxypenciclovir, to penciclovir is catalyzed by the molybdenum hydroxylase, aldehyde oxidase. In the present study the oxidation of famciclovir and 6-deoxypenciclovir with partially purified molybdenum hydroxylases from human, guinea pig, rabbit, and rat livers and bovine milk xanthine oxidase has been investigated. Famciclovir and 6-deoxypenciclovir were oxidized predominantly to 6-oxo-famciclovir and penciclovir, respectively, by human, guinea pig, and rat liver aldehyde oxidase. Small amounts of 8-oxo and 6,8-dioxo-metabolites were also formed from each substrate. Famciclovir and 6-deoxypenciclovir were good substrates for rabbit liver aldehyde oxidase but, in each case, two major metabolites were formed. 6-Deoxypenciclovir was converted to penciclovir and 8-oxo-6-deoxypenciclovir in approximately equal quantities; famciclovir was oxidized to 6-oxo-famciclovir and a second metabolite which, on the basis of chromatographic and UV spectral data, was thought to be 8-oxo-famciclovir. Two groups of Sprague Dawley rats were identified; those containing hepatic aldehyde oxidase and xanthine oxidase and those with only xanthine oxidase. These have been designated AO-active and AO-inactive rats, respectively. Famciclovir was not oxidized by enzyme from AO-inactive rats or bovine milk xanthine oxidase although 6-deoxypenciclovir was slowly converted to penciclovir by rat liver or milk xanthine oxidase. Inhibitor studies showed in human, guinea pig, and rabbit liver that xanthine oxidase did not contribute to the oxidation of famciclovir and 6-deoxypenciclovir; thus it is proposed that drug activation in vivo would be catalyzed solely by aldehyde oxidase.
Famciclovir 2-[2-(2-Amino-9H-purin-9-yl) ethyl]-1,3-propanediol diacetate (ester), a recently introduced drug, is a synthetic guanine derivative which is metabolized to the potent antiviral compound penciclovir. Penciclovir is active against herpes simplex virus types 1 and 2, varicella zoster virus, Epstein-Barr virus, and hepatitis B (1). Like aciclovir, another 9-substituted guanine derivative, penciclovir is selectively phosphorylated in virus-infected cells to a monophosphate ester by thymidine kinase, followed by further phosphorylation to a triphosphate ester which inhibits virus DNA polymerases (2). Compared with aciclovir, penciclovir administration leads to higher triphosphate ester concentrations in virus-infected cells and its antiviral activity persists for a longer time after removal of the compound (2, 3). Famciclovir is absorbed rapidly and extensively after oral administration, and total systemic availability of penciclovir is 77% (4), which is about four times higher than that of aciclovir (5).
Metabolism of famciclovir involves sequential hydrolysis of both acetyl groups to give 6-deoxypenciclovir which is subsequently oxidized to penciclovir (fig. 1) (6). The oxidative step was initially attributed to the action of the molybdenum hydroxylase, xanthine oxidase (xanthine: O2 oxidoreductase EC 1.2.3.2), on the basis of studies performed with 6-deoxyaciclovir and a structural similarity between 6-deoxypenciclovir and guanine which is a substrate of xanthine oxidase (6-8). It was later shown that in human liver cytosol the conversion of 6-deoxypenciclovir to penciclovir was catalyzed by another molybdenum hydroxylase, aldehyde oxidase (aldehyde: O2 oxidoreductase EC 1.2.3.1) (9). However, it is not known whether famciclovir is a substrate for aldehyde oxidase or whether famciclovir or 6-deoxypenciclovir reacts with xanthine oxidase. Indeed, in the study of Clarke et al. (9) the xanthine oxidase activity of human liver cytosol was not monitored, whereas human liver xanthine oxidase may be labile in the enzyme preparation (10). Furthermore, the enzymology of penciclovir formation in other species has not been investigated. It is now well established that the activity of aldehyde oxidase towards different substrates may be species dependent (11-15). Significant species differences have been demonstrated in the biotransformation of other guanine-based antiviral agents, such as 6-deoxyaciclovir (7, 16), 6-deoxycarbovir (17), and BRL 55792 (18). Therefore, the present study investigates the role of aldehyde oxidase and xanthine oxidase in the metabolism of famciclovir and 6-deoxypenciclovir in partially purified human, guinea pig, rat, and rabbit liver fractions.

The major metabolic pathway of famiciclovir in man (6).
Materials and Methods
Chemicals.
Phthalazine and phenanthridine were purchased from Aldrich Chemical Company (Gillingham, Dorset, UK). Leupeptin, dithioerythritol, phenylmethylsulfonylfluoride, menadione, isovanillin, xanthine, and bovine milk xanthine oxidase (Grade I) were obtained from Sigma Chemical Company (Poole, Dorset, UK). Famciclovir and its analogues were supplied by SmithKline Beecham Pharmaceuticals (Great Burgh, Surrey, UK).
Preparation of Molybdenum Hydroxylase Fractions.
Mature male or female Dunkin-Hartley guinea pigs (400–600 g, University of Bradford, Bradford, UK) and Sprague-Dawley rats (200–300 g, Bantin and Kingman, Hull, UK), previously maintained on a standard laboratory diet, were killed between 9 and 10 a.m. by cervical dislocation. Livers were immediately excised, placed in ice-cold isotonic potassium chloride solution (1.15% KCl w/v) containing 0.1 mM EDTA, and the gall bladder and excess fat removed. Partially purified molybdenum hydroxylase fractions were prepared from liver homogenates by heat treatment and ammonium sulfate precipitation according to the method of Beedham et al. (15).
Enzyme fractions were also prepared from the livers of male New Zealand white rabbits (800–1000 g, obtained from Stocks House Farm, Bradford, England) using a similar method to that described for guinea pig and rat livers. Human liver fractions were prepared from livers which had been flash frozen (−80°C) on collection samples coded H30, H32, H28, H33, and H102, obtained from International Institute for the Advancement of Medicine, Exton, PA), using the following modification to the above method:
Each liver sample was weighed, defrosted, and placed in 3- 4 volumes of 20 mM Tris buffer pH 7.4 containing 1 mM EDTA, 1 mM dithioerythritol, 50 μM p-methylsulphonyl fluoride, and 10 μM leupeptin at 4°C and homogenized at 12,000 rpm for 1–2 min, using a Ystral X10/25 homogenizer at 4°C. The resulting homogenate was then heated at 55–57°C for 10 min on a steam bath and treated as for guinea pig fractions. The final EDTA suspension was stored in liquid nitrogen until required.
Spectrophotometric measurement of enzyme activity.
All spectrophotometric determinations were carried out at 37°C using a Pye-Unicam SP8–250 UV/VIS spectrophotometer fitted with a Pye-Unicam cell temperature control unit. Enzyme activity of partially purified fractions was monitored using phenanthridine, phthalazine, and xanthine (19, 20). Oxidation of 1 mM phthalazine was followed at 420 nm using 1 mM potassium ferricyanide as electron acceptor (ε = 2080 M−1cm−1), whereas oxidation of 0.05 mM phenanthridine and 0.05 mM xanthine was monitored at 322 nm (ε = 9000 M−1cm−1) and 295 nm (ε = 11000 M−1cm−1) using molecular oxygen as the electron acceptor, respectively. All reactions were carried out in 67 mM Sorenson’s phosphate buffer pH 7.0 containing 0.1 mM EDTA at 37°C.
The reaction of famciclovir and 6-deoxypenciclovir with guinea pig, rat, rabbit, and human liver fractions or bovine milk xanthine oxidase was also determined in Sorenson’s phosphate buffer pH 7.0 containing 0.1 mM EDTA at 37°C either by repeated scanning of the UV spectra between 200–350 nm up to 30–60 min or by measurement of the initial oxidation rates at 420 nm using potassium ferricyanide as electron acceptor as described for phthalazine. Substrates were tested for non-enzymatic interaction with potassium ferricyanide prior to the addition of enzyme to cuvettes.
Incubations were also performed in the presence of 1–100 μM specific aldehyde oxidase (menadione and isovanillin) and xanthine oxidase (allopurinol) inhibitors (10, 13, 21, 22).
Protein Determination.
Protein concentrations of partially purified enzyme fractions were determined spectrophotometrically using a Pierce BCA Protein assay kit with bovine serum albumin as a protein standard (23).
Incubations with Partially Purified Molybdenum Hydroxylase Fractions.
Famciclovir and 6-deoxypenciclovir (50 μM or 500 μM) were incubated with 0.1 ml partially purified human, guinea pig, rabbit, and rat liver fractions or bovine milk xanthine oxidase at 37°C in a total volume of 3 ml 67 mM Sorenson’s phosphate buffer pH 7.0 containing 0.1 mM EDTA. Incubations were performed in 10-ml closed vials which were placed in a shaking water bath and pre-warmed to 37°C. Aliquots (200 μl) were removed at 1, 5, 10, 15, 30, 45, 60, and 90 min and added to either 200 μl methanol (in the case of famciclovir) or 100 μl 20% trichloroacetic acid (in the case of 6-deoxypenciclovir) to terminate the reaction. Samples were centrifuged in a Beckman (High Wycombe, Bucks, UK) bench-top microcentrifuge for 3–5 min and the supernatants were subsequently analyzed by HPLC.
Incubations were also carried out in the presence of 1–100 μM of menadione, isovanillin, and allopurinol.
HPLC Analysis of Famciclovir and 6-Deoxypenciclovir Oxidation.
HPLC analysis was carried out using a system supplied by Waters Associates (Northwich, Cheshire, UK) which consisted of a 510 pump, 710 B WISP automatic injector, Lambda-Max LC Spectrophotometer, and 740 data module. Chromatographic separation was achieved using a Spherisorb ODS2 5μm (25 cm × 4.6 mm i.d.) column with a μBondapak C18 Guard-Pak insert and 0.5 mM ammonium acetate, pH 4.65/acetonitrile as the mobile phase at a flow rate of 1.5 ml/min. The percentage of the organic modifier was as follows: 5% for the separation of 6-deoxypenciclovir and its oxidation metabolites, 9% for the analyses of famciclovir and all its metabolites, 15% for the metabolism of famciclovir and its oxidation metabolites. UV detection was at 280 nm. Oxidized metabolites were identified by comparison of their HPLC retention times and UV spectra with those of authentic standards. Calibration lines for all compounds were linear in the range of 0.5–600 μM with exception being that the calibration line obtained for famciclovir quantitation with 9% acetonitrile was linear between 10–600 μM.
Determination of Kinetic Constants for the Oxidation of Phenanthridine, Famciclovir and 6-Deoxypenciclovir by Molybdenum Hydroxylases.
UV Determination: KM (Michaelis-Menten constant) and Vmax (maximum initial velocity) values for the oxidation of phenanthridine with human, guinea pig, and AO-active rat liver fractions, and the oxidation of 6-deoxypenciclovir with guinea pig liver fractions and bovine milk xanthine oxidase were determined spectrophotometrically using a method similar to that described by Beedham et al. (20). Kinetic constants were determined using ferricyanide or molecular oxygen as electron acceptor.
HPLC Determination: KM andVmax values for the oxidation of 6-deoxypenciclovir with human, AO-active rat, AO-deficient rat liver fractions, and bovine milk xanthine oxidase (very low oxidation rates), and metabolite formation from the oxidation of famciclovir or 6-deoxypenciclovir catalyzed by rabbit enzyme fractions were determined using HPLC. At least eight different substrate concentrations were used in the range of 0.1 KM to 4–5KM in 67 mM phosphate buffer pH 7.0 containing 0.1 mM EDTA at 37°C in a shaking water bath. The reactions were initiated by the addition of 0.1-ml enzyme fraction, aliquots (200 μl) removed, and protein precipitated at 1 min intervals to determine the linear range of substrate oxidation. The samples were centrifuged for 3–5 min and analyzed by HPLC. To compare the results obtained from HPLC with those determined spectrophotometrically, kinetic constants were also calculated for the oxidation of 6-deoxypenciclovir with guinea pig liver fractions and bovine milk xanthine oxidase using the spectrophotometric method as described above. In both spectrophotometric and HPLC methods, KM andVmax values were determined from a Lineweaver-Burke double reciprocal plot of 1/V against 1/[S]. The line of the best fit through the points on the plot was calculated using linear regression by the least squares method.
Results
Oxidative Activity of Partially Purified Liver Fractions.
Activities of the molybdenum hydroxylase fractions from each species were measured spectrophotometrically using 50 μM phenanthridine and xanthine, specific substrates of aldehyde oxidase (24) and xanthine oxidase (25, 26), respectively, and 1 mM phthalazine which is predominantly oxidized by aldehyde oxidase (19). The ratio of the oxidation rates of phthalazine/phenanthridine/xanthine was calculated to reflect the relative molybdenum hydroxylase activities in the species investigated (table 1).KM values were calculated for the oxidation of phenanthridine by hepatic aldehyde oxidase; these were <1 μM with guinea pig and rabbit liver enzyme and 6 μM and 14 μM with rat and human liver aldehyde oxidase, respectively. KMvalues for xanthine oxidation by xanthine oxidase also range from <1 μM-14 μM (26), whereas the KM values for the oxidation of phthalazine by hepatic aldehyde oxidase are around 50–100 μM (14, 15, 20).
Hepatic xanthine oxidase and aldehyde oxidase activity in different species
Aldehyde oxidase activity towards phenanthridine or phthalazine was highest in rabbit liver fractions (table 1). Molybdenum hydroxylase activities of guinea pig and rabbit liver were comparable with those reported by others under similar experimental conditions (19, 27). Two groups of Sprague-Dawley rats, containing approximately equal numbers, were identified with respect to hepatic molybdenum hydroxylase activity; group A rats (N = 10), which were active towards phenanthridine, phthalazine, and xanthine; and group B rats (N = 15), which catalyzed the oxidation of xanthine but which did not show measurable activity when the oxidation rate of phenanthridine was determined up to 5 min. The enzyme fractions from group B rats were also much less active towards phthalazine (table 1). It would thus appear that Sprague-Dawley rats can be divided into two groups: those containing hepatic aldehyde oxidase and xanthine oxidase activity and those with just xanthine oxidase activity but zero or minimal aldehyde oxidase levels. These have been designated AO-active (group A) and AO-deficient (group B) rats respectively. There was no significant difference between the aldehyde oxidase activity in male and female AO-active rats. The level of xanthine oxidase in AO-deficient rat livers was also lower than that in AO-active rat livers (p < 0.01) which may indicate some deficiency in xanthine oxidase activity in these animals as well. Hepatic xanthine oxidase activity in male and female AO-active rats was almost identical, whereas in AO-deficient rats xanthine oxidase was 72% higher (p< 0.002) in males than in females.
Human liver is thought to contain low aldehyde oxidase levels (12, 16,28); however, all five enzyme fractions obtained from frozen human liver samples showed reasonable activity towards aldehyde oxidase substrates (table 1) although these values were lower than those found with fresh animal liver enzyme. This may be, in part, a result of the lability of human liver aldehyde oxidase as enzyme activity rapidly disappears upon storage in post-mortem tissues and after freezing and thawing of post-mortem samples (28,29). Interestingly, in contrast to other studies (12,28), with all aldehyde oxidase substrates used in this study human liver aldehyde oxidase activity was much higher than that of xanthine oxidase.
Oxidation of famciclovir and 6-deoxypenciclovir was followed initially at 420 nm using 1 mM potassium ferricyanide as an artificial electron acceptor. At 1 mM concentrations, the oxidation rates of famciclovir and 6-deoxypenciclovir were between those obtained for phthalazine and phenanthridine with human, guinea pig, and rabbit liver enzymes, whereas the two substituted purines were oxidized relatively slowly by AO-active rat liver fractions (table 1). In contrast to famciclovir, which did not react with AO-deficient rat enzyme, 6-deoxypenciclovir was slowly oxidized by this fraction.
Spectrophotometric Measurement of 6-Deoxypenciclovir and Famciclovir Oxidation by Partially Purified Liver Molybdenum Hydroxylase Fractions.
6-Deoxypenciclovir (fig. 2) and famciclovir gave identical UV spectra at pH 7.0 with a λmax at 305 nm. The spectra of the possible oxidation products of 6-deoxypenciclovir i.e.penciclovir, 8-oxo-6-deoxypenciclovir, and 8-oxopenciclovir, are shown in fig. 3. With human, guinea pig, and AO-active rat liver fractions, the successive spectrum scans of 6-deoxypenciclovir and famciclovir incubations showed complete reduction of the absorption maximum, at 305 nm, and appearance of a new peak at 252 nm with a shoulder at ≈268 nm (fig. 2). In each case, the final spectrum corresponded to that of the 6-oxo-metabolite, indicating that both 6-deoxypenciclovir and famciclovir are predominantly oxidized at carbon 6 giving penciclovir and 6-oxo-famciclovir, respectively, as the major metabolites. In contrast, no significant change in the spectrum of 6-deoxypenciclovir or famciclovir was observed when either substrate was incubated with AO-deficient rat liver fractions. Similarly, the UV spectrum of 6-deoxypenciclovir did not vary markedly in incubations with bovine milk xanthine oxidase although it was slowly metabolized to penciclovir with high enzyme concentrations. Famciclovir showed no reaction at all even with higher concentrations of bovine milk xanthine oxidase.

Comparison of spectra obtained from the incubation of 100 μM 6-deoxypenciclovir (—–) with guinea pig (— —) and rabbit (- - - -) liver fractions at 37°C in phosphage buffer pH containing 0.1 mM EDTA for 25 min.
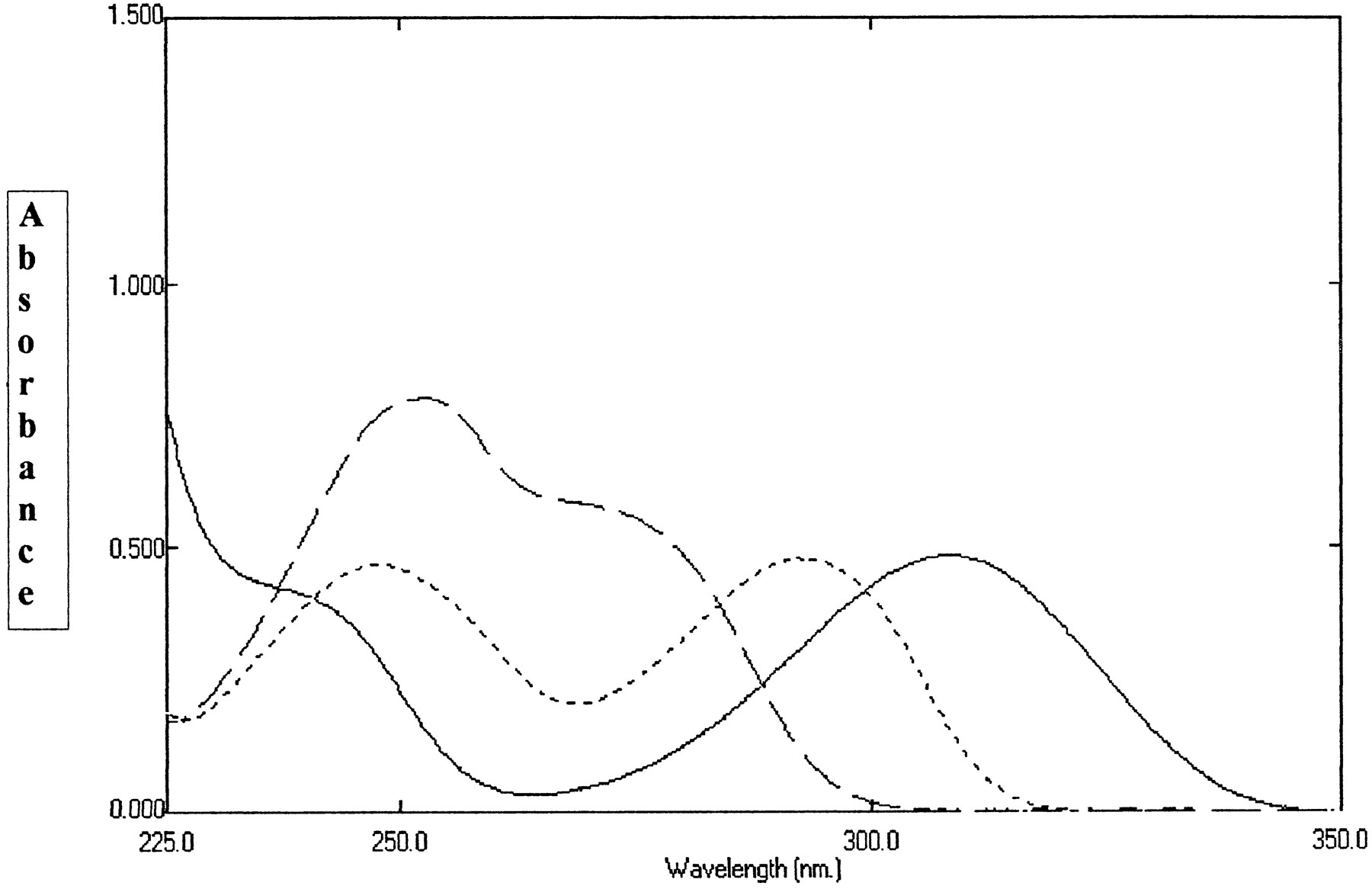
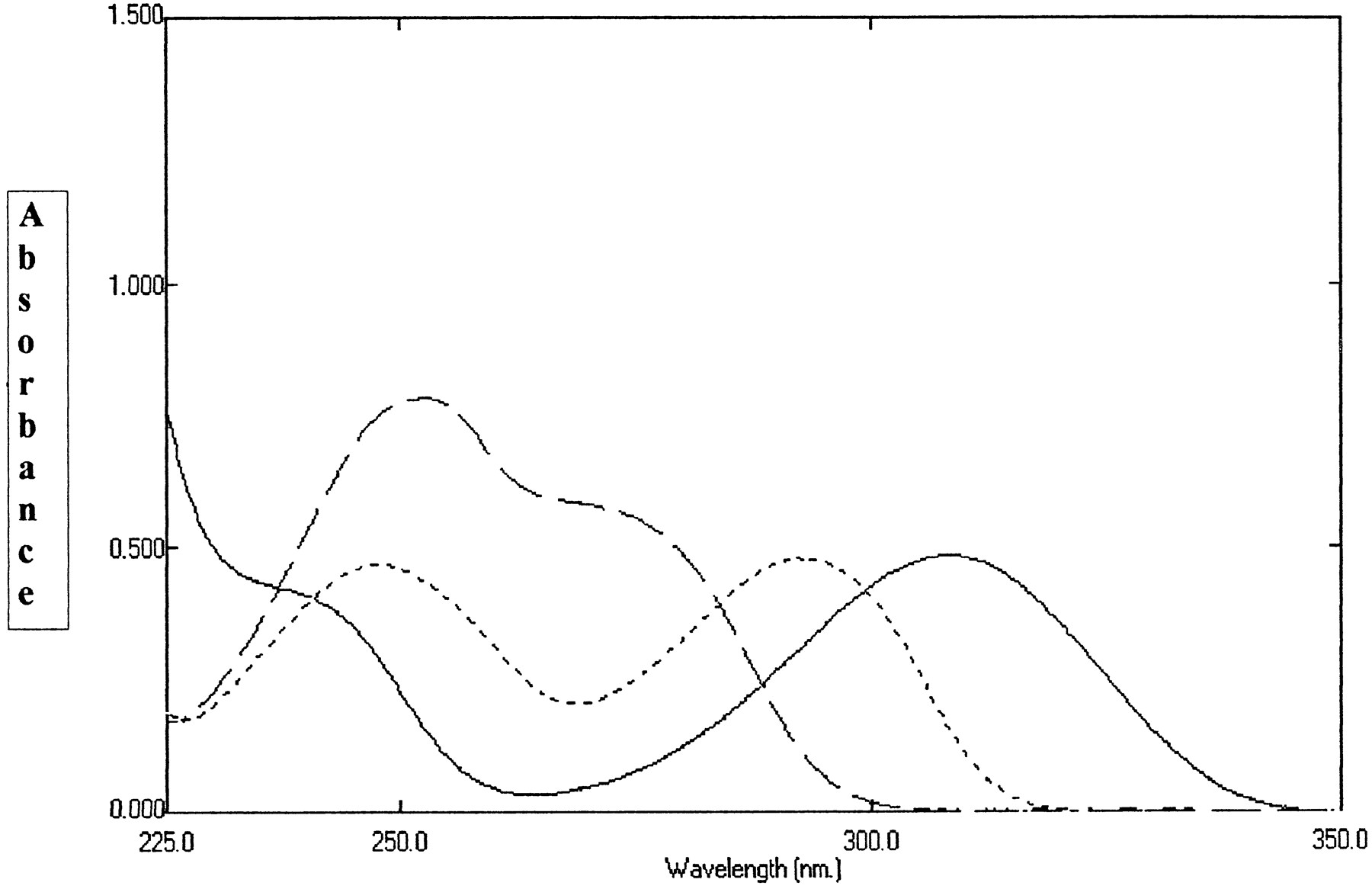
UV spectra of 50 μM 8-oxo-6-deoxypenciclovir (——–), penciclovir (— —) and 8-oxo-penciclovir (- - - - ) measured at 37°C in phosphate buffer pH 7.0 containing 0.1 mM EDTA.
When 6-deoxypenciclovir was incubated with rabbit enzyme, the final UV spectrum did not correspond to that of 6-deoxypenciclovir or penciclovir and the resulting spectrum showed λmax at 246 nm and 310 nm with a shoulder at about 280 nm (fig. 2). The absorption maximum of 8-oxo-6-deoxypenciclovir, an alternative metabolite of 6-deoxypenciclovir, is at 310 nm with a shoulder at ≈240 nm (fig. 3); thus, it would seem that rabbit liver enzyme oxidizes 6-deoxypenciclovir to 8-oxo-6-deoxypenciclovir. However, the final UV spectrum was not identical to that of the 8-oxo-metabolite and the absorbance at ≈310 nm indicated that approximately 50% 6-deoxypenciclovir had been converted to 8-oxo-6-deoxypenciclovir. The absorption at ≈250 nm indicated that penciclovir was also present.
The results obtained from famciclovir incubations with rabbit liver fractions were similar to those obtained from 6-deoxypenciclovir. As the presence of the diacetyl function did not alter the UV chromophore in famciclovir and 6-oxo-famciclovir, it is assumed that 8-oxo-famclovir would have the same UV spectrum as that of 8-oxo-6-deoxypenciclovir. Therefore, by analogy with 6-deoxypenciclovir, it is suggested that famciclovir is oxidized by rabbit liver fractions to 6-oxo-famciclovir and 8-oxo-famciclovir.
HPLC Analysis of the Oxidation of Famciclovir and 6-Deoxypenciclovir by Partially Purified Liver Molybdenum Hydroxylase Fractions.
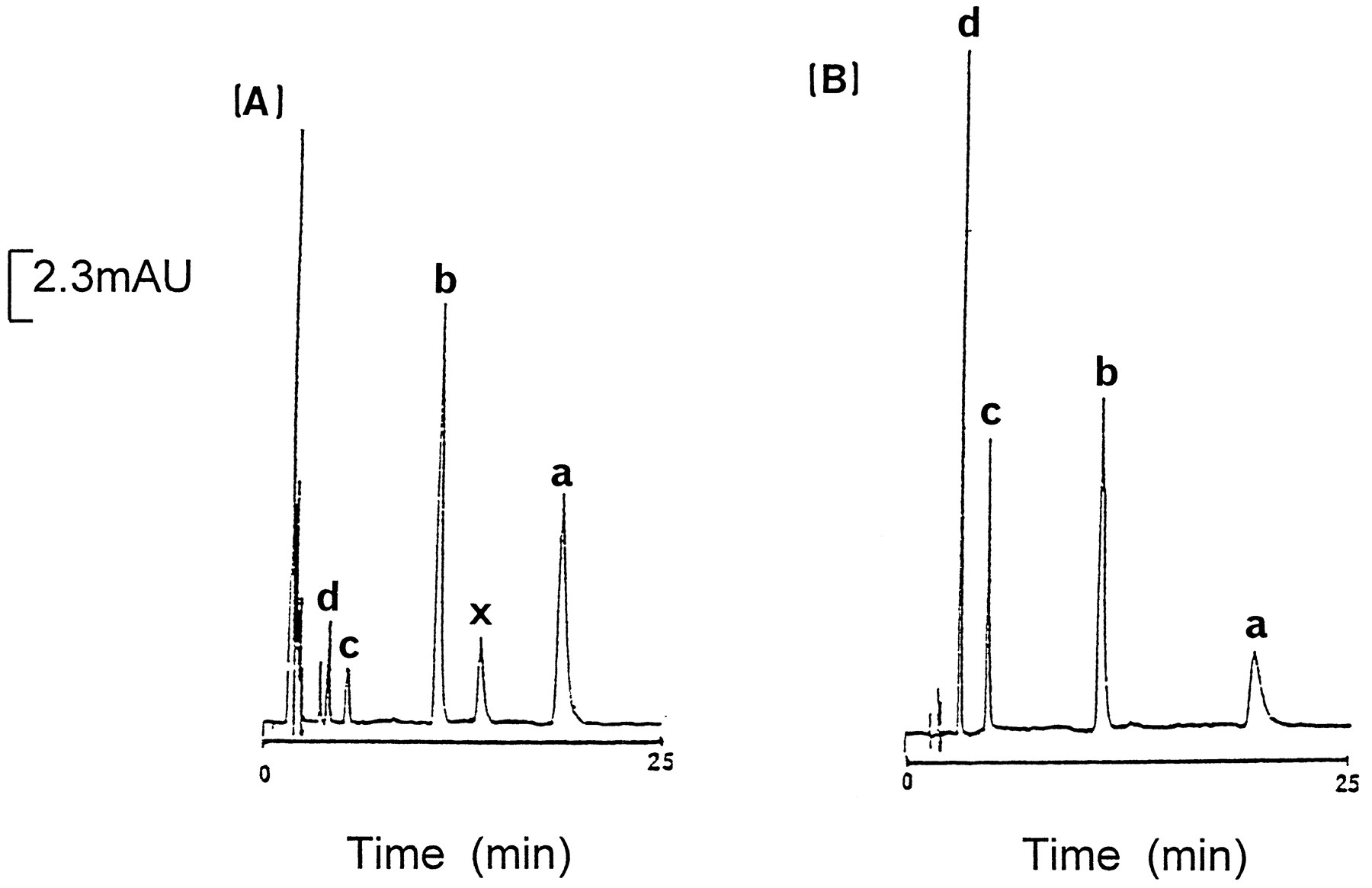
Table 2 summarizes the product formation from the incubation of 6-deoxypenciclovir with hepatic human, guinea pig, rat, and rabbit liver fractions and bovine milk xanthine oxidase after 15 min. 6-Deoxypenciclovir was converted to three metabolites by guinea pig, rabbit, and AO-active rat liver molybdenum hydroxylases. These co-eluted with penciclovir, 8-oxo-6-deoxypenciclovir, and 8-oxo-penciclovir. The major metabolite in 6-deoxypenciclovir incubations with guinea pig and AO-active rat liver fractions was penciclovir with much lower amounts of 8-oxo-6-deoxypenciclovir and 8-oxo-penciclovir produced (table 2). 6-Deoxypenciclovir was also predominantly oxidized by human liver aldehyde oxidase to penciclovir with only low concentrations of 8-oxo-6-deoxypenciclovir detected. In contrast, with rabbit liver aldehyde oxidase, penciclovir and 8-oxo-6-deoxypenciclovir were produced in approximately equal amounts with much lower concentrations of 8-oxo-penciclovir also formed (fig.4). This confirms the results obtained from the spectral analysis of 6-deoxypenciclovir oxidation by rabbit aldehyde oxidase. The only product formed in 6-deoxypenciclovir incubations with AO-deficient rat fractions or bovine milk xanthine oxidase was penciclovir (table 2).
Products obtained after incubation of 500 μM 6-deoxypenciclovir2-a with partially purified human, guinea pig, rabbit, and rat liver molybdenum hydroxylases and bovine milk xanthine oxidase for 15 min2-b

(A) HPLC-UV chromatogram from the incubation of 500 μM 6-deoxypenciclovir with rabbit liver aldehyde oxidase for 15 min. (B) The HPLC separation of 6-deoxypenciclovir (a), 8-oxo-6-deoxypenciclovir (b), 8-oxo-penciclovir (c), and penciclovir (d) under the same chromatographic conditions (see Methods).
Penciclovir was refractory to further oxidation when incubated with guinea pig, rabbit, or AO-active rat enzyme although 8-oxo-6-deoxypenciclovir was slowly oxidized to 8-oxo-penciclovir by guinea pig, rabbit, and AO-active rat liver enzyme fractions. Therefore, it seems that the 8-oxo-penciclovir detected in 6-deoxypenciclovir incubations arises from further oxidation of 8-oxo-6-deoxypenciclovir and not penciclovir.
Chromatographic analysis showed that famciclovir predominantly underwent oxidation to 6-oxo-famciclovir by guinea pig and AO-active rat fractions. Small amounts of mono-desacetylfamciclovir, the initial de-esterified metabolite of famciclovir, and its oxidation product, 6-oxo-desacetylfamciclovir, were also produced with both enzyme fractions. In addition to 6-oxo-famciclovir, a second major peak (Metabolite X) with a RT of 13.7 min was also formed from famciclovir with rabbit enzyme fractions (fig. 5). Although authentic 8- and 6,8-oxo-metabolites of famciclovir were not available, by comparison with the relative retention times and UV-spectral analysis of 6-deoxypenciclovir and famciclovir metabolites, the unknown metabolite generated in rabbit incubations is thought to be 8-oxo-famciclovir.
(A) HPLC-UV chromatogram from the incubation of 500 μM famciclovir with rabbit liver aldehyde oxidase for 15 min. (B) The HPLC separation of famiclovir (a), 6-oxofamciclovir (b), desacetylfamciclovir (c), and 6-oxo-desacetylfamciclovir (d) under the same chromatographic conditions (see Methods).
Very low concentrations of 6-oxo-famciclovir were produced after prolonged (>90 min) incubation of famciclovir with AO-deficient rat liver fractions, which probably arises from residual aldehyde oxidase activity in the enzyme fractions.
Inhibition of 6-Deoxypenciclovir and Famciclovir Oxidation.
The relative contribution of aldehyde oxidase and xanthine oxidase to the oxidation of 6-deoxypenciclovir and famciclovir was determined using specific enzyme inhibitors. The effects of the inhibitors on the initial rates of substrate oxidation are summarized in table3. Menadione (1 μM), which is a potent aldehyde oxidase inhibitor but an electron acceptor of xanthine oxidase (21), caused significant inhibition of 6-deoxypenciclovir and famciclovir oxidation by human, guinea pig, and rabbit liver enzyme (table 3). Higher menadione concentrations (10 and 100 μM) reduced initial oxidation rates by 90–100%. In contrast, 100 μM allopurinol, a mechanism-based inhibitor of xanthine oxidase (22), had little effect with any of the enzyme fractions. Isovanillin, a reversible inhibitor of aldehyde oxidase (10,30), also caused a significant decrease in the oxidation of 6-deoxypenciclovir and famciclovir in guinea pig, human,and AO-active rat incubations (table 3); however, it was found to be a substrate of rabbit liver aldehyde oxidase undergoing oxidation to isovanillic acid (KM = 0.045 mM,Vmax = 211 nmol/mg/ml, N = 2,r = 0.999). HPLC analysis showed that the formation of all metabolites from 6-deoxypenciclovir, by human, guinea pig, and rabbit fractions was significantly inhibited by the aldehyde oxidase inhibitors, and minimal reduction was observed with allopurinol. In addition, the production of 6-oxo-, 8-oxo-famciclovir, and 6-oxo-desacetylfamciclovir was completely inhibited by 100 μM menadione and the reactions were not affected by 100 μM allopurinol. Therefore, aldehyde oxidase seems to be the major molybdenum hydroxylase involved in the oxidation of 6-deoxypenciclovir and famciclovir in human, guinea pig, and rabbit liver.
Effects of aldehyde oxidase and xanthine oxidase inhibitors on the oxidation of 50 μM 6-deoxypenciclovir and famciclovir with partially purified human, guinea pig, rabbit, and AO-active rat liver molybdenum hydroxylases monitored at 420 nm 3-a
Famciclovir oxidation by enzyme from AO-active rats was also significantly reduced in the presence of menadione (table 3), but the oxidation of 6-deoxypenciclovir was only inhibited by 43%, 56%, and 75% in the presence of 1, 10, and 100 μM menadione, respectively. Likewise, when metabolite production was monitored by HPLC, the formation of 6-oxo-, 8-oxo-famciclovir, and 6-oxo-desacetylfamciclovir from famciclovir was almost completely abolished by 100 μM menadione. However, formation of penciclovir from 6-deoxypenciclovir was only decreased by 60%, 66%, and 23% in the presence of 100 μM menadione, isovanillin, or allopurinol, respectively (table4). Metabolism of 6-deoxypenciclovir was completely inhibited by a combination of menadione (100 μM) and allopurinol (100 μM) (tables 3 and 4). In contrast, aldehyde oxidase inhibitors completely reduced the formation of the minor metabolites, 8-oxo-6-deoxypenciclovir and 8-oxo-penciclovir, from 6-deoxypenciclvor, whereas allopurinol had no effect. Therefore, in AO-active rat liver fractions, 8-oxo-6-deoxypenciclovir and 8-oxo-penciclovir are produced solely by aldehyde oxidase, while both aldehyde oxidase and xanthine oxidase participate in the formation of penciclovir from 6-deoxypenciclovir.
Effects of aldehyde oxidase and xanthine oxidase inhibitors on the formation of 6-oxo-metabolites from 500 μM 6-deoxypenciclovir and famciclovir with partially purified rat liver molybdenum hydroxylases
In contrast to the other species tested, the slow oxidation of 6-deoxypenciclovir by AO-deficient rats was markedly inhibited in the presence of allopurinol, whereas menadione and isovanillin had little effect (table 4). Similarly, the oxidation of 6-deoxypenciclovir catalyzed by bovine milk xanthine oxidase was inhibited by 100% and 5% in the presence of 100 μM allopurinol and isovanillin, respectively, whereas 100 μM menadione caused 114% increase in the oxidation rate. 6-Deoxypenciclovir oxidation in AO-deficient rats is thus thought to be catalyzed by xanthine oxidase.
Kinetic parameters for the oxidation of 6-deoxypenciclovir and famciclovir with partially purified human, guinea pig, rabbit, and rat molybdenum hydroxylases and bovine milk xanthine oxidase.
KM and Vmax values for the formation of 6-oxo-metabolites, measured by HPLC, by human, guinea pig, rabbit, and rat enzyme fractions and bovine milk xanthine oxidase are tabulated in table 5. The KMand Vmax values for the oxidation of 6-deoxypenciclovir with partially purified guinea pig liver aldehyde oxidase and bovine milk xanthine oxidase were also determined spectrophotometrically as described in Methods. The kinetic parameters obtained were: KM = 0.37 ± 0.07 mM and Vmax = 158 ± 52 nmol/mg/min (Mean ± SD, N = 3) for guinea pig liver aldehyde oxidase and KM = 1.04 mM andVmax = 131 nmol/mg/min for bovine milk xanthine oxidase. No difference was observed using either potassium ferricyanide or molecular oxygen as an electron acceptor.
Kinetic parameters for 6-oxo-metabolite formation by partially purified guinea pig, human, rabbit, rat liver molybdenum hydroxylases and bovine milk xanthine oxidase determined by HPLC
Famciclovir was a more efficient substrate than 6-deoxypenciclovir with human, guinea pig, and AO-active rat liver enzymes both in terms of enzyme affinity and higher oxidation rates (table 5). SimilarKM values were obtained for the oxidation of 6-deoxypenciclovir with human, guinea pig, rabbit, and AO-active rat liver aldehyde oxidase, although guinea pig enzyme gave the highestVmax/KM value for penciclovir formation. As 6-deoxypenciclovir was converted to two major metabolites, 8-oxo-6-deoxypenciclovir and penciclovir, by rabbit liver aldehyde oxidase, the kinetic values for the formation of 8-oxo-6-deoxypenciclovir was also determined by HPLC. AKM value of 0.32 μM andVmax of 91 nmol/min/mg were calculated (N = 2), which were very similar to those found for penciclovir formation (table 5).
6-Deoxypenciclovir gave high KM andVmax values with bovine milk xanthine oxidase. Although the overall efficiency of bovine milk xanthine oxidase towards 6-deoxypenciclovir seems to be almost as high as that of guinea pig or rabbit aldehyde oxidase (Vmax/KM = 0.172 ml/min/mg), the milk xanthine oxidase was of a much higher purity than the hepatic enzymes. Furthermore, very high xanthine oxidase concentrations had to be used to produce measurable reaction rates with 6-deoxypenciclovir. Compared with xanthine (26), 6-deoxypenciclovir is ≈800-times less efficient as a substrate for bovine milk xanthine oxidase.
Discussion
As previously reported (10, 14, 20), high aldehyde oxidase activity has been detected in guinea pig and rabbit liver with lower, but nevertheless significant, aldehyde oxidase levels in rat and human liver. With all enzyme fractions, aldehyde oxidase-catalyzed oxidation of phthalazine was faster than that of phenanthridine, and in guinea pig, rabbit, and AO-active rat liver fractions aldehyde oxidase activity was much higher than that of xanthine oxidase (table 1). In contrast, 60% of Sprague-Dawley rats were devoid of hepatic aldehyde oxidase. Other evidence indicates marked variation (12, 31-33) and/or a lack (14, 31, 32) of aldehyde oxidase activity in Sprague-Dawley rats using N1-methylnicotinamide (31, 32), pyridoxal (31), 6-methylpurine (12), phenazine methosulphate (33), phthalazine, and carbazeran (14) as substrates. Three groups of Sprague Dawley rats were identified by Stanulovic and Chaykin (31), using pyridoxal and N1-methylnicotinamide as aldehyde oxidase substrates, which were classified as high-activity, intermediate-activity, and zero-activity animals. A structural gene alteration has been suggested for the variation of aldehyde oxidase activity in this group of animals (31).
This study has shown that famciclovir and 6-deoxypenciclovir are extensively oxidized by guinea pig, human, rabbit, and AO-active rat liver aldehyde oxidase and that, in each case, famciclovir is a more efficient substrate than its desacetyl metabolite. This may be a result of the higher lipophilicity of famciclovir, which is a trend that has been noted previously for aldehyde oxidase substrates (13, 15, 24). In contrast, famciclovir did not react with xanthine oxidase whereas 6-deoxypenciclovir was a relatively poor substrate for this enzyme compared with xanthine (26). However, a major difference between rabbit and guinea pig or human aldehyde oxidase is emphasized again by these results. Oxidation of famciclovir and 6-deoxypenciclovir by human and guinea pig liver aldehyde oxidase occurred predominantly at carbon 6 whereas incubations with rabbit liver enzyme produced approximately equal quantities of 8-oxo- and 6-oxo-metabolites. Both pathways were sensitive to inhibition by menadione and thus attributable to aldehyde oxidase although it is possible that different isozymes could be responsible for the formation of each metabolite. This appears to be unlikely as the KM andVmax values calculated for the formation of 8-oxo-6-deoxypenciclovir and penciclovir by rabbit liver enzyme were very similar. Not only does this point to the involvement of a single isozyme form but also indicates that both oxidation reactions occur at the same active site. Whether the substrates bind in two different orientations or the binding site is sufficiently flexible to facilitate nucleophilic attack at carbons 6 and 8 in the purine ring is not clear from the present study. Simultaneous formation of two metabolites from aldehyde oxidase substrates has been observed previously both with quaternary heterocycles such as N1-methylnicotinamide orN-methylquinolinium salts (31, 32, 34) and other purine-based antiviral agents (7, 17). These workers also showed that the ratio of the two oxidation products obtained with rabbit liver aldehyde oxidase differed significantly from that of other species such as guinea pig and man. Rabbit liver aldehyde oxidase is therefore a poor model for human liver enzyme activity (15). figs. 6and 7 summarize the oxidation of famciclovir and 6-deoxypenciclovir with human, guinea pig, and rabbit aldehyde oxidase.

The oxidative metabolic pathways of famciclovir and 6-deoxypenciclovir catalyzed by partially purified human and guinea pig aldehyde oxidase.

The oxidative metabolic pathways of famciclovir and 6-deoxypenciclovir catalyzed by partially purified rabbit liver aldehyde oxidase.
The enzymology of purine oxidation in Sprague Dawley rat liver was different for each substrate and dependent on the aldehyde oxidase status of the liver. On the basis of the results with aldehyde oxidase inhibitors, rat liver aldehyde oxidase is thought to be responsible for famciclovir oxidation. However, oxidation of 6-deoxypenciclovir appears to be catalyzed by both aldehyde oxidase and xanthine oxidase; aldehyde oxidase has the predominant role in AO-active rat liver whereas, in the absence of aldehyde oxidase, in AO-deficient rats 6-deoxypenciclovir is slowly oxidized by xanthine oxidase. As Sprague-Dawley rats are frequently used in metabolism/toxicity studies during drug development programs (16-18), random selection of rats would, at the least, lead to highly variable results. Typing of rat liver with a test aldehyde oxidase substrate would thus be advantageous. It is likely that the contradictory findings observed in some studies (18) arise from an absence of aldehyde oxidase activity in some rats rather than the nonparticipation of aldehyde oxidase in a particular reaction.
Although allopurinol caused a slight decrease in 6-deoxypenciclovir oxidation in guinea pig, human, and rabbit incubations (table 3), it is unlikely that this is a consequence of xanthine oxidase inhibition but that allopurinol is acting as a competitive substrate of aldehyde oxidase (35). Furthermore, it is proposed that xanthine oxidase would not contribute to the oxidation of either 6-deoxypenciclovir or famciclovir in vivo in guinea pig, rabbit, or humans. Thus co-administration of allopurinol with famciclovir to healthy volunteers did not significantly alter penciclovir formation (36).
Human aldehyde oxidase had a higher catalytic activity towards famciclovir than 6-deoxypenciclovir; however, it is unlikely that famciclovir reacts with hepatic aldehyde oxidase in vivo as it is rapidly deacetylated in the intestine and blood to 6-deoxypenciclovir (1). Plasma metabolites, after oral administration of 125–750 mg famciclovir, consist almost entirely of penciclovir with much lower amounts of 6-deoxypenciclovir but no 6-oxo-famciclovir (1,4). 6-Deoxypenciclovir is thus thought to be the in vivosubstrate of aldehyde oxidase. Furthermore, the major famciclovir metabolite, penciclovir, does not appear to undergo further oxidationin vivo (1-4). This correlates well with our in vitro results which showed that penciclovir was not converted to 8-oxo-penciclovir by guinea pig, rabbit, or rat liver aldehyde oxidase although 8-oxo-6-deoxypenciclovir was slowly converted to the 6,8-dioxo-metabolite. Peak plasma levels of penciclovir are formed within 1 hr of famciclovir administration with very little parent drug detected in plasma (4). Oxidation of 6-deoxypenciclovir in vivo thus occurs very quickly, which indicates that aldehyde oxidase activity in human liver is very high. This is in contrast to the findings of in vitro studies (12, 28) and has important implications for other drugs and xenobiotics, particularly those that are better substrates for aldehyde oxidase than 6-deoxypenciclovir. This compound has a relatively low oxidation rate and highKM value compared with other heterocycles or aldehydes. Under similar experimental conditions the substrate efficiency (Vmax/KMvalue) for the oxidation of 6-deoxypenciclovir by human liver aldehyde oxidase is some 10- to 200-fold lower than those of other substrates such as famciclovir, phenanthridine, phthalazine, carbazeran, quinazoline, or vanillin (10, 14, 15). Nevertheless, rapid turnover of 6-deoxypenciclovir occurs in man (1, 4). Similar results were obtained with the inotropic agent, carbazeran, which was rapidly inactivated in man via oxidation by hepatic aldehyde oxidase (37, 38). This further illustrates the high metabolic capacity of human liver aldehyde oxidase (39).
Footnotes
-
Send reprint requests to: Dr. C. Beedham, Pharmaceutical Chemistry, School of Pharmacy, University of Bradford, Bradford, BD7 1 DP, UK.
-
↵1 Current address: Pharmaceutical Chemistry Department, Tabriz School of Pharmacy, Tabriz Medical University of Sciences, Iran.
-
These results were presented in part at a meeting of the British Pharmacological Society, Manchester, UK (1994) and appeared in abstract form in Br. J. Clin Pharmacol. 38, 16P (1994).
-
M.R.R. was supported by a grant from the government of the Islamic Republic of Iran and an ORS award administered by the CVCP.
- Received November 27, 1996.
- Accepted March 17, 1997.
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}