


1996 ASPET N-Glucuronidation of Xenobiotics Symposium
Abstract
Conjugation of many primary, secondary, and tertiary amine–containing xenobiotics with glucuronic acid can result in the formation of N-glucuronide metabolites. For carcinogenic arylamines and their N-hydroxylated metabolites,N-glucuronidation can result in the formation of either inactive metabolites or labile conjugates, which can be transported to their target tissue (urinary bladder) where they may be converted to reactive metabolites. Drugs with primary amine (e.g.dapsone) or secondary amine moieties (e.g. sulfadimethoxine and clozapine) can also be metabolized to N-glucuronides. The metabolism of a number of tertiary amine–containing pharmacological agents to quaternary ammonium–linked glucuronides represents a unique and important metabolic pathway for these compounds that is highly species-dependent. This review summarizes our present knowledge of the uridine diphosphate (UDP)-glucuronosyltransferase enzymes involved in catalyzing N-glucuronide formation. Of the more than 30 UDP-glucuronosyltransferases that have been purified or cloned and expressed, many catalyze N-glucuronide formation for primary and secondary amine substrates. In contrast, only human UDP-glucuronosyltransferases 1A3 and 1A4 have been shown to catalyze quaternary ammonium–linked glucuronide formation for aliphatic tertiary amines. The structure of the UGT1 gene complex is highly conserved across species, and it appears that a mutation in the first exon encoding UDP-glucuronosyltransferase 1A4, resulting in a pseudo-gene, may explain the inability of some species to form quaternary ammonium–linked glucuronides.
In 1949, Smith and Williams (1949) showed that the main urinary metabolite (more than 50% of the dose) of aniline in the rabbit was an acid-labile glucuronide and suggested that this metabolite was anilineN-glucuronide. Subsequently, many primary, secondary, and tertiary amines have been shown to be excreted asN-glucuronide metabolites. In general, glucuronidation is considered to be a significant means of detoxication and elimination from the body because of the conversion of hydrophobic xeno- and endobiotics to hydrophilic metabolites. UnderstandingN-glucuronidation and identifying the enzymes involved in this process and their regulation is especially important in humans for two reasons. First, N-glucuronidation of many primary and secondary aromatic amines has been suggested to have implications in the potential carcinogenicity of these compounds. In addition, metabolic elimination of many tertiary amine therapeutic agents used in humans (antipsychotic agents, antihistamines, and tricyclic antidepressants) is dependent on N-glucuronidation of the parent drugs to form quaternary ammonium–linked glucuronide metabolites.
Excretion of N-glucuronides of aromatic amines (e.g. α- and β-NA1 4-ABP, or benzidine) in urine accounts for only a low percentage of the administered dose of the compound. This is due to the competing metabolic pathways that exist for primary amines and to the lability of the aromatic amineN-glucuronides (Boyland and Manson, 1966; Kadlubar et al., 1977; Kadlubar et al., 1981; Hammons et al., 1985; Gorrod and Manson, 1986; Babu et al., 1992;Kadlubar et al., 1992). In general, primary aromatic amines are eliminated by three competing metabolic processes:N-acetylation, oxidation to form N- or ring-hydroxylated metabolites, and conjugation of the parent amine or of the N-hydroxylated metabolite. N-acetylation is thought to lead to detoxication of aromatic amines because it decreases the amount of amine that can be oxidized toN-hydroxylated compounds (Hein, 1988). Indeed, it has been shown that individuals expressing the slow acetylator phenotype had higher amounts of 4-ABP adduct formation, compared with high acetylators (Vineis et al., 1990; Vineis et al., 1994). In contrast to the inactivation of aromatic amines byN-acetylation, N-hydroxylation of aromatic amines is correlated with adduct formation from these compounds (Kadlubaret al., 1977). The presumptive reactive intermediates involved in this covalent binding are nitrenium ions formed from theN-hydroxylated aromatic amines (Kadlubar et al., 1977). Hydroxylation of the aromatic ring, on the other hand, results in the formation of metabolites that are substrates for sulfation andO-glucuronidation (Boyland and Manson, 1966; Gorrod and Manson, 1986). Sulfation or glucuronidation of the primary aromatic amine or its N-hydroxylated secondary amine metabolite are the two main conjugation reactions involved in the elimination of these compounds.
As shown by Smith and Williams (1949) and others (Babu et al., 1992; Babu et al., 1995),N-glucuronides of all primary amines and many secondary amines are very labile and are susceptible to cleavage to the parent amine and glucuronic acid under very mildly acidic conditions. For example, the half-time for stability of benzidineN-glucuronide has been shown to be approximately 3 min at pH 5.5 (Babu et al., 1992). At low pH,N-glucuronides of N-hydroxylated metabolites of α-NA, β-NA, and 4-ABP (secondary amines) have also been shown be converted to their corresponding N-hydroxylated derivatives, which bind DNA (Irving, 1977; Kadlubar et al., 1977; Gorrod and Manson, 1986). It has been proposed that N-glucuronides of N-hydroxy aromatic amines may act as the transport mechanism for delivering carcinogenic amines to the urinary bladder (Kadlubar et al., 1977; Poupko et al., 1979). Benzidine (4-4′-diaminobiphenyl) has two amine groups that can undergoN-acetylation, N-hydroxylation, andN-glucuronidation. Babu et al. (1995) suggested that hepatic N′-glucuronidation ofN-acetylbenzidine can result in the delivery of the parentN-acetylated N′-amine to the bladder epithelium, whereas hepatic N′-glucuronidation of theN′-hydroxylated metabolites of N-acetylbenzidine leads to a product that has a higher acid stability. Thus the latter reaction might be considered a detoxification process.
Many chemicals that contain aliphatic tertiary amine moieties are clinically useful drugs, such as antihistamines, tricyclic antidepressants, and antipsychotic agents. The metabolic processes involved in the overall elimination of aliphatic tertiary amines include N-demethylation, hydroxylation reactions, andN-glucuronidation. Demethylation of tertiary amines generally results in the formation of compounds with reduced clinical efficacy. Oxidation of compounds with tertiary amine moieties also generally results in inactivation of the drugs and aids in their elimination by the formation of hydroxyl groups that can be conjugated to form O-glucuronides or sulfates.N-Glucuronidation of the tertiary amine group results in the formation of quaternary ammonium–linked glucuronides. Quaternary ammonium–linked glucuronide formation is highly species-dependent (Fischer et al., 1980; Dulik and Fenselau, 1987; Coughtrie and Sharp, 1991; Remmel and Sinz, 1991) and is an important elimination pathway for many tertiary amines in humans. For example, the quaternary ammonium–linked glucuronide of tripelennamine represents the major metabolic product of this compound found in the urine (Chaudhuriet al., 1976). In contrast to N-glucuronides of primary and secondary amines, which tend to be acid-labile, quaternary ammonium glucuronides tend to be susceptible to hydrolysis under mildly basic conditions. Dulik and Fenselau (1987) reported that at pH 8.5, about 40% of cyproheptadine quaternary-ammonium glucuronide is hydrolyzed over a 24-hr period and suggested that this is an explanation for the inability of investigators to detect the presence of cyproheptadine quaternary ammonium glucuronide in rabbit urine.
UDP-Glucuronosyltransferases and N-Glucuronidation
Glucuronidation of endo- and xenobiotics is catalyzed by the UGT superfamily of enzymes. UGTs catalyze the transfer of glucuronic acid from UDP-glucuronic acid to compounds with amine, hydroxyl, and carboxylic acid moieties (Clarke and Burchell, 1994; Burchell et al., 1995). Individual UGT isoforms do not catalyze the conjugation of only a single type of chemical moiety; in other words, no single UGT catalyzes conjugation of only amine substrates—UGTs generally demonstrate conjugation reactivity toward diverse chemical moieties. Despite this diversity of reactivity, each UGT isoform can be distinguished functionally from another based on its reactivity and/or nonreactivity with different classes of prototypical substrates. For example, expressed rat UGT2B1 protein catalyzes the glucuronidation of many simple phenolic compounds and coumarins like many other UGTs. However, it also catalyzes the glucuronidation of the 17-hydroxyl position of testosterone and related steroids, the hydroxyl group of chloramphenicol, the carboxylic acid moiety of profen nonsteroidal anti-inflammatory drugs and the phenolic 3-hydroxy group of morphine (Mackenzie, 1986; Mackenzie, 1987; Pritchard et al., 1994). The major pharmacologic importance of rat UGT2B1 is its ability to catalyze conjugation of morphine and other opioids. Recent studies in our laboratory suggest that rat UGT2B1 is the major enzyme in untreated rat liver that catalyzes the glucuronidation of morphine in this species (King et al., 1997). Another UGT isoform, rat UGT1A1, also catalyzes morphine glucuronidation, but at much lower rates compared to the UGT2B1 isoform (King et al., 1997). A discussion of the complex substrate specificities of UGTs is outside the scope of this review; however, recent reviews address this subject (Clarke and Burchell, 1994; Burchell et al., 1995).
Currently, UGTs are named in accordance with a nomenclature system based on evolutionary divergence of the UGT genes (Mackenzie et al., 1997), and this nomenclature will be used in this review. To date, three UGT families have been identified in humans:UGT1, UGT2, and UGT8. Of these three families, UGT1 and UGT2 proteins have been shown to catalyze glucuronidation of xenobiotics. The UGT1 and UGT2genes appear to be structurally different in that UGT1 proteins result from alternate slicing of different first exons with five shared exons encoded by the UGT1 gene complex, while UGT2 proteins appear to be encoded by unique genes. In the human genome, at least 12 different first exons have been identified for the UGT1 gene (Ritter et al., 1992; Cho et al., 1995). A further consideration of the UGT1 gene complex and the UGT proteins encoded by it will be discussed later in this review. TheUGT2 gene family is currently divided into three subfamilies: UGT2A, UGT2B, and UGT2C. Only one member of the UGT2A subfamily has been described, namely, UGT2A1. UGT2A1 was isolated from rat and bovine olfactory epithelium and is preferentially expressed in this tissue (Lazardet al., 1991). In contrast, over 18 members of theUGT2B subfamily have now been isolated (Mackenzie et al., 1997). In general, proteins encoded by the UGT2Bsubfamily have been characterized primarily for their ability to catalyze the glucuronidation of steroids. However, as noted above for rat UGT2B1, the pharmacologic and physiologic importance of the enzymes of the UGT2B gene subfamily may not relate entirely to their ability to catalyze steroid glucuronidation. Little is known about the substrate specificity of the UGT2C1 protein.
Over the last several years, many UGTs, from a number of different species, have been shown to catalyze the glucuronidation of amines. The purpose of this review is to present current information about the enzymes that catalyze N-glucuronidation.N-Glucuronidation of primary amines has been demonstrated for a number of purified and cloned and expressed UGT proteins. More recently, human and rabbit UGT cDNAs have been shown to encode for proteins that catalyze the glucuronidation of primary, secondary, and tertiary amines. In addition, evidence will be presented which suggests that there is a correlation between species that are able to conjugate sapogenins with glucuronic acid and those that can form quaternary ammonium–linked glucuronides. Finally, a genetic basis for the ability, or inability, of a species to glucuronidate some classes of tertiary amines will be postulated.
N-Glucuronidation of Primary and Secondary Amines Catalyzed by Purified and Cloned and Expressed Rat UGTs.N-Glucuronidation of primary amines occurs in many species, but this process has been studied most extensively in the rat. The most commonly investigated substrates have been α- and β-NA, and 4-ABP, α- and β-NA are planar amines, whereas 4-ABP is considered to be more “bulky.”
Three purified rat liver UGTs have been shown to catalyzeN-glucuronidation of primary amines (table1). Purified rat liver 3α-hydroxysteroid UGT (UGT2B2), 17β-hydroxysteroid UGT (UGT2B3), and a 3-methylcholanthrene-inducible p-nitrophenol UGT (UGT1A6) catalyze the glucuronidation of α- and β-NA, whereas glucuronidation of 4-ABP is only catalyzed by UGT2B2 (Green and Tephly, 1987). In addition, purified rat UGT2B2 has been shown to catalyze the glucuronidation of aniline (Roy Chowdhury et al., 1986). For each purified enzyme, α-NA was the best substrate, in that its catalytic efficiency (Vmax/Km ) was at least tenfold higher than either β-NA or 4-ABP (Green and Tephly, 1987). Primary amines are not substrates for purified rat liver digitoxigenin mono-digitoxoside UGT, morphine UGT, and 4-hydroxybiphenyl UGT (von Meyerinck et al., 1985; Puig and Tephly, 1986; Styczynskiet al., 1991). Glucuronidation of secondary amines by purified rat liver UGTs has received less attention. Only purified rat liver UGT1A6 has been tested for reactivity toward a secondary amine substrate (N-OH β-NA) (Bock et al., 1979).
Reactivity of rat UGTs with primary and secondary amines
In an attempt to determine the potential role of UGT2B2 in the glucuronidation of aromatic amines in vivo, we determined UGT activity for α- and β-NA and 4-ABP in microsomal preparations from HA and LA Wistar rats (Green and Tephly, 1987). LA Wistar rats are deficient in their ability to glucuronidate androsterone and other substrates for UGT2B2, compared with normal glucuronidation rates for these substrates in HA Wistar rats (Matsui and Nagai, 1986). The inability of LA Wistar rats to glucuronidate UGT2B2 substrates is due to a deletion in the UGT2B2 gene in these animals (Corseret al., 1987). Our results showed that hepatic LA Wistar rat microsomal UGT activity for α-NA, β-NA and 4-ABP were 60%, 50%, and 20%, respectively, of the rates obtained in microsomes from HA Wistar rats. These data suggest that a significant amount of primary aromatic amine glucuronidation is catalyzed by UGT2B2 in untreated rat liver. However, other UGTs also catalyze the glucuronidation of primary amines in rat liver.
Rat UGT2B2 and 2B3 appear to be constitutively expressed proteins in rat liver and do not appear to be significantly inducible by xenobiotic treatment (Mackenzie, 1987; Haque et al., 1991). However, expression of rat hepatic UGT1A6 is induced by polycyclic aromatic hydrocarbons. Orzechowski et al. (1994) have shown that hepatic microsomal N-glucuronidation of α- and β-NA and N-OH β-NA is induced by treatment of rats with 3-methylcholanthrene, presumably because of induction of UGT 1A6, whereas N-glucuronidation of 4-ABP and N-OH 4-ABP is not enhanced by 3-methylcholanthrene treatment. These data confirm the inability of polycyclic aromatic hydrocarbons to induce rat UGT2B2 and suggest that the UGT2B2 enzyme is important for glucuronidation of both 4-ABP and N-OH 4-ABP. Also, because 3-methylcholanthrene induced rat UGT1A6 glucuronidation activity toward the naphthylamines over twofold, this enzyme is probably the predominant protein involved in their conjugation in 3-methylcholanthrene–treated animals.
Similar to the results obtained from preparations of purified rat UGT1A6, the expressed enzyme also was shown to catalyze the glucuronidation of primary and secondary amines (table 1). The substrate specificity and reactivity of the purified enzyme and the expressed protein are similar. Stably expressed rat UGT1A6 glucuronidates planar aromatic amines and N-hydroxylated naphthylamines at higher rates, compared with the bulkier 4-ABP and N-OH 4-ABP (Orzechowski et al., 1994). In contrast to the results of studies using purified rat UGT1A6 (Green and Tephly, 1987),Orzechowski et al. (1994) showed that stably expressed rat UGT1A6 protein catalyzed the glucuronidation of 4-ABP, though at very low rates. The inability to detect 4-ABP glucuronidation activity in purified enzyme preparations is probably due to the obligatory presence of detergent in these preparations, which may inhibitN-glucuronidation (Green and Tephly, 1996). In addition, the low levels of 4-ABP glucuronidation activity in rat liver that is catalyzed by enzymes other than rat UGT2B2 probably explain the very low levels of 4-ABP glucuronidation activity in LA Wistar rats. Expressed rat UGT2B1 protein displays very low catalytic activity for the glucuronidation of β-NA (Pritchard et al., 1994). Glucuronidation of primary amines has not been detected when cloned and expressed rat UGT2B6, 2B12, and 1A1 were examined (Mackenzie, 1990;Green et al., 1995; King et al., 1996).
N-Glucuronidation of Primary and Secondary Amines Catalyzed by Human and Rabbit UGTs.
Rabbits have been a useful species for the characterization of oxidative drug-metabolizing enzymes’ genes and have proven useful as an animal model to investigate the molecular events associated with the expression of some of these genes. In addition, based upon the speciation period of rodents, legamorphs, and humans, legamorphs appear to be more closely related in evolution to humans than are rodents. For example, the predicted primary amino acid sequence of rabbit UGT1A6 is most related to the human (81%) UGT1A6, compared with the rat (78%) and mouse (78%) enzymes (Lamb et al., 1994). Therefore, the identification and characterization of UGT genes and gene products from rabbits could serve to broaden our understanding of how these genes have evolved, while providing an additional mechanism to study how the UGT genes may be regulated in humans.
Purified rabbit and human UGTs that catalyze the glucuronidation of primary and secondary amines are listed in table2. A purified rabbit hepatic UGT that catalyzes the glucuronidation of estrone with high efficiency also catalyzes the N-glucuronidation of primary amines (Tephlyet al., 1988). Similar to the results obtained for purified rat UGT2B2, purified rabbit estrone UGT catalyzes the glucuronidation of both planar and bulky primary amines. Interestingly, the glucuronidation rates for α- and β-NA and 4-ABP using purified rabbit estrone UGT are comparable to each other. In contrast, a purified rabbit liver UGT that catalyzes the glucuronidation of phenolic compounds (rabbit p-nitrophenol UGT) does not catalyze the glucuronidation of primary amines.
Reactivity of rabbit and human UGTs with primary and secondary amines
Two purified human UGTs have been shown to catalyze the glucuronidation of primary amines (Irshaid and Tephly, 1987). The reactivity of purified human pI7.4 UGT for primary amines is similar to that observed for purified rat UGT2B3 and 1A6 in that only planar amines are substrates and that α-NA is a better substrate than β-NA. Irshaid and Tephly (1987) showed that the bulky amine, 4-ABP, was not a substrate for this purified enzyme. Purified human pI6.2 UGT showed reactivity toward amine substrates similar to that observed for purified rat UGT2B2 in that the glucuronidation rate for 4-ABP is intermediate to those of α- and β-NA.
Of the five members of the human UGT1 gene family that have been cloned and expressed to date, four UGT isoforms have been shown to catalyze the glucuronidation of primary and secondary amines (table2). For stably expressed human UGT1A6 and UGT1A9, planar aromatic amines are better substrates than the bulky 4-ABP (Orzechowski et al., 1994). In contrast, stably expressed human UGT1A4 catalyzes the glucuronidation of planar amines at enzymatic rates that are comparable to those of the more bulky amines, such as 2- and 4-ABP (Green et al., 1995; Green and Tephly, 1996). Interestingly, the position of the amino group in 2-ABP (compared with 4-ABP) did not have any effect on the glucuronidation efficiency for these two amines for the expressed human UGT1A4 (Green and Tephly, 1996), whereas it did for expressed human UGT1A3 (Greenet al., 1998). For expressed human UGT1A4, addition of a second amine to the biphenyl moiety [e.g. benzidine (4, 4′ diaminobiphenyl)] or the addition of a sulfone moiety between the benzene rings [e.g. dapsone (4-4′-diaminobiphenyl sulfone)] greatly decreased glucuronidation rates. The decreased glucuronidation rates for these substrates appear to be due to increased Km values (Green and Tephly, 1996).
While both human UGT1A3 and UGT1A4 catalyze glucuronidation of primary amines, the glucuronidation efficiencies for these substrates for the two expressed proteins are different. In contrast to expressed human UGT1A4, which catalyzes the glucuronidation of amines with high efficiency, glucuronidation of amines catalyzed by UGT1A3 exhibits low efficiency. The low glucuronidation efficiency of UGT1A3 for primary amines is due to 100-fold–higher apparent Kmvalues for the substrates, compared with the values obtained for UGT1A4 (Green et al., 1998). Thus it is probable that UGT1A3 makes only a limited contribution to the metabolic elimination of these compounds in vivo.
A variety of secondary amines have been tested as substrates for expressed human UGTs (table 2). Similar to the results obtained for expressed rat UGT1A6, N-OH β-NA and N-OH 4-ABP are also substrates for expressed human UGT1A6 (Orzechowski et al., 1994). In addition, these compounds are also substrates for expressed human UGT1A9 (Ebner and Burchell, 1993). In all cases, the planar naphthylamines were better substrates, compared with the bulky aminobiphenyls. Other classes of amines, such as triazoles and tetrazoles, are substrates for expressed human UGT1A6 (Huskey et al., 1994). N,N-diphenyl amine and desmethyl clozapine are substrates for expressed human UGT1A4 (Green and Tephly, 1996).
Stably expressed human UGT2B15 protein has been shown not to catalyze the glucuronidation of primary amines (Green et al., 1994). The ability of other members of the human UGT2 gene family to catalyze the glucuronidation of primary and secondary amines is still largely unknown.
Glucuronidation of Tertiary Amines Catalyzed by Expressed Human and Rabbit UGT Proteins.
While expressed human UGT1A9, 1A4, 1A3, and 1A6 react with some of the same primary and secondary amine substrates, their reactivity with tertiary amine substrates varies considerably. In humans, tertiary amines are an important class of substrates because many clinically important therapeutic agents have this chemical moiety and are extensively conjugated to form quaternary ammonium–linked glucuronides. Expressed human UGT1A3 and 1A4 catalyze the glucuronidation of a wide variety of aliphatic tertiary amines (table3).
Reactivity of cloned and expressed UGTs with tertiary amines
The tertiary amines glucuronidated by UGT1A4 contain either anN,N-dimethyl moiety at the end of a side chain, an N-methylated piperazine, or an N-methylated piperidinyl moiety (fig. 1A). While the structure-activity requirements for quaternary ammonium–linked glucuronide formation have not been extensively investigated, some general observations on the reactivity of tertiary amine substrates for expressed human UGT1A4 protein can be made. Compounds that have a propyl N,N-dimethyl side chain extending from a polycyclic nucleus (e.g.amitriptyline or imipramine) are glucuronidated at higher rates, compared with compounds that have a propyl N-methyl piperazine (e.g. cyproheptadine) or piperdine side chain (e.g. clozapine). It appears that the presence of sulfur or oxygen atoms in the polycyclic nucleus leads to lower glucuronidation rates, whereas the presence of nitrogen atoms in the polycyclic nucleus does not. The presence of a polycyclic nucleus favors the reactivity of tertiary amines. Compounds that do not possess a polycyclic nucleus (e.g. diphenhydramine, cyclizine, and pheniramine analogs [fig. 1B]) are glucuronidated at low rates.N,N-Dimethyl aniline is not a substrate for expressed human UGT1A4, and UGT1A4 does not catalyze the glucuronidation of morphine and similar opioids (Green and Tephly, 1996).
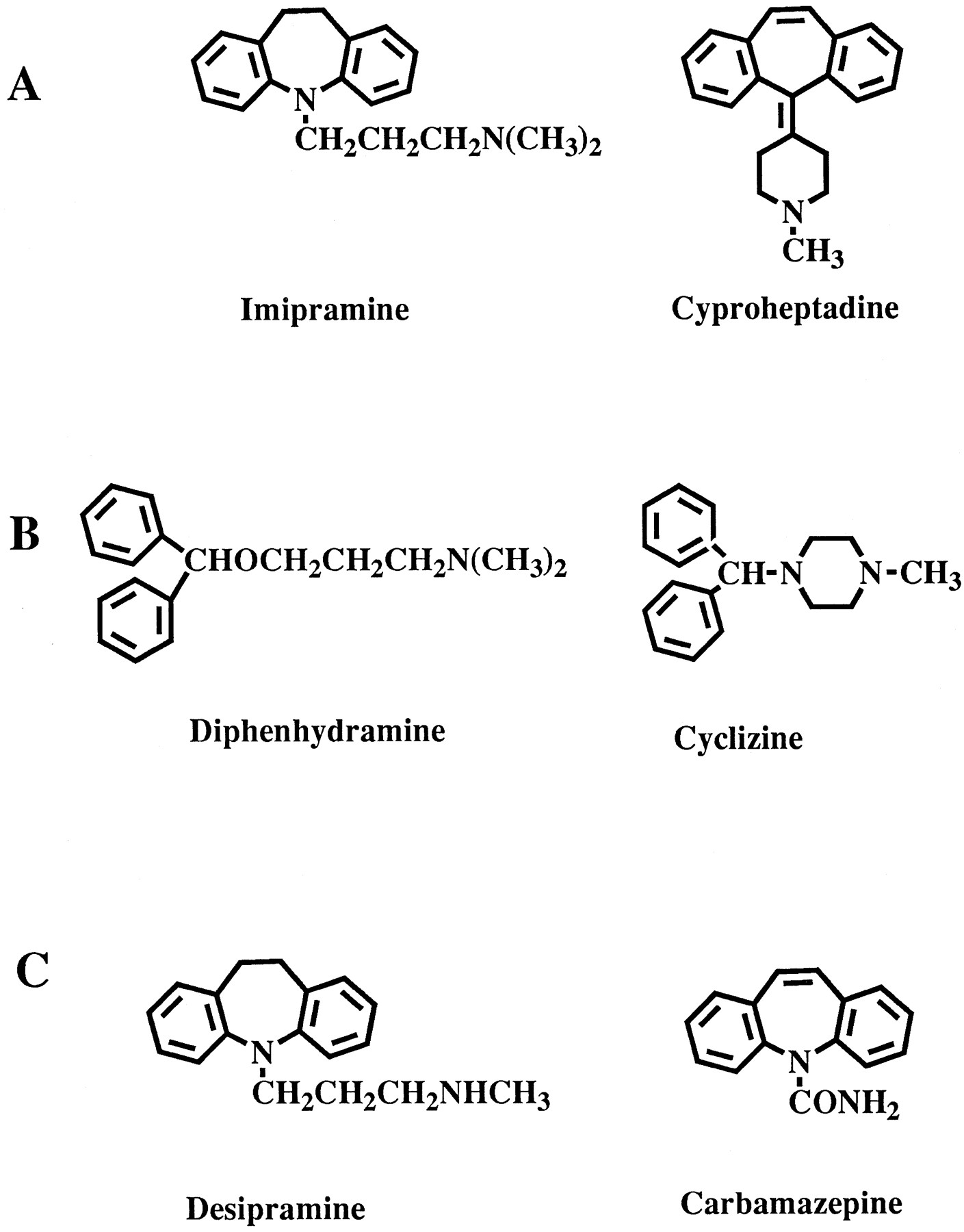
Chemical structures for various amine-containing drugs.
Prototypical polycyclic amines that are substrates for expressed human UGT1A4 are depicted in row A. Other tertiary amine substrates for expressed human UGT1A4 are shown in row B. Amines that are not substrates for the expressed enzyme are shown inrow C.
It is interesting to note that the N-dimethylated metabolites of imipramine and amitriptyline (desipramine [fig.1C] and nortriptyline, respectively) are not substrates for expressed human UGT1A4. In contrast, the demethylated metabolite of clozapine is a substrate for the enzyme. These data suggest that demethylated piperazines and piperdine moieties interact with the active site of the human UGT1A4 protein better than demethylated propylN,N-dimethyl side chains. Expressed human UGT1A4 does not catalyze the glucuronidation of carbamazepine (fig.1C) at the amide nitrogen. Sulfadimethoxine (fig.1C) and sulfathiazole, which form sulfonamideN-glucuronides (Bridges et al., 1961; Dulik and Fenselau, 1987) despite having a free amino group, are also not substrates for expressed human UGT1A4.
Besides catalyzing the glucuronidation of amines, expressed human UGT1A4 catalyzes glucuronidation at aliphatic hydroxyl groups (Green and Tephly, 1996). Stably expressed human UGT1A4 exhibits a very limited range of substrate specificity. In this respect, simple phenols, monoterpenoid alcohols, coumarins, anthraquinones, flavonoids, and estrogens are either not substrates for the enzyme or are glucuronidated at very low rates. The low reactivity of simple phenols and monoterpenoid alcohols is due to the very high apparentKm values for these compounds, which result in very low glucuronidation efficiencies (Green and Tephly, 1996). In contrast, expressed human UGT1A4 protein catalyzes the glucuronidation of sapogenins, progestins, and certain androgens are glucuronidated by expressed human UGT1A4 at quite high rates and with very high efficiencies (Green and Tephly, 1996). Of the progestins tested, pregnanediols and 16α-hydroxy pregnenolone displayed the highest rates of glucuronidation. Of the xenobiotics tested, sapogenins have been shown to be the best substrates for expressed human UGT1A4 protein. The glucuronidation efficiencies for sapogenins, pregnanediol, androstanediol, and certain amines are comparable, suggesting that these are the preferred substrates for human UGT1A4.
Sapogenins are interesting compounds in that they are naturally occurring plant-derived steroidal compounds. They are widely used in the chemical industry as synthetic precursors for the synthesis of many important animal steroids. We have determined that sapogenins are not substrates for rat UGT2B12, 2B1, and 1A1; they are also not substrates for expressed human UGT2B15 and 1A1. These data led us to try to determine whether sapogenins are specific substrates for UGT1A4. In order to test this hypothesis, we studied the ability of hepatic microsomes from different species to catalyze the glucuronidation sapogenins and aliphatic tertiary amines. Data in table4 show that, as expected, glucuronidation of sapogenins and imipramine were detected in human liver microsomes, but that sapogenin and imipramine glucuronidation was not detected in rat liver microsomes. These data suggest that sapogenins, like aliphatic tertiary amines, are specific substrates for expressed human UGT1A4.
Glucuronidation of imipramine and sapogenins by hepatic microsomes from different species
One area of controversy concerning the function of human UGT1A4 is whether this enzyme plays any role in the glucuronidation of bilirubin.Ritter et al. (1991) originally showed that the expressed enzyme catalyzed the glucuronidation of bilirubin. Subsequently, Bosmaet al. (1994) showed that UGT1A4 does not catalyze bilirubin glucuronidation and concluded that human UGT1A1 is the only physiologically relevant enzyme involved in bilirubin glucuronidation in human liver. In support of their hypothesis, Crigler-Najjar type I patients having a genetic defect in only exon A1 of the UGT1gene complex do not glucuronidate bilirubin. Our studies on the substrate specificity of the expressed human UGT1A4 protein may help clarify whether or not bilirubin is a substrate for UGT1A4. We find very low bilirubin glucuronidation rates for expressed human UGT1A4 (approximately 2 pmol/min/mg protein) when it is assayed using 5 mM UDP-glucuronic acid (Green and Tephly, 1996). Since human UGT1A4 catalyzes the N-glucuronidation of amines, it is possible that the very minor amount of bilirubin glucuronidation activity measured in vitro may be due to slightN-glucuronidation activity of the enzyme toward the pyrrole moieties of bilirubin or a breakdown product of bilirubin.
Human UGT1A3 was cloned from a human colon cDNA library, and the expressed protein was shown to catalyze the glucuronidation of estrone, 2-hydroxyesterone, and hydroxylated benzo[a]pyrene and 2-acetylaminofluorene metabolites (Mojarrabi et al., 1996). Human UGT1A3 is 93% identical in primary amino acid sequence to UGT1A4. Interestingly, despite this high degree of amino acid identity, the substrate specificities of the two expressed proteins are very different. Similar to UGT1A4, we have shown that expressed human UGT1A3 catalyzes glucuronidation of tertiary amines (Green et al., 1998). The reactivity of expressed UGT1A3 toward other xenobiotics and endobiotics is also very different from that of UGT1A4. While human UGT1A4 catalyzes glucuronidation at aliphatic hydroxyl groups (e.g. sapogenins, monoterpenoid alcohols, and progestins), these compounds are not substrates for UGT1A3. Expressed UGT1A3 catalyzes glucuronidation at aromatic hydroxyl groups (e.g.coumarins, anthraquinones, and flavonoids) and carboxylic acid moieties, but UGT1A4 does not. Thus amine compounds are the only common substrates for these proteins, which are >90% identical in primary amino acid sequence, suggesting that only a limited number of amino acids can define the aglycone binding site for UGT proteins.
Recently, two rabbit UGTs (tentatively identified as rabbit UGT1A4 and 1A7) have been shown to catalyze glucuronidation of tertiary amines to form quaternary ammonium–linked glucuronides (Bruck et al., 1997). While little is known about the reactivity of these expressed proteins with a wide range of tertiary amines, Bruck et al.(1997) have shown that the imipramine glucuronidation efficiency for expressed rabbit UGT1A4 is ten times that for expressed UGT1A7.
Possible Genetic Basis for Species Differences in Quaternary Ammonium Glucuronide Formation
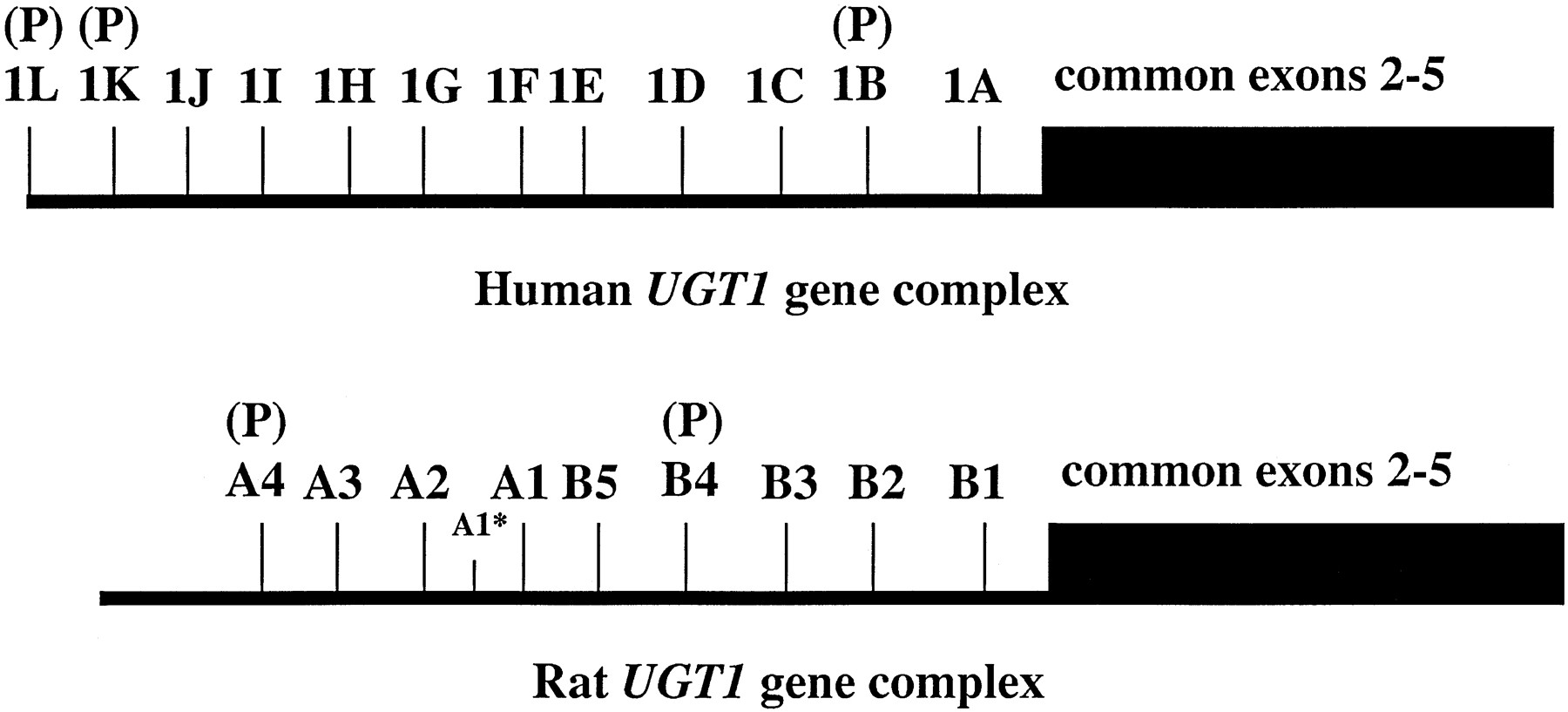
The rat and human UGT1 gene complexes are structurally similar (fig. 2). The humanUGT1 gene complex appears to encode for at least 12 unique gene products, each of which is formed by alternate splicing of multiple first exons with common exons 2–5 (Ritter et al., 1992; Cho et al., 1995). Similarly, at least nine unique first exons have been described for the rat UGT1 gene complex (Emi et al., 1995). Alternate splicing results in the production of mRNAs that code for proteins with unique amino acid sequences in the amine terminal portion of the protein, whereas the carboxyl portion of the protein is identical within the UGT1gene family. The first five exon 1s (human A1–F1 and rat B1–B5) have been described to code for proteins that are “bilirubin-like,” while the rest of the first exons code for UGTs that are “phenol-like” (Ritter et al., 1992; Emi et al., 1995). Human exon A1 and rat exon B1 both code for the amino terminal portion of UGT1A1. Expressed UGT1A1 proteins catalyze the glucuronidation of bilirubin, opioids, and other compounds, and we have recently shown that the rat and human UGT1A1 proteins are functionally similar and are likely to be orthologous enzymes (King et al., 1996). Likewise, human, rabbit, mouse, and rat UGT1A6 have been shown to be functionally similar and have been suggested to be orthologous (Lamb et al., 1994; Emi et al., 1996). Finally, the recent results of Green et al. (1998)and Bruck et al. (1997) show that expressed human and rabbit UGT1A4 proteins catalyze glucuronidation of tertiary amines to quaternary ammonium–linked glucuronides. These data suggest that theUGT1 gene structure and the function of the different proteins coded by this gene are highly conserved between different species.

Structures of the human and rat UGT1 gene complex.
The multiple first exons of the human [1A-1L(P)] and rat [B1–B5 and A1–A4(P)] UGT1 gene complexes are indicated. The drawing is not represented to scale and is not meant to imply accurate distances between the first exons. The common exons 2–5 are depicted as a block without the intervening introns. The human UGT1gene complex is compiled from Ritter et al. (1992) and Choet al. (1995), while the rat data is from Emi et al. (1995, 1996). A (P) indicates that the exon codes for a pseudo gene. In the rat gene complex, A1* indicates an extension of the first exon that codes for a 5′ nontranslated portion of the UGT1A6 mRNA that has been shown to have upstream transcriptional activation factors associated with it.
In the rat genome, UGT1 exon B2 (UGT1A2) codes the amine terminal portion of a second protein that catalyzes the glucuronidation of bilirubin (Sato et al., 1990; Sato et al., 1994). However, this exon encodes a premature stop codon and is, therefore, a pseudo-gene in the human genome (Ritter et al., 1992). Thus rats express two bilirubin UGTs, whereas humans express only one. These observations are consistent with the fact that patients with Crigler-Najjar type I syndrome, who have genetic defects only inUGT1 exon A1, are unable to glucuronidate bilirubin. These data also indicate that it is unlikely that the other “bilirubin-like” first exons code for proteins that catalyze the glucuronidation of bilirubin in humans. The amine terminal portion of human UGT1A4 is encoded by UGT1 exon D1. The corresponding rat enzyme is encoded by rat UGT1 exon B4. However, in the rat, exon B4 is a pseudo-gene and does not code for a full-length UGT protein. Therefore, it seems reasonable to suggest that the reason that rats do not glucuronidate many tertiary amines is that they cannot produce a functional UGT1A4 due to a genetic defect in first exon B4. The recent observation (Bruck et al., 1997) that expressed rabbit UGT1A4 also catalyzes glucuronidation of tertiary amines further supports this hypothesis.
At the present time, it is not known whether UGTs other than human UGT1A3 and UGT1A4 catalyze quaternary ammonium–linked glucuronide formation. A significant contribution of human UGT1A3 to the overall metabolism of primary and tertiary amines is somewhat doubtful, given that these compounds tend to exhibit higher apparentKm values, compared with UGT1A4 (Green et al., 1998) and that the expression of UGT1A3 is very low in human liver (Mojarrabi et al., 1996). Human UGT1A5 is >90% identical in primary amino acid sequence to human UGT1A3 and 1A4, and it is possible that this protein may also catalyze glucuronidation of amine substrates. However, to date, expression of UGT1A5 has not been demonstrated in a number of human tissues that have been examined (Strassburg et al., 1997). If human UGT1A7 has the ability to catalyze the formation of quaternary ammonium–linked glucuronides, as does the rabbit protein (Bruck et al., 1997), its contribution to the overall metabolism of tertiary amines is also probably insignificant because of the possible higher apparentKm values of the amine substrates and because human UGT1A7 is not expressed in liver (Strassburg et al., 1997). The observation that UGT1A7 is expressed in gastric tissue but is not expressed in human liver suggests that tissue-specific expression of UGT isoforms may have significance for the local metabolism of xenobiotics and endobiotics. Future studies are necessary to further characterize and identify the proteins responsible for amine glucuronidation in human tissues.
Footnotes
-
Send reprint requests to: Mitchell D. Green, M.S., Department of Pharmacology, 2-459 Bowen Science Building, The University of Iowa, Iowa City, IA 52242.
-
This work was supported by National Institutes of Health Grant GM 26221.
- Abbreviations used are::
- α-NA
- α-naphthylamine
- β-NA
- β-naphthylamine
- 4-ABP
- 4-aminobiphenyl
- UDP
- uridine diphosphate
- 2-ABP
- 2-aminobiphenyl
- OH
- hydroxy
- LA Wistar rats
- strain of Wistar rats deficient in UGT2B2
- HA Wistar rats
- strain of Wistar rats with normal UGT2B2
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}
{kind=link}