


Abstract
Although there are numerous studies of glucuronidation of endogenous compounds, information on the glucuronidation of fatty acids is lacking. In the present studies, both linoleic acid (LA) and its biologically active oxidized derivatives, 13-hydroxyoctadecadienoic acid (13-HODE) and 13-oxooctadecadienoic acid (13-OXO), have been shown to be effective substrates for human liver UDP-glucuronosyltransferases (UGT) and recombinant UGT2B7. LA (carboxyl glucuronide) and 13-OXO (carboxyl glucuronide, unproven) were actively glucuronidated by human liver microsomes (HLM) and human recombinant UGT2B7 with similar activities, in the range of 2 nmol/mg · min. The hydroxyl derivative of LA, 13-HODE, was glucuronidated at both the hydroxyl and carboxyl functions with carboxyl glucuronidation predominating (ratio of COOH/OH, 2:1). For all substrates, the Kmfor formation of the carboxyl-linked glucuronide was in the range of 100 to 200 μM while that for the hydroxyl-linked glucuronide was somewhat lower (>100 μM). This is the first demonstration of glucuronidation of LA and its oxidized derivatives, 13-HODE and 13-OXO, by HLM and recombinant UGT2B7.
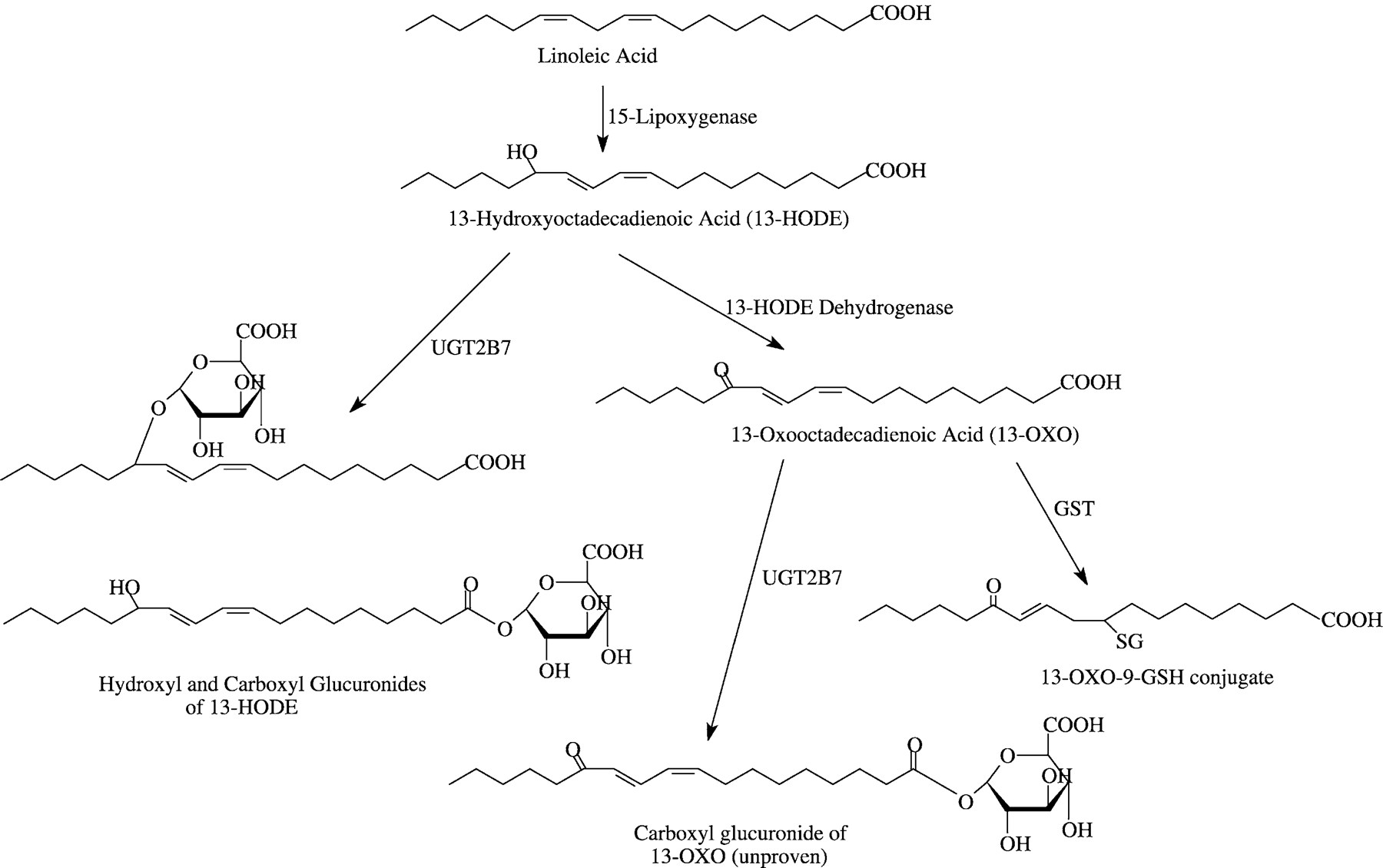
There are multiple oxidative metabolites of linoleic acid (LA1). In the present report, we examine glucuronidation of two naturally occurring metabolites of LA, 13-hydroxyoctadecadienoic acid (13-HODE) and 13-oxooctadecadienoic acid (13-OXO). The metabolic pathways and structures involved are shown in Fig. 1. 13-HODE has a number of different cellular effects: as a chemorepellant in endothelial cells, preventing platelet and cellular adhesion (Buchanan et al., 1985, 1991, 1998; Haas et al., 1988; Honn et al., 1992); as a modulator of cellular proliferation (Mani et al., 1998; Shureiqi et al., 1999); and as an inhibitor of protein kinase C isoforms (Liu et al., 1995;Pongracz and Lord, 1999), possibly as a result of 13-HODE incorporation into phospholipids (Cho and Ziboh, 1994). Thus, 13-HODE is implicated in cellular signaling. Little information is available on the biological activity of 13-OXO, aside from its formation of adducts with thiol groups of cellular proteins, conjugation with glutathione (Bull et al., 1996; Blackburn et al., 1997), and a correlation between cellular differentiation and the activity of 13-HODE dehydrogenase, the enzyme responsible for the formation of 13-OXO (Bronstein and Bull, 1993; Bull et al., 1993; Silverman et al., 1996).

Structures and metabolism of LA, 13-HODE, and 13-OXO.
In contrast, the 9,10- and 12,13-LA-diols have been reported to be cytotoxic (Moran et al., 1997). Glucuronides of these compounds have been recovered in urine of patients with generalized peroxisomal disorders (Street et al., 1996), and recently, we have demonstrated the in vitro glucuronidation of both diols by human liver microsomes (Jude et al., 1999) and human recombinant UGT2B7 (Jude et al., 2001). Interestingly, the addition of the two hydroxyl groups to the LA molecule directs the glucuronidation exclusively toward the hydroxyl groups at high levels of activity.
We show here, for the first time, that LA and both 13-HODE and 13-OXO are glucuronidated by human liver microsomes and human recombinant UGT2B7. LA is glucuronidated at the only available functional group, the carboxyl function, while 13-HODE is actively glucuronidated at both the hydroxyl and carboxyl functions with significantly higher activity (2-fold) toward the latter. With 13-OXO, which has the carboxyl function available for glucuronidation, it is assumed that the carboxyl glucuronide has been formed; however, this structure has not been verified. The exact type of glucuronide that is formed with 13-OXO is currently under investigation.
Materials and Methods
[14C]UDP-glucuronic acid ([14C]UDPGlcUA) and [14C]linoleic acid were from PerkinElmer Life Sciences (Boston, MA). Unlabeled UDPGlcUA, LA, and all other chemicals for enzymatic assays were from Sigma (St. Louis, MO). Solvents for TLC and HPLC were all Optima HPLC grade (Fisher Scientific, Pittsburgh, PA).
Synthesis of 13-HODE and 1-[14C]13-HODE.
LA was oxygenated by soybean lipoxygenase to prepare 13-hydroperoxy-(z,e)-9,11-octadecadienoic acid (13-HPODE) as described by Funk et al. (1976). The 13-HPODE was reduced with sodium borohydride to yield 13-HODE (Funk et al., 1976; Bull et al., 1984). The 13-HODE was purified by silicic acid chromatography, and the purity was analyzed by normal phase HPLC. Radiolabeled 1-[14C]13-HODE was prepared in the same manner using 1-[14C]LA.
Preparation of 13-OXO and 1-[14C]13-OXO.
LA was oxygenated by soybean lipoxygenase to prepare 13-HPODE, which was directly dehydrated by acetyl chloride in pyridine to yield 13-OXO (Porter and Wujek, 1987). 1-[14C]13-OXO was prepared as described above for 1-[14C]13-HODE. Both 13-OXO and 1-[14C]13-OXO were purified by silicic acid chromatography and analyzed by normal phase HPLC. The compounds were at least 95% pure with an isomeric composition of 88% (z,e) and 12% (z,z).
Conditions for HPLC.
Normal phase HPLC conditions were as follows: a 3-mm × 30-mm PerkinElmer pecosphere column was eluted isocratically with 0.7% isopropyl alcohol and 0.1% acetic acid in hexane, at a flow rate of 1.0 ml/min. Analytes were detected at 255 nm.
Human Liver Microsomes and Human Recombinant UGT2B7.
Human liver microsomes (HLM) were prepared from a sample of transplantable liver (HLM18, a 13-year-old female who died of brain damage) received from Drs. F. Kuipers and Roel Vonk (Academic Hospital, Groningen, The Netherlands) or from a liver obtained from an organ donor (HLM2, a 67-year-old female who died from a stroke) by transplant surgeons at The University Hospital (Little Rock, AR) according to a protocol approved by the Human Research Advisory Committee of the University of Arkansas for Medical Sciences. Livers were histologically normal. Human UGT2B7 was expressed in HK293 cells, a gift from Dr. T. Tephly (Department of Pharmacology, University of Iowa, Iowa City, IA), as previously described (Coffman et al., 1997). A membrane fraction enriched in UGT2B7 was prepared as described by Battaglia et al. (1994), and aliquots were stored at −80°C until used.
Enzyme Assays.
Initially, UGT activities toward 13-HODE and 13-OXO were assayed using [14C]UDPGlcUA as the sugar donor. Subsequently, [14C]13-HODE and [14C]13-OXO were used as substrates with unlabeled UDPGlcUA as described in detail previously for bile acids and steroids (Radominska-Pyrek et al., 1986; Radominska-Pandya et al., 1998). Briefly, substrates (100 μM for the standard assay) were solubilized in micelles with Brij 58 and preincubated with human liver microsomes or UGT2B7 membrane fraction (50 μg of protein) in 100 mM HEPES-NaOH, pH 7.4, with 5 mM MgCl2 and 5 mM saccharolactone at room temperature for 10 min. Reactions were started by the addition of [14C]UDPGlcUA or UDPGlcUA (4 mM) and incubated at 37°C for 10 min. Reactions were stopped by the addition of ethanol, aliquots were applied to TLC plates [silica gel; Si250-PA (19C), J.T. Baker, Phillipsburg, NJ], and glucuronidated products and the unreacted substrate were separated by two successive developments in chloroform-methanol-glacial acetic acid-water (65:25:2:4, v/v). Radioactive compounds were localized on TLC plates by autoradiography at −80°C. Zones corresponding to the glucuronide bands were scraped into scintillation vials, and radioactivity was measured by liquid scintillation counting (LS5000TD, Beckman Instruments, Fullerton, CA).
Initial identification of the position of glucuronidation (hydroxyl or carboxyl group) was determined by TLC. Two sets of reaction mixtures were prepared. Following incubation, reactions were stopped as usual, and one set was left on ice while the other set was brought to 0.1 N NaOH by addition of 1 μl of 8 N NaOH and incubated at room temperature for 1 h. Both sets of samples were then chromatographed as described above. Following development, the plates were subjected to autoradiography. Since carboxyl-linked glucuronides are alkali-labile, comparison of the recovery of product in each set of samples (hydroxyl- and carboxyl-linked glucuronides from the control samples; only hydroxyl-linked glucuronides from hydrolyzed samples) allows for a tentative assignment of the position of glucuronidation.
For further structural characterization, the products were isolated by TLC and analyzed by fast atom bombardment (FAB) mass spectrometry at the Michigan State University Mass Spectrometry Facility (East Lansing, MI). Spectra were obtained in both glycerol and nitrobenzyl alcohol matrices. Some samples were methylated with trimethylsilyldiazomethane prior to TLC.
Kinetic Analysis.
For analysis of the kinetics of the reactions, enzymatic activities were determined as described above using human liver microsomes or UGT2B7 with varying concentrations of substrates (25–500 μM) at a fixed concentration of UDPGlcUA (4 mM). The data were analyzed using EnzymeKinetics (Trinity Software, Compton, NH), and the kinetic parameters, Km andVmax, were determined.
Results and Discussion
Enzymatic Activity.
Enzymatic glucuronidation of LA, 13-HODE, and 13-OXO by human liver microsomes and recombinant UGT2B7 is summarized in Table1, and the autoradiograph of a typical TLC separation of the products of glucuronidation of the three substrates by HLM18 is shown in Fig. 2. The glucuronidation is shown to be specific by the lack of any reaction in the absence of UDPGlcUA. LA and 13-OXO were effectively glucuronidated by both microsomes and recombinant UGT2B7 resulting in single products. However, in both cases, microsomal glucuronidation activity was higher than that of UGT2B7. Two products result from incubation of 13-HODE with either microsomes or UGT2B7 and, here also, the microsomal activity was roughly twice that of UGT2B7. With both proteins, the more polar product (Rf = 0.31) was biosynthesized at approximately twice the level of the second, less polar (Rf = 0.45) conjugate.
Glucuronidation of LA, 13-HODE, and 13-OXO by human liver microsomes and recombinant UGT2B7
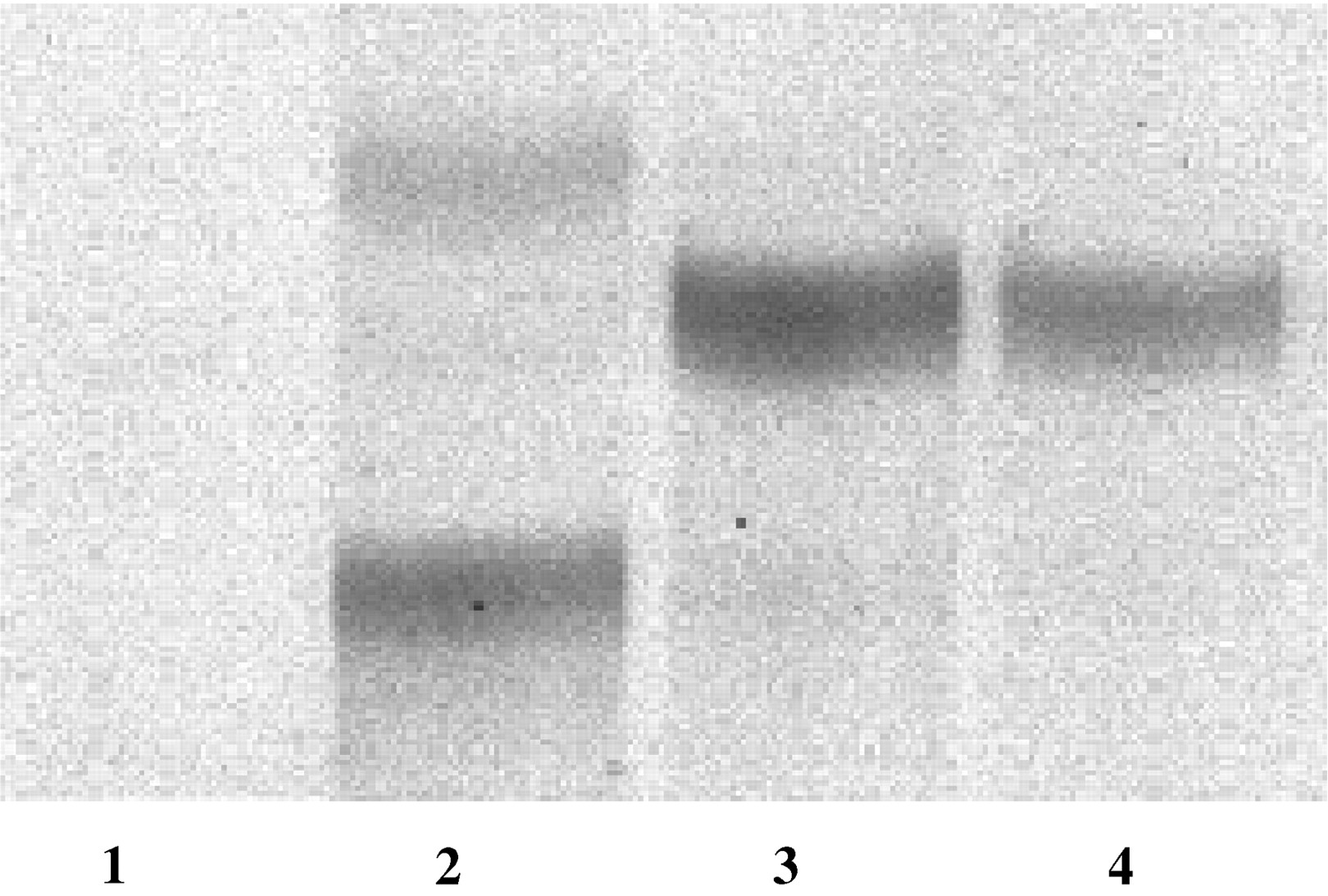
Autoradiograph of a representative TLC separation of the glucuronides of LA, 13-HODE, and 13-OXO.
Substrates (100 μM) were incubated with HLM18 (50 μg of protein) and [14C]UDPGlcUA (4 mM) and subjected to TLC as described in the text. Samples are as follows: 1, control incubation run in the absence of substrate. 2, 13-HODE; lower band, carboxyl-linked glucuronide; upper band, hydroxyl-linked glucuronide. 3, 13-OXO, carboxyl glucuronide, structure unproven. 4, LA, carboxyl-linked glucuronide.
A preliminary assignment of the position of glucuronidation was accomplished by subjecting the products to alkaline hydrolysis. Since acyl (carboxyl-linked) glucuronides are alkali-labile whereas hydroxyl-linked glucuronides are not, the carboxyl-linked products should be detected in unhydrolyzed but not in hydrolyzed samples following TLC. Using this technique, tentative identifications were as follows: LA and 13-HODE, Rf = 0.31, carboxyl-linked glucuronides; 13-HODE, Rf = 0.45, hydroxyl-linked glucuronide. The results of alkaline hydrolysis of the 13-OXO glucuronide suggest a carboxyl-linked glucuronide.
The FAB mass spectrum of the carboxyl-linked glucuronide of 13-HODE showed ions at m/z 472 for the molecular ion of the underivatized parent compound as well as an ion atm/z 486 representing the methyl ester. The FAB mass spectrum of the hydroxyl-linked glucuronide showed ions atm/z 472 for the underivatized compound and an ion at m/z 542 for the permethylated compound. These mass spectral observations are consistent with the results of the alkaline hydrolysis experiments.
The FAB mass spectrum for the 13-OXO glucuronide showed ions of equal intensity at m/z 469 and 471 (M+ + H and M+ − H) in glycerol and an ion of 470 in nitrobenzyl alcohol. From these results, it is clear that only a monoglucuronide was present, as there were no ions indicative of diglucuronides. However, the data were not sufficient to assign the position of glucuronidation for 13-OXO.
There is little information available on glucuronidation of fatty acids. Human recombinant UGT1A3 and UGT1A8 have been shown to glucuronidate decanoic (80 and 19 pmol/mg · min for UGT1A3 and UGT1A8, respectively) and dodecanoic (114 and 12 pmol/mg · min for UGT1A3 and UGT1A8, respectively) acids (Green et al., 1998; Cheng et al., 1999). Glucuronides of linoleic acid diols have been found in the urine of patients with peroxisomal disorders (Street et al., 1996) and 20-hydroxyeicosatetraenoic acid glucuronide has been identified in normal human urine (Prakash et al., 1992). Our studies provide the first data on the in vitro glucuronidation of these biologically important fatty acids. These are novel substrates for recombinant UGT2B7, which is expressed in liver, kidney, and intestine and is known to glucuronidate the carboxyl or hydroxyl functions of various drugs and endogenous compounds (Radominska-Pandya et al., 1999). The data presented here demonstrate that UGT2B7 is capable of glucuronidating both functional groups present in a single compound (13-HODE). Similar data have been presented previously for hydroxylated derivatives of bile acids (Gall et al., 1999) and retinoic acid (Samokyszyn et al., 2000).
Kinetic Analysis.
Analysis of apparent kinetic parameters showed that theKm values for glucuronidation of LA by HLM and UGT2B7 were similar, as were those for carboxyl glucuronidation of 13-HODE (Table 2). However, the latter were approximately 2.5 times higher than the former. The Km values for hydroxyl glucuronidation of 13-HODE by the two enzyme sources were similar but were approximately 5 times lower than those for formation of the carboxyl glucuronide of 13-HODE and 2 times lower than theKm values for linoleic acid glucuronidation. The position of glucuronidation for 13-OXO has not been definitively assigned, but the Km for its glucuronidation is more in line with the formation of the carboxyl glucuronide of 13-HODE than with formation of a hydroxyl glucuronide. Despite the differences in Km andVmax among the three compounds, the efficiencies of most of the reactions, as indicated byVmax/Km, are similar, suggesting that all three compounds are natural substrates for UGTs.
Apparent kinetic parameters for glucuronidation of LA, 13-HODE, and 13-OXO by human liver microsomes
Summary
13-HODE is a major oxidative metabolite of LA, which has a range of bioactive properties. Depending on the situation, the effect of this activity can be positive or negative. For instance, 13-HODE has been shown to prevent cellular adhesion to endothelial cells both in vitro and in vivo (Honn et al., 1992; Bertomeu et al., 1993). In contrast, 13-HODE activation of peroxisome proliferator-activated receptor-γ has been implicated in the development of atherosclerosis (Chen et al., 1999; Vamecq and Latruffe, 1999).
Glucuronidation can act to limit the biological activity of a compound, and this may be its role with LA and its oxidized derivatives. Since LA and 13-HODE are ligands for nuclear receptors (Mazzachi, 1992; Chen et al., 1999; Vamecq and Latruffe, 1999) and UGT2B7 has been identified in human liver nuclei (A. Radominska-Pandya, I. D. Pokrovskaya, G. Chen, J. M. Little, A. R. Jude, and P. J. Czernik, unpublished data), another role for UGTs may be to modulate the availability of ligands for nuclear receptors, as has been suggested by Nebert (1991)for drug-metabolizing enzymes in general. The activity of nuclear UGTs toward fatty acid substrates is presently being investigated.
Footnotes
-
Send reprint requests to: Anna Radominska-Pandya, Department of Biochemistry and Biophysics, University of Arkansas for Medical Sciences, 4301 W. Markham St., Slot 516, Little Rock, AR 72205. E-mail: RadominskaAnna{at}exchange.uams.edu
-
This research was supported in part by National Institutes of Health Grants DK56226 and DK49715 (to A.R.-P.) and CA76420 (to A.W.B.).
- Abbreviations used are::
- LA
- linoleic acid
- 13-HODE
- 13-hydroxyoctadecadienoic acid
- 13-OXO
- 13-oxooctadecadienoic acid
- UGT
- UDP-glucuronosyltransferase
- UDPGlcUA
- UDP-glucuronic acid
- 13-HPODE
- 13-hydroperoxyoctadecadienoic acid
- z
- cis
- e
- trans
- HLM
- human liver microsomes
- TLC
- thin-layer chromatography
- HPLC
- high-performance liquid chromatography
- FAB
- fast atom bombardment
- Rf
- retardation factor
- Received July 7, 2000.
- Accepted January 2, 2001.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}