


Abstract
It was recently shown by others that the clearance of midazolam/kg body weight after iv administration correlates with hepatic cytochrome P450 (CYP or P450) 3A content in liver transplant patients. However, after po administration midazolam undergoes significant first-pass metabolism, with significant intestinal extraction. The relationship between hepatic CYP3A and midazolam disposition after po administration had not previously been investigated. The aim of this study was to compare intraindividually hepatic CYP3A content and activity with thein vivo pharmacokinetics of midazolam (7.5 mg) administered po. For 15 patients scheduled for partial liver resection, the AUC values for the observed time period (AUC0–5hr) and to infinity (AUCinf) and the clearance were determined. In a macroscopically normal area of resected liver tissue, the microsomal CYP3A4 content (nanomoles per nanomole of total P450) was measured by immunoblot analysis and parameters (apparentVmax, apparentKM , and intrinsic clearance) for the microsomal α-hydroxylation of midazolam were determined. Clearance/kgin vivo correlated with the apparentVmax (r2 = 0.45, p < 0.01) and the CYP3A4 content (r2 = 0.29, p < 0.05). We conclude that interindividual variability in the pharmacokinetics of po administered midazolam is in part determined by interindividual variability in the hepatic microsomalVmax for the α-hydroxylation of midazolam. However, the relationship between the disposition of midazolam administered po and hepatic CYP3A content is weaker than that reported after iv administration, indicating the importance of the contribution of intestinal CYP3A to the in vivo disposition of midazolam administered po.
CYP3A1has been identified as the enzyme responsible for the metabolism of midazolam (Fabre et al., 1988; Kronbachet al., 1989), with α-hydroxymidazolam being the most important metabolite in humans. Several lines of evidence suggest that CYP3A activity influences the pharmacokinetics of midazolam in vivo. Coadministration of midazolam with other CYP3A substrates (Backman et al., 1994; Hiller et al., 1990) or CYP3A inhibitors (Olkkola et al., 1994, 1996; Fee et al., 1987; Kupferschmidt et al., 1995) has been shown to increase the plasma levels of midazolam. Conversely, CYP3A induction by rifampicin decreased the plasma levels of midazolam (Backmanet al., 1996). In a recent in vitro/in vivo comparison, Thummel et al. (1994) demonstrated a correlation between hepatic microsomal CYP3A content and midazolam clearance/kg body weight after iv administration among liver transplant recipients.
However, after po administration midazolam is exposed to intestinal CYP3A (McKinnon et al., 1995; Kolars et al., 1992) as well as hepatic CYP3A, and recent studies estimated the intestinal extraction ratio of po administered midazolam to be about 0.43 (Thummel et al., 1996; Paine et al., 1996). Because the calculated hepatic and intestinal extraction ratios of midazolam reported by Thummel et al. (1996) for individual subjects were not correlated with each other, the hepatic and intestinal CYP3A activities do not seem to be co-regulated. Thus, it is uncertain whether the significant relationship between hepatic microsomal CYP3A content and the clearance of midazolam demonstrated after iv administration would be maintained for midazolam administered po. In this study we compared the CYP3A content and the midazolam kinetics in hepatic microsomes with the in vivopharmacokinetics of midazolam after po administration to patients scheduled for partial liver resection.
Patients and Methods
Patients.
All patients gave their written informed consent for participation in this study, which was approved by the local ethics committee (University of Heidelberg, Heidelberg, Germany). The subjects were patients scheduled for partial liver resection. Patients receiving any preoperative medication were excluded, to avoid any inhibitory or inducing effects on CYP3A activity produced by concurrent medication. The intake of grapefruit juice, which increases the plasma levels of midazolam (Kupferschmidt et al., 1995), was excluded (as determined by questionnaire) for the last 24 hr before midazolam administration.
Protocol.
On the evening before surgery, the patients received 7.5 mg of midazolam (Dormicum; Hoffmann-LaRoche AG, Grenzach-Wyhlen, Germany) po, after a 4-hr fasting period. Blood samples for measurement of midazolam were obtained before midazolam administration and 20, 40, 60, 90, 120, 180, and 300 min after administration. The samples were centrifuged immediately, and the plasma was frozen at −18°C until analyzed.
Anesthesia was induced with atracurium, fentanyl (100–150 μg), and propofol (2–2.5 mg/kg body weight) and maintained with isoflurane (0.5–1.2%, v/v), N2O (66%, v/v), fentanyl (500–750 μg until removal of the liver tissue, and thereafter as needed), and atracurium as needed. If necessary, atropine, nitroglycerin, or noradrenaline was administered to maintain the heart rate and blood pressure within a range of 85–115% (referring to preoperative values determined with the patients in the supine position and at rest). The capillary O2-saturated hemoglobin concentration was maintained above 96% of the total hemoglobin. Warm ischemia of the resected liver section lasted between 30 and 40 min. After removal of the resected portion, the sample was kept on ice. A pathologist separated a sample of 3–5 g from a macroscopically normal area; the sample was frozen in liquid N2 within 20 min after removal and then stored at −70°C until the P450 content was determined.
Analysis of Hepatic Microsomal P450 Content and Activity.
The microsomal fraction was separated as described by van der Hoevenet al. (1974). The total P450 content was determined according to the method of Omura and Sato (1964). The CYP3A4 content was determined by Western blot analysis, using 50 pmol of total P450. We used 1, 2, and 5 pmol of purified human CYP3A as a standard to calculate the percentage of CYP3A4 in the total P450 content. Over the tested range (0.2–10 pmol), we observed a linearly increasing signal in Western blots. A human CYP3A4-selective antibody was purchased from Gentest Corp. (catalog no. 242458; Woburn, MA). It detects both CYP3A4 and CYP3A5, yielding two distinct bands in Western blots. The antibody inhibited the α-hydroxylation of midazolam by 70% at a concentration of 10 mg/nmol P450 (using the liver sample from patient 7).
The metabolism of midazolam in vitro was assessed in 0.5 ml of 0.1 M phosphate buffer, pH: 7.4, containing 50 pmol of P450 from the hepatic microsomal fraction, midazolam (2.5, 5, 10, 25, 50, 100, and 150 μM), and a NADPH-generating system. After 20 min the reaction was stopped by addition of 1 ml of methanol, and α-hydroxymidazolam concentrations were determined in the organic phase by HPLC, as previously described (Wandel et al., 1993). The peak area was linearly related to the concentration within the tested range of 25–500 ng/ml for midazolam and α-hydroxymidazolam. The interassay variability was 5.4% for midazolam and 7.5% for α-hydroxymidazolam. The Michaelis-Menten kinetics, expressed in terms of apparentVmax, apparentKM , and intrinsic clearance (apparentVmax/apparentKM ) for the α-hydroxylation of midazolam, were calculated using Eadie-Hofstee plots. Over the midazolam concentration used, we did not observe the phenomenon of substrate inhibition.
Because the dependence of midazolam’s α-hydroxylation on CYP3A content may be modified by the presence of an alternate metabolic pathway involving 4-hydroxylation and the suggested substrate activation (Ghosal et al., 1996), functional CYP3A activity was also assessed by determining the Michaelis-Menten kinetics of the oxidation of denitronifedipine with the same procedure as outlined for the α-hydroxylation of midazolam, using nifedipine concentrations of 10, 25, 50, 100, and 150 μM. For this oxidation, the considerations described for midazolam have not been proven. The denitronifedipine metabolite was determined by HPLC/UV analysis (Böcker and Guengerich, 1986; Guengerich and Böcker, 1988).
Additionally, the activity of another P450 enzyme, CYP1A2, was characterized, to demonstrate that small amounts of CYP3A in tissue samples were not due to nonspecific damage from processing. The metabolite formation rate for methoxyresorufin demethylation (up to 5 min after addition of the substrate) was measured in a total volume of 2000 μl containing the NADPH-generating system, 500 pmol of P450, and 2 μM methoxyresorufin (saturating concentration). The metabolite was determined by spectrofluorometric measurement (Burke et al., 1994; Krainev et al., 1992). The microsomal protein content was determined by the method of Lowry et al. (1951), and the specific total P450 content in microsomes was calculated as nanomoles per milligram of microsomal protein.
Analysis of Midazolam in Plasma with GC/MS.
After addition of 20 ng of diazepam (as the internal standard) to 0.5 ml of plasma, the pH was adjusted to 11 with 0.5 ml of 150 mM NaHCO3. Diethyl ether (4 ml) was added, the sample was shaken for 20 min at room temperature and centrifuged, and the organic phase was separated for evaporation under nitrogen at 60°C in a water bath. The residue was dissolved in 40 μl ofN,O-bis(trimethylsilyl)trifluoroacetamide and incubated at 60°C for 1 hr before GC/MS analysis. The peak area increased linearly over the tested concentration range of 2.5–50 ng/ml. The interassay variability was 9.7% at a midazolam concentration of 15 ng/ml.
Analysis of Pharmacokinetic Data.
The pharmacokinetics of midazolam in vivo were described by the AUC values for the observed time period (AUC0–5hr) and to infinity (AUCinf) and the clearance (dose/AUCinf)/kg body weight, which were calculated using the MKModel program with the plasma concentration-time plot fitted to a linear two-compartment model.Cmax was obtained from the plasma concentration-time plot.
Statistical Analysis.
For testing relationships among parameters, the linear regression analysis was used; for comparisons between groups, we applied the Student t test, with p < 0.05 being the minimum level of significance accepted. The data are shown as mean ± SD.
Results
Patients.
Sixteen Caucasian patients were enrolled in this study. The demographic data, the indications for partial liver resection, and the liver function test results are listed in table1. For all patients, the parameters for blood coagulation were within the normal range. For patient 13, we could not calculate the in vivo kinetics due to technical problems. For patient 15, the in vitro metabolism of midazolam and denitronifedipine could not be assessed because the liver tissue sample was inadequate. For two patients (patients 3 and 14), the immunoblots demonstrated the presence of CYP3A5 and CYP3A4. Statistical analysis was therefore performed with and without these two patients.
Demographic data for the patients, indications for liver resection, and liver function test results
In Vitro Data.
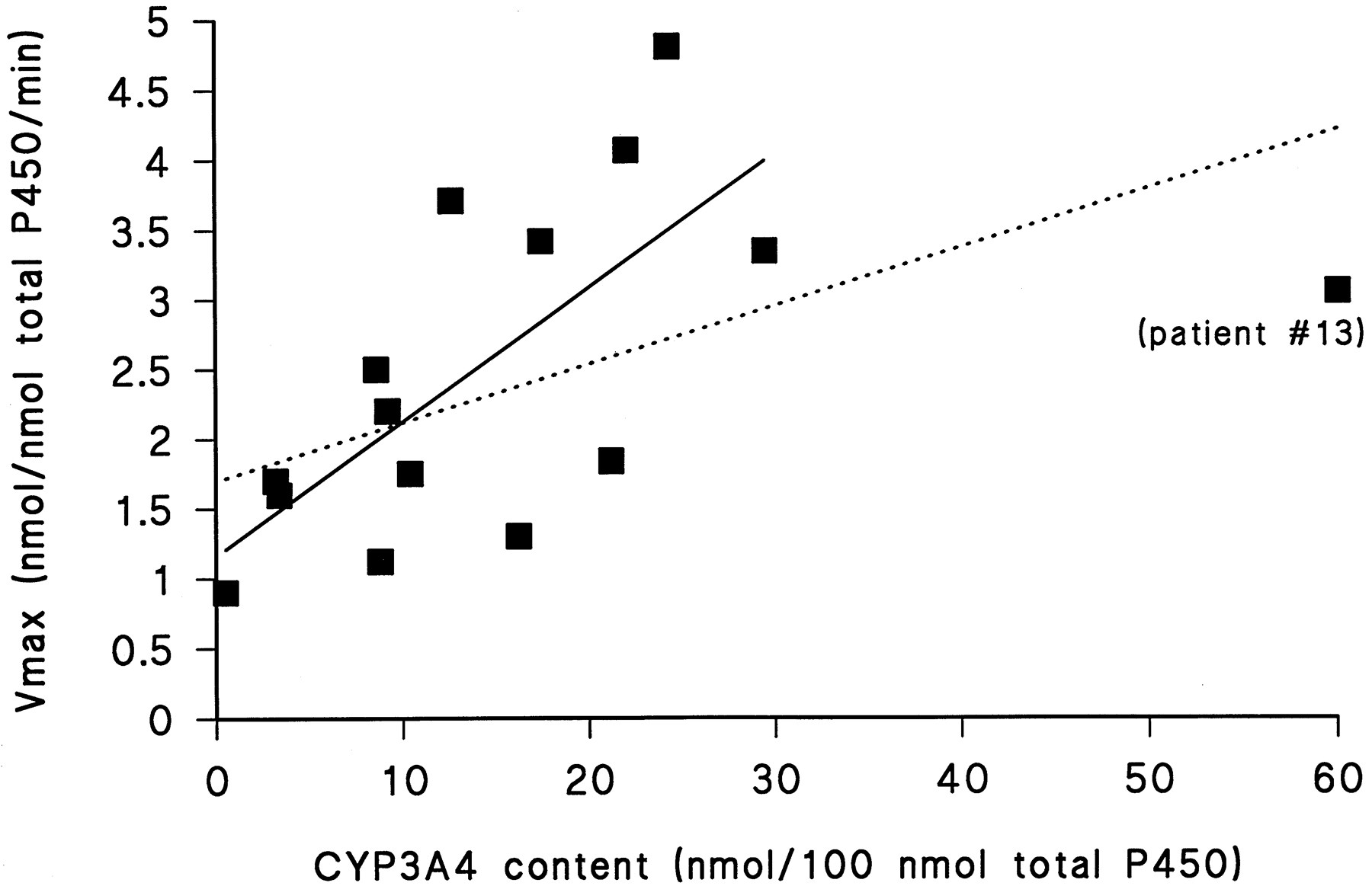
Table 2 shows the following in vitro data: the specific microsomal content of total P450, the CYP3A4 content per total P450 content, the Michaelis-Menten data for the α-hydroxylation of midazolam (including the goodness of fit) and for the oxidation of denitronifedipine (for which the goodness of fit ranged from r2 = 0.85 tor2 = 0.92), and the metabolite formation rate for the demethylation of methoxyresorufin. The average content of CYP3A4 in microsomes was 15.7 ± 14.2 nmol/100 nmol total microsomal P450. There was, as expected, a significant correlation between CYP3A4 content and the Vmax for the α-hydroxylation of midazolam (r2 = 0.31,p < 0.05), which improved tor2 = 0.52 (p < 0.01) (fig. 1) after exclusion of patient 13. It is reasonable to examine the data with and without patient 13 because the CYP3A4 content for this patient was >3 SD higher than the group mean. The in vitro/in vivo comparison is not affected by patient 13 because, as mentioned above, the in vivo disposition of po administered midazolam could not be determined for this patient due to technical problems. When the patients expressing hepatic microsomal CYP3A5, which shows a higher rate of midazolam α-hydroxylation than does CYP3A4 (Wandel et al., 1993; Gorski et al., 1994), were included, we obtained a similar correlation coefficient ofr2 = 0.5 (p < 0.01).
Summary of the in vitro parameters describing P450 content and activities

The apparent Vmaxfor the α-hydroxylation of midazolam correlated significantly with the CYP3A4 total content (r2 = 0.31,p < 0.05) (dotted line).
Without patient 13, whose CYP3A4 content was >3 SD higher than the group mean (see text), the correlation between the apparentVmax for the α-hydroxylation of midazolam and the microsomal CYP3A4 content improved tor2 = 0.52 (p < 0.01) (solid line); with inclusion of the patients expressing CYP3A5 in the liver, the correlation improved tor2 = 0.5 (p < 0.01; not shown).
The apparent Vmax for the α-hydroxylation of midazolam correlated with the rate of formation of α-hydroxymidazolam at the saturating substrate concentration of 100 μM midazolam, with r2 = 0.85 (p < 0.001). The apparentKM and intrinsic clearance for the α-hydroxylation of midazolam did not correlate with the immunologically determined CYP3A content.
The apparent Vmax for denitronifedipine oxidation also correlated with the CYP3A4 content (r2 = 0.41, p < 0.025) and, as for the α-hydroxylation of midazolam, the apparentKM and intrinsic clearance did not correlate with the microsomal CYP3A4 content. TheVmax values for the two oxidations correlated with each other, with r2 = 0.4 (p < 0.025). The even higher correlation coefficient for the comparison of the CYP3A4 content with the apparentVmax for the α-hydroxylation of midazolam, compared with that for the comparison with the apparentVmax for the oxidation of denitronifedipine, indicates that 4-hydroxylation and substrate activation did not obscure the relationship between CYP3A4 content and apparent Vmax for the α-hydroxylation of midazolam in our samples.
In Vivo Data.
Table 3 gives AUC0–5hr, AUCinf, clearance/kg, and Cmax values for midazolam. AUC0–5hr andCmax varied about 3–4-fold and AUCinf and clearance/kg about 7–8-fold.
Pharmacokinetic data for midazolam after po administration of 7.5 mg to 15 patients scheduled for partial liver resection
Relationships between In Vitro Parameters and In Vivo Data.
Clearance/kg was the only in vivo parameter that correlated with an in vitro parameter. There was a significant correlation between clearance/kg and Vmaxfor the α-hydroxylation of midazolam (r2= 0.45, p < 0.01) (fig.2). This correlation remained significant when the analysis was restricted to the patients who expressed CYP3A4 but not CYP3A5 in their liver microsomes (r2 = 0.45, p < 0.01). The correlation between clearance/kg and microsomal CYP3A4 content was slightly significant (r2 = 0.29,p < 0.05) (fig. 3) and was not significant after exclusion of both patients expressing CYP3A5 in the liver (p = 0.06).

The clearance/kg for midazolam administered po correlated with the apparent Vmax for the α-hydroxylation of midazolam (r2 = 0.45,p < 0.01).
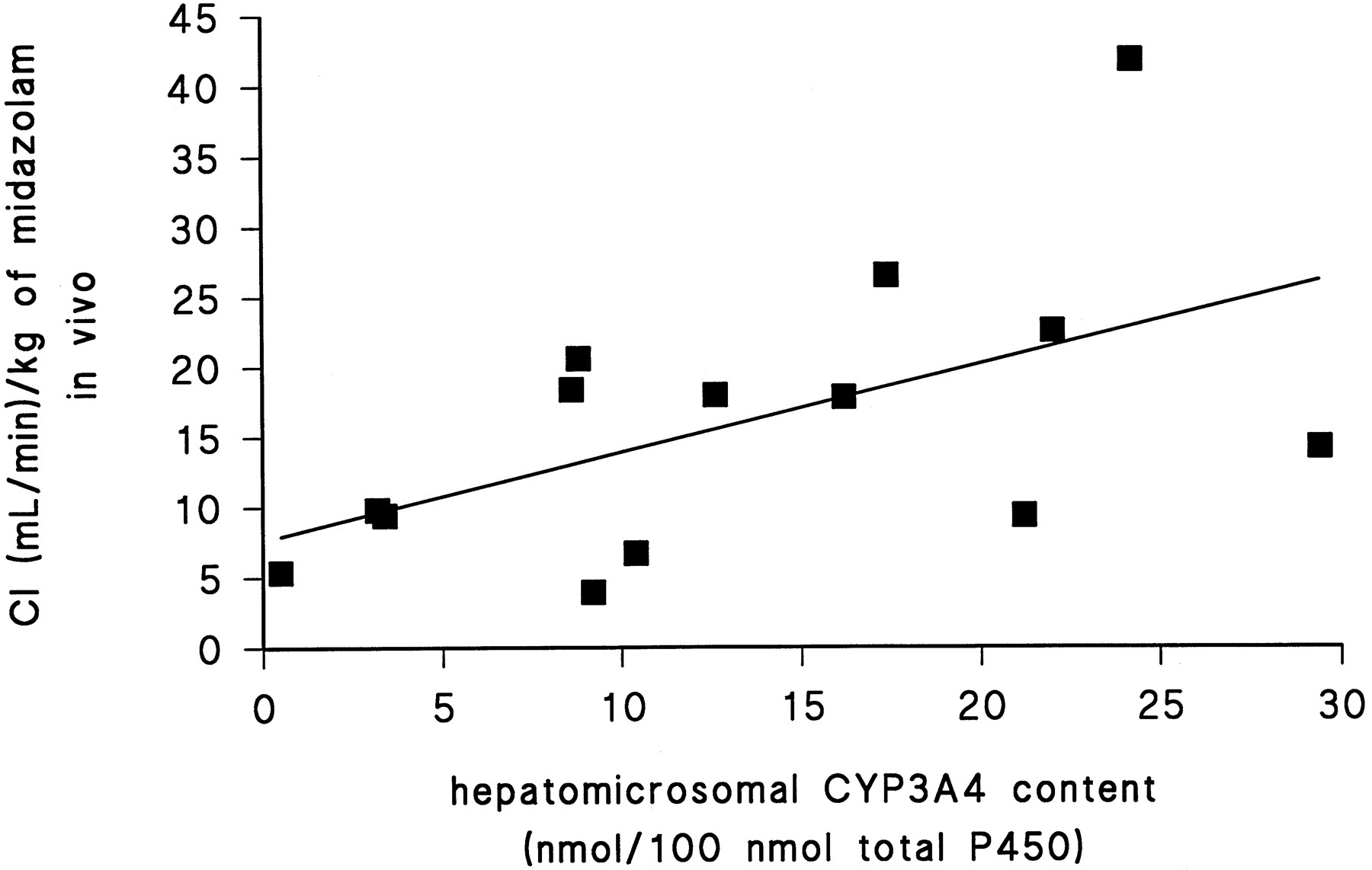
The correlation between the clearance/kg for midazolam administered po and the hepatic microsomal CYP3A4 content was significant (r2 = 0.29,p < 0.05).
The microsomal intrinsic clearance did not correlate with any in vivo pharmacokinetic parameter for midazolam. The same was true when the predicted hepatic organ clearance was compared with the disposition of midazolam in vivo. The predicted hepatic organ clearance of midazolam by α-hydroxylation was calculated as outlined by Thummel et al. (1994).
Discussion
This study found that patients with a low rate of formation of α-hydroxymidazolam in their liver microsomes are more likely to exhibit a low clearance/kg for midazolam after its po administration than patients with high activity for the α-hydroxylation of midazolam. This finding is clinically relevant because the pharmacodynamic effects of midazolam are related to its plasma levels (Koopmans et al., 1988; Persson et al., 1988). However, the correlation coefficient for the correlation between hepatic microsomal midazolam metabolism and the in vivodisposition of midazolam administered po (r2 = 0.5) was lower than that for the correlation between microsomal CYP3A content and clearance/kg observed after iv administration of midazolam (r2 = 0.86, p < 0.01) (Thummel et al., 1994).
Two aspects may contribute to the weaker relationship between thein vivo and in vitro disposition of midazolam after po administration, compared with iv administration. Firstly, midazolam is exposed to intestinal CYP3A to a much greater extent after po administration than after iv administration, and the intestinal extraction ratio for midazolam administered po has been calculated to be 0.43 (Thummel et al., 1996; Paine et al., 1996). Because intestinal and hepatic CYP3A activities do not seem to be co-regulated (Thummel et al., 1996), the relationship between the pharmacokinetics of po administered midazolam and hepatic CYP3A activity may be attenuated due to the increased influence of intestinal CYP3A activity. Secondly, the in vivo/in vitro comparison after iv administration of midazolam was performed with patients undergoing liver transplantation, who were being treated with drugs that may have influenced both the in vivo and in vitro kinetics of midazolam. The inclusion of such patients who are receiving medications, including CYP3A inducers (glucocorticoids) (Watkins et al., 1985; Schuetzet al., 1984) or CYP3A substrates (cyclosporine) (Combalbertet al., 1988; Kronbach et al., 1988), may make the in vivo/in vitro relationship more apparent.
The hepatic organ clearance was found to predict the total clearance of midazolam after iv administration, with hepatic α-hydroxylation of midazolam accounting for 70% of the total midazolam clearance (Thummelet al., 1994). When we applied the same formula for calculation of the hepatic organ clearance of midazolam to our data, the calculated hepatic organ clearance accounted for only 10% of the total in vivo clearance and did not correlate with anyin vivo pharmacokinetic parameter for midazolam. These findings probably reflect the profound effect of intestinal extraction on po administered midazolam. In this context, it should be noted that hepatic organ clearance in each study was calculated without exact measurement of hepatic blood flow and liver weight, which were estimated according to body weight.
The correlation between in vitro and in vivomeasures of the disposition of drugs may be affected by the process of obtaining microsomes for in vitro studies. Potential considerations include not only variability in liver weight but also variability in the recovery of P450 in the microsomes from the isolation procedure. These factors may have contributed to the relatively weak in vitro/in vivo relationship observed in this study, despite the fact that the range of specific microsomal P450 contents is narrow and comparable to that reported by other research groups (Sadeque et al., 1997). It should be noted that the significant correlation between clearance/kg and CYP3A content in microsomes after iv midazolam administration (r2 = 0.86, p < 0.01) (Thummel et al., 1994) was seen even without normalization of the microsomal CYP3A content to liver weight, emphasizing the close relationship between hepatic microsomal CYP3A content and clearance/kg when the contribution of intestinal CYP3A activity to the disposition of midazolam in vivo is eliminated (Paine et al., 1996). Additionally, to confirm that the variability in hepatic microsomal CYP3A activity and content was not due to any nonspecific damage to the liver samples from processing, we determined the activity of another P450 enzyme, CYP1A2. Although such a procedure does not definitely exclude the possibility of destruction of P450 activity during processing, the high CYP1A2 activity in samples with low CYP3A activity (table 2) (in the same range as for the liver samples with high CYP3A content and activity) indicates that it is unlikely that nonspecific tissue damage affected microsomal P450 activity.
When generalizing from the in vivo/in vitrorelationship found in this study, one must consider that the liver tissues were obtained from patients scheduled for partial liver resection for a number of reasons, including resection of liver metastases from colon cancer. Although liver function tests did not indicate general liver disorders and the disease process was locally limited within the liver, it is uncertain whether and how much the CYP3A activity was changed by the underlying disease. It should be noted that Guengerich and Turvy (1991) showed that the CYP3A levels in livers of patients with metastatic colon cancer did not differ significantly from those in liver tissue obtained from liver transplant donors. Anesthetic agents administered intraoperatively may also influence hepatic CYP3A activity. In vitro experiments showed inhibition of some P450 enzymes by propofol (Chen et al., 1995a; Baker et al., 1993), but this was not observed in vivo (Chen et al., 1995b). Fentanyl was shown to be metabolized by CYP3A in vitro (Feierman, 1996; Tateishi et al., 1996); however, in rats the acute administration of fentanyl did not alter CYP3A activity (Loch et al., 1995). Isoflurane is mainly metabolized by CYP2E1 (Kharasch and Thummel, 1993). Previous studies did not indicate that isoflurane influences CYP3A activity (Baker et al., 1995; Baker and Ronnenberg, 1992). Thus, it seems unlikely that the anesthetic agents used altered CYP3A activity significantly in our patients.
We conclude that low in vitro hepatic microsomal activity for the α-hydroxylation of midazolam predicts a lower clearance/kg value in vivo after po midazolam administration. However, after po administration both the intestinal and hepatic CYP3A activities are involved in the in vivo pharmacokinetics for midazolam, and this may explain the weaker relationship betweenin vitro kinetics and in vivo disposition of midazolam after po, compared with iv, administration.
Acknowledgments
We are grateful to K. Schmidt, Department of Pharmacology and Toxicology, Friedrich-Alexander-University (Erlangen, Germany), and W. Forster, Division of Clinical Pharmacology, Ruprecht-Karls-University (Heidelberg, Germany), for excellent technical assistance in the preparation of microsomes and the measurement of midazolam in plasma.
Footnotes
-
Send reprint requests to: Christoph Wandel, M.D., Division of Clinical Pharmacology, Vanderbilt University, Nashville, TN 37232.
-
This work was supported in part by Hoffmann-LaRoche AG (Grenzach-Wyhlen, Germany).
- Abbreviation used is::
- P450 or CYP
- cytochrome P450
- Received January 31, 1997.
- Accepted October 6, 1997.
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}
{kind=link}
{kind=link}