


Abstract
To study cytochrome P-450 (CYP) 2B6 contribution to methoxychlor metabolism within human liver microsomes and to initiate an investigation of CYP2B6 protein expression, we developed a polyclonal antibody targeted to a 20-residue peptide within that protein. The antibody was found to be highly sensitive and monospecific for CYP2B6 on immunoblots. Although many immunological studies have described the absence or low expression of CYP2B6 in human livers, in the present investigation, we have found this not to be the case. We immunoquantified CYP2B6 apoprotein expression in a panel of 28 livers and found concentrations ranging from 2 to 82 pmol/mg protein, with a mean value of 25 pmol/mg protein. Five livers (∼18%) displayed relatively high levels of CYP2B6 (>40 pmol/mg protein). There were no sex-related differences, although the highest level was observed in a 1-week postpartum donor given several medications. A marked diminution in variability was found in individuals aged 56 or older (n = 12), but there were no age-related trends in mean CYP2B6 content. We suggest that CYP2B6 represents a significant portion of total CYP in human liver. The exquisite sensitivity of this antibody (fmol quantities are detected easily on immunoblots) may explain our detection of CYP2B6 in 100% of livers versus its detection in a limited number of livers by certain other investigators. The antibody also was found to immunoinhibit CYP2B6-catalyzed N-demethylation of (S)-mephenytoin in human liver microsomes by 68 to 79%. The utility of this antibody for determining human liver microsomal CYP2B6 contribution to theortho-hydroxylation of methoxychlor was demonstrated.
The cytochrome P-450 (CYP)2 2B subfamily is among the most thoroughly investigated in the superfamily of CYP proteins (Lewis, 1998). Facile purification of certain members of this subfamily (particularly rat CYP2B1) from the livers of barbiturate-treated mammals has made CYP2B enzymes prototypes for diverse studies of CYP function. However, CYP2B6, the sole member of this subfamily expressed in humans (Yamano et al., 1989), has not been as extensively investigated. This is probably due to the unavailability or uncertainty of probes used to study this enzyme (Ekins et al., 1997) and the relative scarcity of human tissues. Additionally, several studies have reported that CYP2B6 protein is low or absent in the majority of individuals (Mimura et al., 1993; Shimada et al., 1994;Baker et al., 1995; Boobis et al., 1996; Imaoka et al., 1996; Edwards et al., 1998) (Table 1) and there is the general perception that CYP2B6 is a negligible component of total hepatic CYP. However, recent studies (Code et al., 1997; Tateishi et al., 1997; Ekins et al., 1998; Yang et al., 1998) have indicated that the incidence and quantity of CYP2B6 protein expression is far greater than reported in the above investigations. The incongruities do not appear to be related to ethnic, age, or sex differences within the populations studied but likely are attributable to the antibody probes or immunological techniques used to examine CYP2B6.
Summary of studies reporting expression and quantification of CYP2B6 protein in hepatic tissue
At least 40 substrates have been identified for recombinant CYP2B6, and some compounds have been reported for which microsomal CYP2B6 is a predominant catalyst (Heyn et al., 1996; Stevens et al., 1997).3 Our investigations of the metabolism of the proestrogenic pesticide methoxychlor have revealed that recombinant CYP2B6 catalyzes hydroxylation ortho to the methoxyl groups (Stresser and Kupfer, 1997). To study the CYP2B6 contribution to methoxychlor metabolism within human liver microsomes and to initiate an investigation of CYP2B6 protein expression, we developed a highly sensitive monospecific polyclonal antibody targeted to a 20-amino acid peptide. Using that antibody, we have quantified CYP2B6 apoprotein expression in a panel of 28 human livers and have examined the role of CYP2B6 in methoxychlor ortho-hydroxylation. With respect to immunoquantitation, our findings are similar to recent findings (Code et al., 1997; Ekins et al., 1998) in that we detected CYP2B6 expression in 100% of the livers. Additionally, we suggest that CYP2B6 represents a significant portion of total CYP in hepatic tissue. The exceptional sensitivity of this antibody (fmol quantities are easily detected) may explain our contradictory results versus those of others who have indicated the absence of this CYP in many individuals.
Materials and Methods
Chemicals and Enzymes.
Tween 20 was obtained from Fisher Scientific (Fair Lawn, NJ). 2,2′-Azino-bis-(3-ethylbenzothiazine-6-sulfonic acid), diammonium salt was purchased from Molecular Probes (Eugene, OR). Tris, glucose 6-phosphate, glucose 6-phosphate dehydrogenase, NADPH, EDTA disodium salt, p-coumaric acid, luminol, affinity-isolated goat anti-rabbit IgG (whole molecule) peroxidase conjugate, and caprylic acid were purchased from Sigma Chemical Co. (St. Louis, MO). Immobilon-P polyvinylidene difluoride transfer membranes were from Millipore Corp. (Bedford, MA). Nitrocellulose paper was obtained from Schleicher & Schuell (Keene, NH). Methoxychlor was purchased from Chem Service (West Chester, PA). Ammonium sulfate was from Schwarz/Mann (Spring Valley, NY). 1,1,1-Trichloro-2,2-bis-([3-3H]-4-hydroxyphenyl)ethane ([o-3H]bis-OH-M) (2.47 Ci/mmol) was custom-synthesized by Chemsyn Science Laboratories (Lenexa, KS). The purity of [o-3H]bis-OH-M was 95% by radio-HPLC and radio-thin-layer chromatography. [o-3H]Methoxychlor was prepared by methylation of [o-3H]bis-OH-M as described previously (Stresser et al., 1996). The final specific activity of the tritium-labeled substrate was 0.45 Ci/mmol for 1 μM incubations and 0.02 Ci/mmol for incubation with 25 μM substrate. Radioinert (S)-mephenytoin was obtained from Gentest Corporation (Woburn, MA), and [14C]-(S)-mephenytoin (56–58 mCi/mmol) was obtained from Amersham Life Sciences (Arlington Heights, IL). Human livers were acquired from the National Disease Research Interchange (Philadelphia, PA). Microsomes were prepared as described previously (Stresser et al., 1996). Additional human liver microsomes were purchased from the International Institute for the Advancement of Medicine (Exton, PA). Microsomes from human lymphoblastoid AHH-1 thymidine kinase +/− cell lines engineered to express single human CYP isoforms, or from the same cell line transfected with the vector only (controls) and microsomes prepared from baculovirus-infected insect cells (“Supersomes”) expressing CYP2B6, were purchased from Gentest Corporation. Recombinant CYP2C9, CYP2C19, and CYP2D6, purified fromEscherichia coli, were provided by Eric Johnson (The Scripps Institute, La Jolla, CA).
Antibody Production.
Peptide synthesis and antibody production were carried out by Research Genetics, Inc. (Huntsville, AL). A 20-amino acid peptide corresponding to residues 247 to 266 within CYP2B6 was synthesized by 9-fluorenylmethyloxycarbonyl chemistry on an Advanced Chemtech ACT396 peptide synthesizer, and the structure was confirmed by matrix-assisted laser desorption ionization/time of flight mass spectrometry. Conjugation at the N terminus to keyhole limpet hemocyanin was achieved using both 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride and glutaraldehyde (to improve chances of successful conjugation) in separate batches. The two batches were combined, diluted to 1 mg/ml in PBS, mixed at a ratio of 1:1 with adjuvant, and then emulsified with Freund’s complete adjuvant (for initial injection) or Freund’s incomplete adjuvant (for subsequent injections). Antibody was raised in four New Zealand White female rabbits that were approximately 12 weeks old. Animals were bled to obtain preimmune serum and subsequently injected intradermally with the keyhole limpet hemocyanin peptide, 250 μg initially (week 0), and then 100 μg subsequently at weeks 2, 6, and 8 and every 4 weeks thereafter. The IgG fraction was obtained from sera by fractionation with caprylic acid and ammonium sulfate (McKinney and Parkinson, 1987).
Enzyme Assays.
The extent of ortho-hydroxylation of [o-3H]methoxychlor by CYP2B6 was calculated by quantifying the release of3H2O using scintillation spectrometry as described previously (Stresser et al., 1996). Incubations were conducted in 1.5-ml microcentrifuge tubes in a 250-μl volume. Reaction mixtures contained 60 mM sodium phosphate buffer (pH 7.4), 10 mM magnesium chloride, various amounts of microsomal protein (1.15% potassium chloride was added to compensate for differences in volumes of microsomal protein solutions), 1 μM or 25 μM [o-3H]methoxychlor [specific activity (S.A.) 40,000–1,000,000 dpm/nmol] delivered in 2.5 μl of ethanol, and an NADPH-regenerating system [glucose 6-phosphate (2.5 mmol), NADPH (0.13 mmol), and glucose 6-phosphate dehydrogenase (0.5 U)]. After a preincubation period of 2–3 min at 37°C, the reaction was initiated with the addition of NADPH-regenerating system. The reaction was terminated at 10 min (liver microsomes) or 60 min (microsomal CYP2B6) by addition of 250 μl ice-cold 0.016 M calcium chloride followed by 250 μl dextran-coated charcoal (1% activated charcoal, 0.05% dextran, grade C, in 10 mM Tris-HCl, pH 8.0) to remove unreacted substrate and hydroxylated metabolites. After vigorous mixing and centrifugation, the supernatant was filtered through a 0.45-mm polyvinylidene difluoride syringe filter and a portion was analyzed by liquid scintillation spectrometry. The extent of CYP2B6-catalyzed (S)-mephenytoin N-demethylation was determined by incubating liver microsomes (0.6 mg/ml) with 100 μM [14C]-(S)-mephenytoin (S.A., 11,000–16,000 dpm/nmol) for 30 min. Product formation is linear with time under these conditions. Incubations were conducted in 1.5-ml microcentrifuge tubes in a 250-μl volume. Additionally, reaction mixtures contained 50 mM sodium phosphate buffer (pH 7.4), 1.3 mM NADP+, 3.3 mM glucose 6-phosphate, 0.4 U/ml glucose 6-phosphate dehydrogenase, and 3.3 mM magnesium chloride. To limit potential contribution of CYP2C9 in liver microsomes to (S)-mephenytoin N-demethylation (Ko et al., 1998), incubations also contained 10 μM sulfaphenazole, a potent and selective inhibitor of that enzyme. After the incubation period, the reaction was stopped by the addition of 50 μl of acetonitrile and placed on ice. The mixture was then subjected to centrifugation to sediment-precipitated protein. Reaction products in 150 μl of supernatant were resolved with a Nucleosil C185-μm 250 mm × 4.6 mm column (Supelco Inc., Bellefonte, PA) using a binary gradient consisting of 10% methanol (solvent A) and methanol (solvent B). Mobile phase was held at 30% B for 3 min and then brought to 70% B using a linear gradient over the next 14 min, before returning to initial conditions. The total solvent flow was 1 ml/min. Nirvanol eluted at 5.7 min and 4′-hydroxymephenytoin eluted at 6.2 min.
Enzyme-Linked Immunosorbent Assay.
The titer of the rabbit preimmune sera or antisera was determined at 0, 4, 8, and 10 weeks after the primary immunization. The peptide was coated to solid phase (1 μg/well) and binding of IgG was detected using biotinylated anti-rabbit IgG-horseradish peroxidase-streptavidin conjugate with 2,2′-azino-bis-(3-ethylbenzothiazine-6-sulfonic acid), diammonium salt as the substrate. Results are expressed as the reciprocal of the serum dilution that produces an absorbance of 0.2 at 405 nm.
Immunoinhibition.
Immunoinhibition of CYP2B6-catalyzed [14C]-(S)-mephenytoin N-demethylation or [o-3H]methoxychlorortho-hydroxylation was determined by preincubating increasing amounts of antipeptide IgG or normal IgG with CYP2B6 or liver microsomal protein. Potassium phosphate buffer (100 mM, pH 7.4) was added to bring the volume to 27 to 35 μl. After 20 min on ice, remaining components (additional buffer, magnesium chloride solution, or potassium chloride solution) of the incubation were added and the reaction was initiated and carried out as described above. When liver microsomal protein was used as an enzyme source in [o-3H]methoxychlorortho-hydroxylation immunoinhibition experiments, it was necessary to correct for background tritium in the supernatant (that was not removed by charcoal treatment). This was accomplished by dividing the supernatant into two equal portions. The radioactivity in one portion was determined and the second portion was evaporated under a stream of nitrogen at room temperature; the residue was dissolved in water, and the solution was evaporated again. The residue again was dissolved in water and subjected to scintillation spectrometry, and this value was subtracted from the value obtained in the other portion of the supernatant that was not evaporated.
Immunoblotting.
Samples of human liver microsomal protein were analyzed by SDS/polyacrylamide gel electrophoresis and immunoblotting. Liver microsomes were diluted to 3 mg/ml with 1.15% potassium chloride, and an equal volume of sample buffer (62.5 mM Tris-HCl, pH 6.8, 2% SDS, 10% glycerol, 0.1% bromophenol blue) was added. The mixture was boiled for 2 min and then placed on ice. Proteins (15 μg) were separated on the basis of size in 10% polyacrylamide gels by electrophoresis in a 30°C incubator. Proteins were transferred (400 mA for 1 h) to polyvinylidene difluoride membranes using the method of Towbin et al. (1979). After overnight drying, membrane-nonspecific binding sites were blocked with 0.5% milk (Carnation nonfat dry milk, obtained at a local supermarket) in Tris (10 mM, pH 7.5)-buffered 0.9% saline containing 0.05% Tween 20 (TBST) for 1 h. After two 5-min wash steps in TBST, the membrane was incubated for 1 h with a primary antibody solution composed of 6.3 μg/ml antipeptide IgG in a carrier of 0.1% BSA in TBST (equivalent to a 1:2000 dilution of whole serum). After four 5-min wash steps in TBST, the membrane was incubated with a secondary antibody solution containing goat anti-rabbit IgG peroxidase conjugate diluted 1:2000 in 0.5% milk/TBST for 1 h. After washing four times with TBST, immunoreactive proteins were visualized by immersing the blot in a solution prepared by mixing 10 μl of 68 mM p-coumaric acid (dissolved in dimethyl sulfoxide) and 5 μl of 30% H202 with 10 ml of 0.1 M Tris, pH 8.5, containing 1.25 mM luminol. A record of the light-emitting bands then was obtained by exposing the blots to light-sensitive film (Hyperfilm-ECL; Amersham Life Sciences, Arlington Heights, IL) for 2 to 20 s. A digital image of the film was obtained using a flat-bed scanner equipped with Deskscan software (Hewlett–Packard, Palo Alto, CA). The densities of the bands were measured by summing the pixel values after correction for background using ImageQuant software (Molecular Dynamics, Sunnyvale, CA). A standard curve was generated (GraphPad Prism software, version 2.0a; GraphPad, San Diego, CA) using spectrophotometrically quantified CYP2B6 in lymphoblast cell microsomes. For some experiments intended to demonstrate competition for epitopes of recombinant and microsomal CYP2B6 on immunoblots, the immunogen peptide was included at various concentrations in the primary antibody solution. Peptide was incubated with the antibody for 60 min at ambient temperature before probing the blots.
Results
Selection of Peptide.
We chose a CYP2B6-peptide sequence for use as an antigen based on several factors, including the extent of homology between other proteins, structural characteristics, and data from other investigators. Table 2 shows the sequence alignments of CYP102 with CYP2B6 and several members of the human CYP superfamily within the region selected as a peptide immunogen (Lewis, 1995, 1998). The most closely related human sequences, as indicated by a search of the Nonredundant GenBank CDS database, belonged to CYP2A6 and CYP2F1, which exhibited 50% identity among overlapping residues. This region exhibits high surface probability [based on the formula ofEmini et al. (1985)] and high hydrophilicity [based on the algorithm of Kyte and Doolittle (1982), averaged over a window of 7 residues], indicating that this portion of the protein likely would be on the exterior (Fig. 1). Alignment with bacterial CYP102 indicates that these 20 residues overlap a loop region between helices G and H in the three-dimensional model of mammalian CYP proposed by Lewis (1995). Finally, peptide antibodies against CYP2D6 [residues 254–273 (Cribb et al., 1995)] and CYP3A4 [residues 253–273 (Wang and Lu, 1997)] have been generated successfully using a similar region of the protein. Those antibodies were found to be useful for both immunoblotting and immunoinhibition studies.
Sequence alignment of CYP102 with CYP2B6 and various other human CYPs2-a

Surface probability plot (upper) and hydrophilicity plot (lower) of 20 residue sequence used to generate polyclonal antibodies against CYP2B6.
Characterization of Antibody.
All four animals (designated R1–R4) displayed high titers for the peptide 8 weeks after the primary injection (Table3) based on enzyme-linked immunosorbent assay analysis. Initial immunoblot analysis indicated that serum (1:2000 dilution) from all rabbits recognized CYP2B6, with serum from R1 giving the strongest signal. Serum from R1 and R2 inhibited 7-ethoxy-4-trifluromethyl coumarin O-deethylase activity and [o-3H]methoxychlor catalyzed by insect cell-expressed CYP2B6, whereas serum from R3 or R4 stimulated or showed no effect on these activities (data not shown). Based on these data, further studies were carried out with serum from R1 and R2. The specificity of the IgG fraction of R1 sera for CYP2B6 was examined by immunoblot analysis (Fig. 2). The antipeptide antibody recognized lymphoblast-expressed CYP2B6 and a single protein of identical molecular mass in human liver microsomes, but failed to recognize lymphoblast-expressed CYP1A1, CYP1A2, CYP1B1, CYP2A6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, or CYP3A4 or any protein in the lymphoblast cell line lacking cDNA-expressed CYP. The antibody also did not recognize recombinant CYP2C9, CYP2C19, or CYP2D6 purified from E. coli. The specificity for CYP2B6 was established further by demonstrating the ability of the immunogen peptide to competitively inhibit binding of antibody to lymphoblast CYP2B6 and the protein of identical Mr in human liver microsomes on immunoblots (Fig.3). Significant competition was observed at a peptide concentration of 4.4 nM (10 ng/ml). The total R1-IgG concentration was approximately 39.4 nM (6.3 μg/ml). Both R1-IgG and R2-IgG were found to be immunoinhibitory to recombinant and liver microsomal CYP2B6 (vide infra).
Antipeptide antibody titer of sera from four rabbits immunized with CYP2B6 peptide3-a

Immunoblot demonstrating the specificity of anti-CYP2B6 peptide antibody.
Lanes 1–11, microsomal protein (2.6–13.2 μg) from lymphoblastoid cells containing 0.5 pmol of the cDNA-expressed CYP indicated; lane 12, microsomal protein (10 μg) from lymphoblastoid cells transfected with a vector lacking a CYP insert; lane 13, liver microsomal protein (10 μg) from a 55-year-old Caucasian male; lane 14, identical with lane 5; lanes 15–17, purified CYP isoforms (2.5 pmol) obtained fromE. coli engineered to express the CYPs indicated.

Inhibition of antibody binding to CYP2B6 and to a protein in human liver microsomes by the immunogen peptide.
Microsomal protein from lymphoblast cells containing 0.3 pmol of CYP2B6 (left lanes) and 10 μg of liver microsomal protein from a 19-year-old Caucasian male (sample H3, right lanes) on immunoblots were probed with solutions of antipeptide antibody (6.3 μg/ml) containing 0-ng/ml, 1-ng/ml, or 10-ng/ml concentrations of the CYP2B6 peptide immunogen. Protein bands were visualized as described under Materials and Methods.
Immunoquantification of CYP2B6.
R1-IgG was used to quantify CYP2B6 apoprotein levels in a panel of human liver microsomes (Fig. 4). Information on the panel of donors is shown in Table4. All 28 livers contained detectable CYP2B6 enzyme, ranging from 2 to 82 pmol/mg protein, with a mean value of 25 pmol/mg protein (Fig. 5). Five livers (∼18%) displayed relatively high levels of CYP2B6 (>40 pmol/mg protein). There were no sex-related differences (females, 28.2 pmol/mg protein; males, 22.7 pmol/mg protein). A marked diminution in the variability of CYP2B6 expression was observed in individuals 56 years old (Fig. 6). The variance among 16 individuals aged 55 years or younger was 546, whereas that in 12 individuals aged 56 or older was 31. The small percentage of non-Caucasian samples (11%) precluded analysis of ethnic differences in CYP2B6 expression. There was no apparent association between clinical history and CYP2B6 expression, although it should be noted that the individual displaying the highest CYP2B6 expression (H51) was 1 week postpartum and had received phenytoin, dexamethasone, and other medications before death. To assess potential coregulation of CYP2B6 content with various catalytic activities, a correlation analysis was performed. CYP2B6 content was found to correlate significantly with ethoxycoumarin O-deethylase, coumarin hydroxylase, testosterone 6β-hydroxylase, and methoxychlorortho-hydroxylase activities in panels of human liver microsomes (Table 5).

Immunoblots demonstrating the presence and heterogeneous expression of CYP2B6 in a panel of 28 human liver samples.
Three different blots are shown. Lanes 1–9, hepatic microsomal protein (15 μg) from 9 donors; lanes 10–15, lymphoblast microsomal CYP2B6 standards (values are expressed in units of pmol)—the total protein loaded (0.8–15.4 μg) was adjusted to give the specified quantity of 2B6; lanes 16–24, hepatic microsomal protein (15 μg) from nine donors; lanes 25–30, lymphoblast microsomal CYP2B6 standards; lanes 31–40, hepatic microsomal protein (15 μg) from 10 donors; lane 41, 0.3 pmol of lymphoblast microsomal CYP2B6; lane 42, 0.5 pmol of lymphoblast microsomal CYP2A6; lane 43, lymphoblast microsomal protein (10 μg) from cells transfected with a vector lacking a CYP insert. Only the central portion of the blots are shown.
Donor information for liver samples

Distribution of CYP2B6 content determined by immunoblotting among liver microsomes from 28 individuals.
The quantity of CYP2B6 within each sample is shown at the top of each bar.
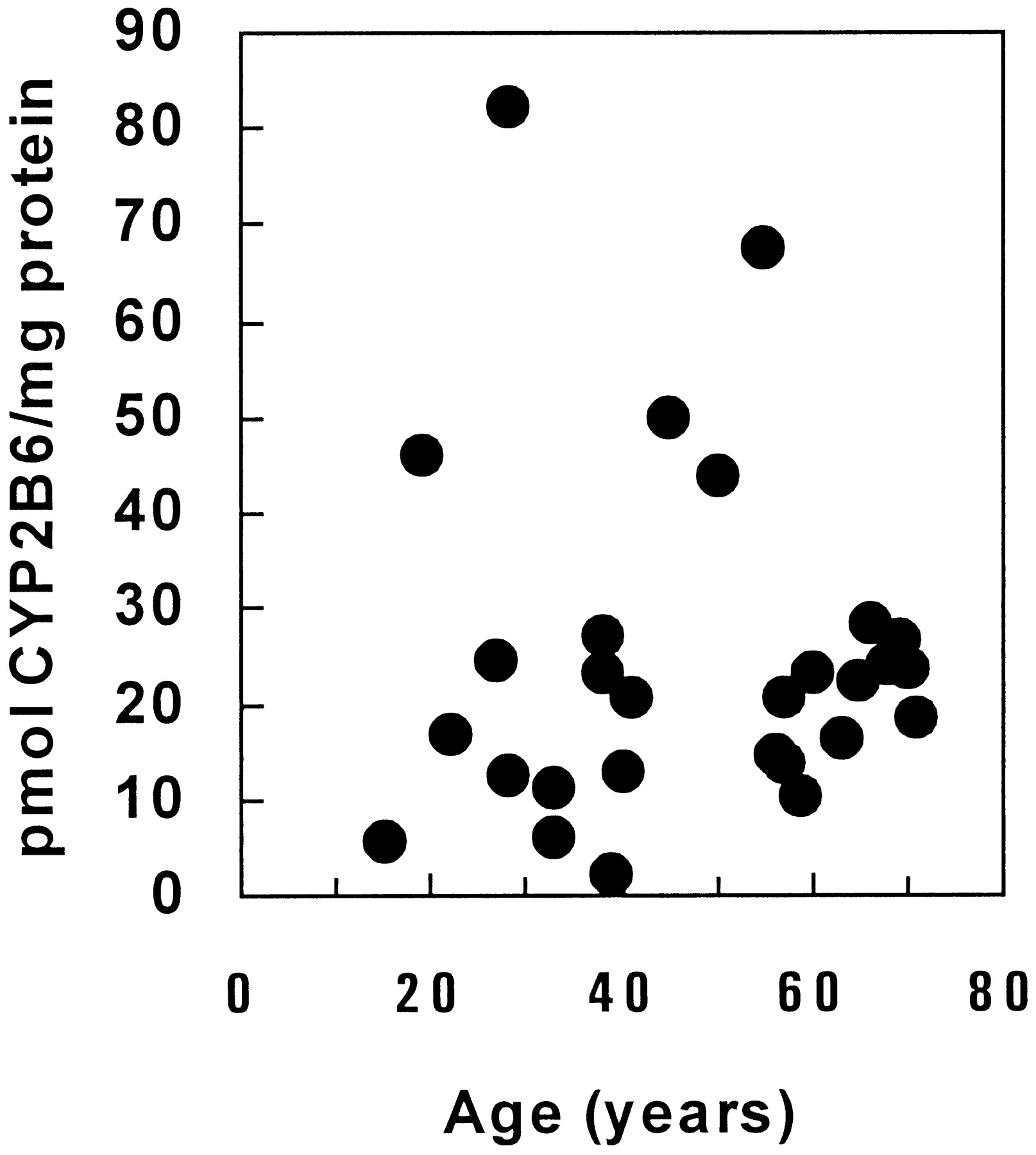
Effect of donor age on CYP2B6 content determined by immunoblotting among liver microsomes from 28 individuals.
Correlation of CYP2B6 content with various CYP-catalyzed activities in a panel of human liver microsomes
Immunoinhibition of [o-3H]Methoxychlorortho-Hydroxylase and [14C]-(S)-MephenytoinN-Demethylase Activity.
Figure 7 shows the effect of R1-IgG and R2-IgG on insect cell-expressed CYP2B6-catalyzed [o-3H]methoxychlorortho-hydroxylase activity. R1-IgG and R2-IgG preparations inhibited catalysis 91% and 82% relative to activity in the absence of IgG (control). Inexplicably, IgG prepared from normal, nonimmunized rabbits (n = 50) inhibited catalytic activity up to 35% relative to control values. To determine whether liver microsomal CYP2B6 also was subject to immunoinhibition, further experiments were carried out examining the effect of R1-IgG on the N-demethylation of (S)-mephenytoin catalyzed by human liver microsomes. The N-demethylation of (S)-mephenytoin is a highly selective marker for CYP2B6 catalytic activity (Heyn et al., 1996) with a limited contribution by CYP2C9 (Ko et al., 1998). To minimize potential contribution to this reaction by CYP2C9 (Ko et al., 1998), 10 μM sulfaphenazole, a specific inhibitor of that enzyme (Ono et al., 1996), was included in the incubations. Relative to activity in the absence of IgG, up to 68% inhibition of (S)-mephenytoin N-demethylation was observed (Fig. 8) in sample H3 human liver microsomes4(containing high levels of CYP2B6), demonstrating that the antibody substantially inhibits CYP2B6 native to liver. The specificity of the antibody for CYP2B6 immunoinhibition is supported by the observation that the antibody does not inhibit CYP2C19-catalyzed 4′-hydroxylation of (S)-mephenytoin (Fig. 8) nor bind to other CYPs on immunoblots (Fig. 2). In the same sample of human liver microsomes (e.g., H3), only 37% inhibition of methoxychlorortho-hydroxylation was found, indicating contribution by other enzymes to this reaction (Fig. 9).

Effect of R1-IgG and R2-IgG on insect cell-expressed CYP2B6-catalyzed ortho-hydroxylation of [o-3H]methoxychlor.
The concentration of methoxychlor in the reaction was 1 μM and that of CYP2B6 was 0.008 μM. The reaction was carried out for 60 min. Normal IgG was obtained from the serum of 50 nonimmunized rabbits. Meanortho-hydroxy metabolite formation in the absence of IgG was 3.6 pmol/60 min/pmol CYP2B6. Bars represent the S.D. of three (uninhibited) or range of two (in the presence of normal, R1, or R2 IgG) samples.

Effect of R1-IgG on human liver microsomal CYP2B6-catalyzed N-demethylation (N) and 4′-hydroxylation of (S)-mephenytoin (4′OH-M).
The concentration of (S)-mephenytoin in the reaction was 100 μM and that of liver microsomal protein (sample H3) was 0.6 mg/ml. The reaction was carried out for 30 min. Normal IgG was obtained from the serum of 50 nonimmunized rabbits. To minimize potential contribution by CYP2C9 to N-demethylation (Ko et al., 1998), experiments were carried out in the presence of 10 μM sulfaphenazole, a selective inhibitor of that enzyme. Mean Nirvanol formation in liver microsomes in the absence of IgG was 39 pmol/min/mg protein and that of 4′-hydroxy mephenytoin was 55 pmol/min/mg protein. Bars represent the S.D. of four (uninhibited) or three (in the presence of normal or R1 IgG) samples.

Effect of R1-IgG on human liver microsomal-catalyzed ortho-hydroxylation of [o-3H]methoxychlor.
The concentration of methoxychlor in the reaction was 25 μM and that of liver microsomal protein (sample H3) was 0.4 mg/ml. The reaction was carried out for 10 min. Normal IgG was obtained from the serum of 50 nonimmunized rabbits. Mean ortho-hydroxy metabolite formation in liver microsomes in the absence of IgG was 18.5 pmol/min/mg protein. Bars represent the S.D. of three (uninhibited) or range of two (in the presence of normal or R1 IgG) samples.
Discussion
The advantages of using peptides as antigens for antibody production have been reviewed (Harlow and Lane, 1988), and there are numerous examples of their use to prepare antibodies against CYP enzymes (Frey et al., 1985; Charnecki et al., 1995; Cribb et al., 1995;Richardson et al., 1997; Edwards et al., 1998). This study describes the utility of a 20-amino acid peptide aligning with amino acids 247 to 266 within CYP2B6 for production of a highly sensitive, monospecific antibody useful for immunoblotting and for immunoinhibition studies. It has become apparent recently that this sequence and others corresponding to a loop region between helices G and H in a three-dimensional model of mammalian CYPs based on bacterial CYP102 (Lewis, 1995) may constitute excellent choices as immunogens for producing antibodies to mammalian CYPs. A peptide antibody against CYP2C18 using amino acids 252 to 263 was useful in detecting recombinant CYP2C18 but did not recognize CYP2C8, CYP2C9, or CYP2C19 (Richardson et al., 1997). The so-called LKM-1 autoantibodies associated with autoimmune chronic active hepatitis (Manns et al., 1991) recognize an epitope on CYP2D6 within this loop region. Antibodies prepared against peptides composed of amino acids 254 to 273 within CYP2D6 (Cribb et al., 1995) and 253 to 273 within CYP3A4 (Wang and Lu, 1997) were found to be highly specific and inhibited the catalytic activity of their respective isoforms by more than 90%. Interestingly, antibodies raised against rat CYP2B1/2 sequence 265 to 276 (differing in only 1 amino acid from CYP2B6) was found to detect CYP2B6 monospecifically in only 6 of 30 livers. The findings that our antibody, too, is strongly inhibitory (more than 90% inhibition of recombinant CYP2B6 with R1-IgG and up to 79% in human liver microsomes) demonstrates its recognition of the catalytically active protein and suggests, as speculated by Wang and Lu (1997), the intriguing possibility that peptides aligning in this region may be universal choices for generation of highly immunoreactive CYP peptide antibodies. However, the observation that two of our preparations (from rabbits R3 and R4) yielded less-sensitive and noninhibitory antibodies underscores the potential for encountering idiosyncrasies in immune responses of outbred New Zealand rabbits or differences in immunogen presentation.
Using the IgG fraction from one of the rabbits (R1), we quantified CYP2B6 apoprotein in a panel of 28 livers. In contrast to most previous studies (see Table 1), we detected CYP2B6 in 100% of the livers. For example, CYP2B6 protein was detected in 2 of 10 livers (Baker et al., 1995), 12 of 50 livers (Mimura et al., 1993), 6 of 30 livers (Edwards et al., 1998), and 8 of 30 livers (Boobis et al., 1996). Our results are most similar to the data of Code et al. (1997) and Ekins et al. (1998), who quantified CYP2B6 in 17 of 17 livers and 19 of 19 livers, respectively. Expression of CYP2B6 protein in all samples is in concert with previous studies examining mRNA expression, where 15 of 15 (Miles et al., 1988) and 10 of 10 (Czerwinski et al., 1994) livers contained CYP2B6 message. The reasons why our results differ from those of most previous investigators are not certain but may be related to the sensitivity of our antibody, with which we could easily detect as low as 33 fmol of apo-CYP2B6 in one of the samples (H7). Indeed, the high affinity and specificity for the CYP2B6 epitope(s) was demonstrated by the observation that 4.4 nM the relevant peptide could inhibit binding to recombinant CYP2B6 and a microsomal protein exhibiting identicalMr on immunoblots. Our findings differ also in that the quantity of CYP2B6 in liver microsomes was found to be substantially higher than generally reported; among 14 livers in which the CYP was quantified, CYP2B6 represented 6% of the total. The discrepancy conceivably could be due to variations in immunological or quantitative methods used.
Nearly all of our samples were from an adult Caucasian population.Shimada et al. (1994) found that CYP2B6 protein was significantly higher in Caucasians compared with a Japanese population. Kirby et al. (1993) noted that CYP2B6 consistently was expressed in higher amounts in a Thai population compared with Caucasians, whereas Kim et al. (1997) observed that livers from Finnish cadavers contained approximately 3-fold-higher CYP2B6 than those from Chinese individuals. There appears to be some developmental differences associated with CYP2B6 expression. Tateishi et al. (1997) found that CYP2B6 was significantly lower in a population that was gestational week 37 to 1 year old compared with a population aged 1 year or older. In our study, there appeared to be no age-related trends in mean levels of expression, although the variation of CYP2B6 expression was considerably less in samples from individuals 56 years old or older. It is conceivable that the aged population is less responsive to putative inducers or suppressors of CYP2B6. Indeed, aged male rats were found to be less responsive to the enzyme-inducing effects of phenobarbital (Groen et al., 1994). There were no sex-related differences or obvious correlations with social/clinical history, although the highest level of CYP2B6 was observed in a 1-week postpartum donor given several medications.
We found that CYP2B6 content correlated significantly with several CYP-catalyzed activities. Correlation with ethoxycoumarinO-deethylase activity suggests a greater involvement of CYP2B6 than was estimated previously (Yamazaki et al., 1996). The observation of a correlation between CYP2A6-catalyzed 7-hydroxylation of coumarin and CYP2B6 content is consistent with previous studies demonstrating correlation of CYP2A6 and CYP2B6 protein expression (Forrester et al., 1992) or catalytic activities (Heyn et al., 1996;Code et al., 1997). The significant correlation of CYP2B6 content with CYP3A4-catalyzed testosterone 6β-hydroxylase activity in the present study is in agreement with previous observations that liver containing high CYP2B6 levels also contained elevated CYP3A4 (Mimura et al., 1993), but this is not a universal observation (Code et al., 1997).
We observed previously that cDNA-expressed CYP2B6 could catalyze theortho-hydroxylation of methoxychlor (Stresser and Kupfer, 1997), a reaction diagnostic of rodent CYP2B catalytic activity (Stresser et al., 1996). This reaction was not catalyzed by eight additional cDNA-expressed human CYPs, with the exception of CYP1A2, which catalyzed this reaction at about 50% the rate of CYP2B6 (Stresser and Kupfer, 1997). Further exploration revealed that [3H]methoxychlorortho-hydroxylation, but not CYP1A2-catalyzed ethoxyresorufin O-deethylation or phenacetinO-deethylation, correlated (r2 = 0.69, n = 18) with CYP2B6 immunoquantified with an antibody commercially available (described in Code et al., 1997; also, unpublished observations). These data suggested the utility of methoxychlor as a probe to measure CYP2B6 catalytic activity in human liver microsomes. We have found since that CYP3A4 contributes to total ortho-hydroxylase activity in liver microsomes, but only after O-demethylation of methoxychlor by CYP1A2 and CYP2C19 (Stresser and Kupfer, 1998a,b). Purified CYP2C19 reconstituted withl-α-dilauroyl-phosphatidyl-choline and NADPH CYP oxido-reductase also was found to catalyzeortho-hydroxylation of [3H]methoxychlor (Stresser and Kupfer, 1998a). In the present study, we observed significant correlation of [3H]methoxychlor ortho-hydroxylation with CYP2B6 content immunoquantified using R1-IgG, confirming our earlier finding using the commercially available antibody. However, R1-IgG was found to immunoinhibit no greater than 37% of methoxychlorortho-hydroxylation in a sample of human liver microsomes with relatively high CYP2B6 content. In the same sample, R1-IgG strongly inhibited the N-demethylation of (S)-mephenytoin, a reaction catalyzed selectively by CYP2B6 (Heyn et al., 1996). These results indicate that methoxychlor ortho-hydroxylation is not a pure indicator of microsomal CYP2B6 catalytic activity.
Compared with other human CYPs, the development of acceptable substrate (Heyn et al., 1996) and immunoinhibitory (Yang et al., 1998) probes to study the role of CYP2B6 in the metabolism of drugs and other xenobiotics has been gradual (Ekins et al., 1997). The current investigation has led to the development of an antipeptide antibody that is highly suitable for CYP2B6 immunoblotting studies; it has been found to give a more reliable and much more sensitive signal (E. L. Code and C. L. Crespi, personal communication) than the monoclonal antibody used in recent reports (Ekins et al., 1998; Yang et al., 1998). In addition, the antibody is immunoinhibitory to both recombinant and liver microsomal CYP2B6. All investigations of CYP2B6, including the present study, have shown expression of the enzyme to be highly variable. The reasons for this are not certain, but ethnicity appears to play a role (Kirby et al., 1993; Shimada et al., 1994; Kim et al., 1997). Our findings that all 28 livers contained detectable quantities of CYP2B6 apoprotein strongly suggest an absence of a prevalent null phenotype, in contrast to most earlier studies. This conclusion is supported by studies of CYP2B6 mRNA expression (Miles et al., 1988; Czerwinski et al., 1994). Further studies are required to elucidate whether there is a modality to CYP2B6 expression as indicated here and in a previous study (Yang et al., 1998). We suggest that CYP2B6 is a significant component of hepatic human CYP. Because several biochemical tools are now available to study metabolism by this enzyme (although a selective CYP2B6 chemical inhibitor has not yet been found), the role of this isoform in the metabolism of xenobiotics can be established with greater certainty.
Acknowledgments
We thank sincerely Drs. Eric Johnson and Toby Richardson (The Scripps Research Institute, La Jolla, CA) for providing the purified recombinant CYPs. We also thank Regina Wang and Anthony Lu (Merck Research Laboratories, Rahway, NJ) for advice on the purification of IgG from serum and Ms. Christine Hoogasian for assistance with polyacrylamide gel electrophoresis. We thank Mahesh Lachyankar of our department and Charles Crespi (Gentest Corporation, Woburn, MA) for helpful comments and discussions. We also thank Dr. Crespi and Gentest Corporation for use of certain materials and facilities to perform the [14C]-(S)-mephenytoinN-demethylase assay.
Footnotes
-
Send reprint requests to: Dr. David Kupfer, Department of Pharmacology and Molecular Toxicology, University of Massachusetts Medical Center, 55 Lake Ave. North, Worcester, MA 01655. E-mail:David.Kupfer{at}ummed.edu
-
↵1 Current address: Gentest Corporation, 6 Henshaw Street, Woburn, MA 01801.
-
The project described was supported by Grants ES00834 and ES05737 from the National Institute of Environmental Health Sciences, National Institutes of Health. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the National Institute of Environmental Health Sciences, National Institutes of Health.
-
↵3 The conversion of buproprion to its major active metabolite, hydroxybuproprion, is catalyzed primarily by CYP2B6 in liver microsomes (E. P. Goodale and J. A. Ascher, personal communication).
-
↵4 In another sample of human liver microsomes, R1-IgG was found to inhibit (S)-mephenytoin N-demethylation by 79% (data not shown).
- Abbreviations used are::
- CYP
- cytochrome P-450
- TBST
- Tris (10 mM, pH 7.5)-buffered 0.9% saline containing 0.05% Tween 20
- R1-IgG and R2-IgG
- IgG fraction obtained from rabbits 1 and 2, respectively
- Received October 28, 1998.
- Accepted January 4, 1999.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}