


Abstract
We established stable HEK293 cell lines expressing double isoforms, UGT1A1 and UGT1A9, UGT1A4 and UGT1A9, or UGT1A6 and UGT1A9, as well as stable cell lines expressing each single isoform. To analyze the protein-protein interaction between the UGT1As, we investigated the thermal stability and resistance to detergent. UGT1A9 uniquely demonstrated thermal stability, which was enhanced in the presence of UDP-glucuronic acid (>90% of control), and resistance to detergent. Interestingly, UGT1A1, UGT1A4, and UGT1A6 acquired thermal stability and resistance to detergent by the coexpression of UGT1A9. An immunoprecipitation assay revealed that UGT1A6 and UGT1A9 interact in the double expression system. Using the single expression systems, it was confirmed that estradiol 3-O-glucuronide, imipramine N-glucuronide, serotonin O-glucuronide, and propofol O-glucuronide formations are specific for UGT1A1, UGT1A4, UGT1A6, and UGT1A9, respectively. By kinetic analyses, we found that the coexpressed UGT1A9 significantly affected the kinetics of estradiol 3-O-glucuronide formation (decreased Vmax), imipramine N-glucuronide formation (increased Km and Vmax), and serotonin O-glucuronide formation (decreased Vmax) catalyzed by UGT1A1, UGT1A4, and UGT1A6, respectively. On the other hand, the coexpressed UGT1A1 increased Km and decreased the Vmax of the propofol O-glucuronide formation catalyzed by UGT1A9. The coexpressed UGT1A4 and UGT1A6 also increased the Vmax of the propofol Oglucuronide formation by UGT1A9. This is the first study showing that human UGT1A isoforms interact with other isoforms to change the enzymatic characteristics.
UDP-glucuronosyltransferases (UGTs) are a family of membrane-bound enzymes that catalyze the conjugation of endogenous and exogenous compounds with UDP-glucuronic acid (UDPGA) (Dutton, 1980). Human UGTs are classified into two subfamilies, UGT1A and UGT2B, on the basis of evolutionary divergence (Mackenzie et al., 2005). Characterization of genomic DNA clones encoding the UGT1 gene has shown that the UGT1 locus comprises multiple first exons that encode isoform-specific sequences and a single set of commonly used exons 2 to 5 that encodes the same sequence of all UGT1 isoforms (Bosma et al., 1992; Ritter et al., 1992). The C-terminal domain with very high homology is directly involved in UDPGA binding, whereas the N-terminal halves contain the aglycone binding site (Mackenzie, 1990). Among the UGT1A subfamily, UGT1A1, UGT1A4, UGT1A6, and UGT1A9 are expressed in human liver (Strassburg et al., 1999; Tukey and Strassburg, 2000).
UGTs have been reported to form homo- or hetero-oligomers as follows. Matern et al. (1982) suggested for the first time, on the basis of gel permeation chromatography, that UGTs existed as oligomers in rat liver microsomes. Radiation inactivation analyses revealed that UGT existed as oligomers composed of one to four subunits (Peters et al., 1984; Gschaidmeier and Bock, 1994). Moreover, cross-linking studies confirmed possible hetero-oligomerizations between UGT1As and UGT2B1 (Ikushiro et al., 1997). In addition, several experiments in recent years have verified the formation of homo- or heterooligomers (Ishii et al., 2001; Kurkela et al., 2004). However, the functional significance of the oligomerization is largely unknown. In the present study, we established stable HEK293 cell lines expressing simultaneously UGT1A1 and UGT1A9, UGT1A4 and UGT1A9, or UGT1A6 and UGT1A9 to investigate the effects of the heterodimerization on the enzymatic activities. Estradiol 3-O-glucuronide, imipramine N-glucuronide, serotonin O-glucuronide, and propofol O-glucuronide formations were determined as specific activities for UGT1A1, UGT1A4, UGT1A6, and UGT1A9, respectively. Using these marker activities, the effects of coexpression of other UGT1A isoforms on thermal stability, resistance to detergent, and the kinetic parameters were investigated.
Materials and Methods
Chemicals and Reagents. UDPGA, alamethicin, estradiol, estradiol 3-O-glucuronide, 4-methylumbelliferone (4-MU), and 4-methylumbelliferone O-glucuronide were purchased from Sigma-Aldrich (St. Louis, MO). Imipramine hydrochloride, serotonin, trifluoperazine dihydrochloride, and G418 were purchased from Wako Pure Chemicals (Osaka, Japan). 7-Hydroxy-4-trifluoromethylcoumarin (7-HFC) was obtained from Daiichi Pure Chemicals (Tokyo, Japan). Propofol was kindly supplied by AstraZeneca (London, UK). Recombinant human UGT1A1, UGT1A4, UGT1A6, and UGT1A9 expressed in baculovirus-infected insect cells (Supersomes) and rabbit anti-human UGT1A polyclonal antibody were obtained from BD Gentest (Woburn, MA). Rabbit anti-human UGT1A6 peptide polyclonal antibody was prepared as described previously (Ikushiro et al., 2006). Primers were commercially synthesized at Hokkaido System Sciences (Sapporo, Japan). All other chemicals and solvents were of analytical grade or the highest grade commercially available.
Isolation of Human UGT1A cDNA and Construction of Expression Vectors. Human UGT1A1 (accession number NM000463), UGT1A4 (NM007120). UGT1A6 (NM001072), and UGT1A9 (NM021027) cDNAs were prepared by a reverse transcription-polymerase chain reaction technique using total RNA from human liver. The sequences of the primers are shown in Table 1. After an initial denaturation at 94°C for 5 min, amplification was performed by denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 90 s for 35 cycles, followed by a final extension at 72°C for 5 min. The polymerase chain reaction products were subcloned into pTARGET Mammalian Expression Vector (Promega, Madison, WI) and the DNA sequences of the inserts were determined using a Thermo Sequenase Cy5.5 Dye Terminator Cycle Sequencing kit (GE Healthcare Bio-Sciences, Piscataway, NJ) with a Long-Read Tower DNA sequencer (GE Healthcare Bio-Sciences).
Primers used in this study
Stable Expression of UGT1A Isoforms in HEK293 Cells. HEK293 (American Type Culture Collection, Rockville, MD) cells were grown in Dulbecco's modified Eagle's medium containing 4.5 g/liter glucose, 10 mM HEPES, and 10% fetal bovine serum with 5% CO2 at 37°C. The cells in six-well plates were transfected with 2 μg of UGT expression vector using Lipofectamine (Invitrogen, Carlsbad, CA). For the double expression systems, each expression vector was cotransfected at the ratio of 1:1. Stable transfectants were selected in medium containing 800 μg/ml of G418 and several clones were isolated. The UGT expression levels were determined by immunoblot analysis as described below. As the single expression systems, clones with the highest content of each UGT1A isoform were selected for the assays.
Preparation of Cell Homogenate. HEK293 cells expressing single or double UGT1A isoforms were suspended in Tris-buffered saline [25 mM Tris-HCl buffer (pH 7.4), 138 mM NaCl, and 2.7 mM KCl] and disrupted by freeze-thawing three times according to the method reported by Ren et al. (2000). Then, the suspensions were homogenized with a Teflon-glass homogenizer for 10 strokes. The protein concentrations were determined according to Bradford (1976).
Immunoblot Analysis. The UGT protein levels were determined by immunoblot analysis. Cell homogenates (2–10 μg) from HEK293 cells or UGT Supersomes (0.2 μg) were boiled for 5 min in Laemmli sample buffer containing 2-mercaptoethanol and separated on a 10% polyacrylamide gel. The separated proteins were electrotransferred onto the polyvinylidene difluoride membrane Immobilon-P (Millipore, Bedford, MA). The membrane was blocked in 3% nonfat dry milk in phosphate-buffered saline (PBS) containing 0.1% Tween 20 at room temperature for 12 h. The membranes were incubated with rabbit anti-human UGT1A polyclonal antibody (1:500, PBS) for 5 h at room temperature. Biotinylated anti-rabbit IgG and a VECTASTAIN ABC kit (Vector Laboratories, Burlingame, CA) were used for diaminobenzidine staining. The densities of the bands were determined using an ImageQuant (GE Healthcare Bio-Sciences). Because the anti-human UGT1A antibody recognizes the conserved C-terminal region of all human UGT1A isoforms, it reacts with all UGT1A isoforms as previously reported (Malfatti and Felton, 2004). The expression level of UGT1A was defined on the basis of a standard curve using the UGT1A1 single expression system (1 unit/1 mg of cell homogenates).
Enzyme Assays. Estradiol 3-O-glucuronide formation was determined according to the method of Fisher et al. (2000) with slight modifications. Briefly, a typical incubation mixture (200 μl of total volume) contained 50 mM Tris-HCl (pH 7.4), 10 mM MgCl2, 2 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates and 2 to 100 μM estradiol. The reaction was initiated by the addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 60 min, the reaction was terminated by addition of 100 μl of cold acetonitrile including 6% acetic acid. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
Imipramine N-glucuronide formation was determined according to the method of Nakajima et al. (2002) with slight modifications. Briefly, a typical incubation mixture (100 μl of total volume) contained 50 mM Tris-HCl buffer, pH 7.4, 5 mM MgCl2, 5 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates and 0.1 to 5 mM imipramine. The reaction was initiated by the addition of UDPGA, following a 3-min preincubation at 37°C. After incubation at 37°C for 60 min, the reaction was terminated by addition of 100 μl of cold acetonitrile. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
Serotonin O-glucuronide formation was determined according to the method of Krishnaswamy et al. (2003) with slight modifications. Briefly, a typical incubation mixture (100 μl of total volume) contained 25 mM phosphate buffer (pH 7.5), 2.5 mM MgCl2, 5 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates, and 0.3 to 20 mM serotonin. The reaction was initiated by the addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 30 min, the reaction was terminated by addition of 100 μl of cold acetonitrile. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
Propofol O-glucuronide formation was determined according to the method of Shimizu et al. (2003) with slight modifications. Briefly, a typical incubation mixture (200 μl of total volume) contained 50 mM potassium phosphate buffer (pH 7.4), 10 mM MgCl2, 5 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates, and 10 μM to 2 mM propofol. The reaction was initiated by addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 30 min, the reaction was terminated by addition of 200 μl of cold acetonitrile. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
4-MU O-glucuronide formation was determined according to the method of Uchaipichat et al. (2004) with slight modifications. Briefly, a typical incubation mixture (200 μl of total volume) contained 50 mM Tris-HCl buffer, pH 7.4, 10 mM MgCl2, 2.5 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates, and 30 μM 4-MU. The reaction was initiated by addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 15 min, the reaction was terminated by addition of 100 μl of cold methanol. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
Trifluoperazine N-glucuronide formation was determined according to the method of Uchaipichat et al. (2006) with slight modifications. Briefly, a typical incubation mixture (100 μl of total volume) contained 50 mM Tris-HCl buffer, pH 7.4, 10 mM MgCl2, 2.5 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates, and 50 μM trifluoperazine. The reaction was initiated by addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 15 min, the reaction was terminated by addition of 100 μl of cold acetonitrile including 6% acetic acid. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
7-HFC O-glucuronide formation was determined according to the method of Ghosal et al. (2004) with slight modifications. Briefly, a typical incubation mixture (200 μl of total volume) contained 50 mM Tris-HCl buffer, pH 7.4, 10 mM MgCl2, 2 mM UDPGA, 25 μg/ml alamethicin, 0.25 mg/ml total cell homogenates, and 50 μM 7-HFC. The reaction was initiated by addition of UDPGA after a 3-min preincubation at 37°C. After incubation at 37°C for 15 min, the reaction was terminated by addition of 100 μl of cold acetonitrile including 6% acetic acid. After removal of the protein by centrifugation at 15,000 rpm for 5 min, a 20-μl portion of the sample was subjected to HPLC.
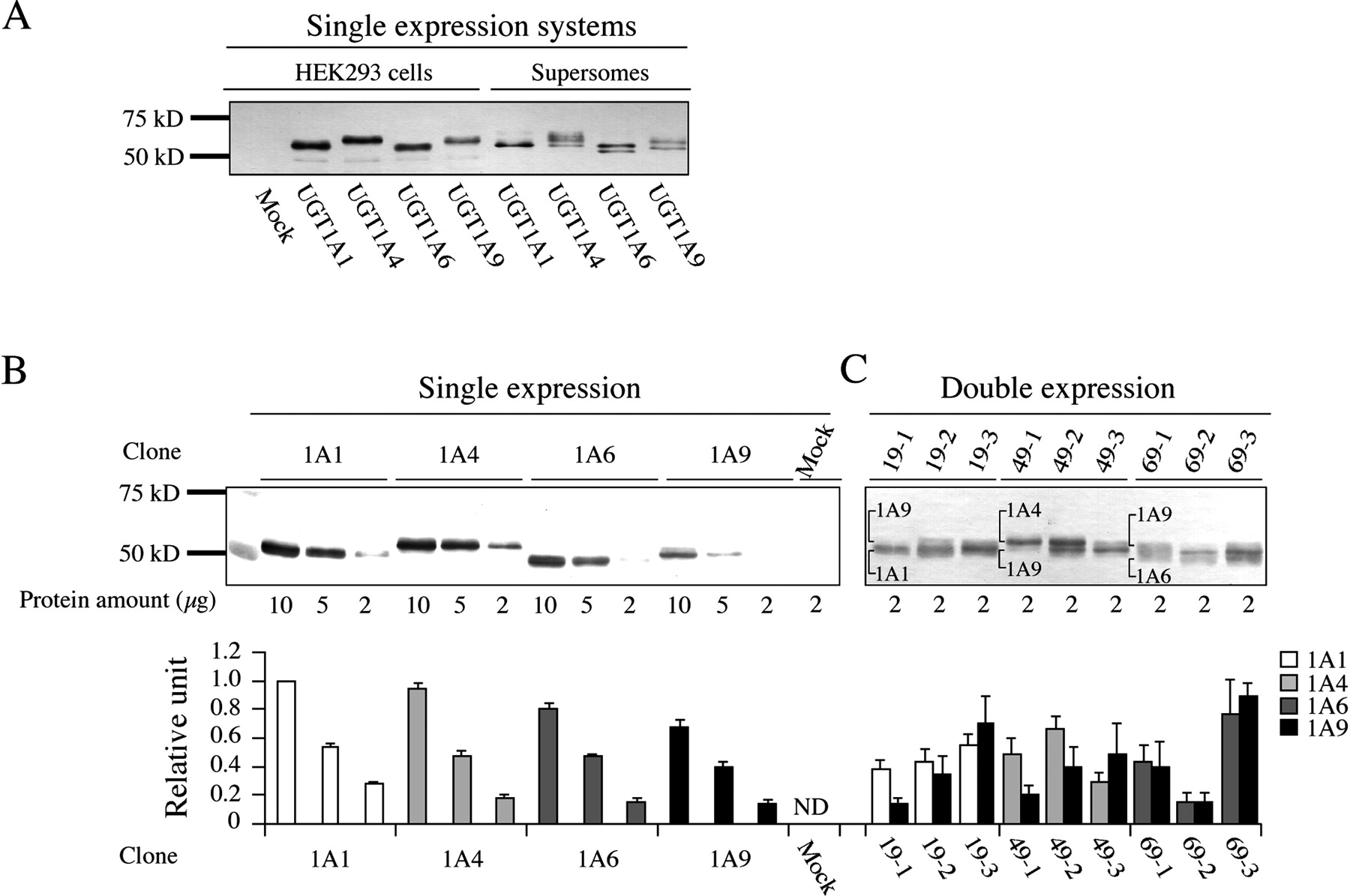
Immunoblot analysis of recombinant human UGT1As expressed in HEK293 cells and baculovirus-infected insect cells (Supersomes) using a rabbit antihuman UGT1A antibody. A, 2 μg of total cell homogenates from the HEK293 cells and 0.2 μg of Supersomes were separated by electrophoresis using 10% SDS-polyacrylamide gel electrophoresis. The lower UGT1A levels in HEK293 cells compared to the Supersomes were due to the usage of the total cell homogenates instead of microsomes. B, various contents of HEK293 cell homogenates from single expression systems of UGT1A were applied to make the standard curves. C, the expression levels of UGT1A isoforms in double expression systems were defined based on a standard curve using the UGT1A1 single expression system (1 unit/1 mg of cell homogenates). Each column is the mean ± S.D. of three independent determinations. ND, not detected.
Quantification of estradiol 3-O-glucuronide or 4-MU O-glucuronide was performed by comparing the HPLC peak height to that of the authentic standard. For the quantification of the other glucuronides, the eluate from the HPLC column containing each glucuronide was collected, and a part of the eluate was hydrolyzed with NaOH at 75°C for 30 min (Hawes, 1998). The hydrolyzed glucuronides were quantified using HPLC by comparison of peak heights with those of the external standard curve of the substrates.
Kinetic parameters were estimated from the fitted curve using a computer program (KaleidaGraph, Synergy Software, Reading, PA) designed for nonlinear regression analysis. The following equations were used:
Michaelis-Menten equation: 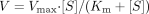 Hill equation
Hill equation  Substrate inhibition equation
Substrate inhibition equation 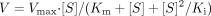 where V is the velocity of the reaction, S is the substrate concentration, Km is the Michaelis-Menten constant, Vmax is the maximum velocity, S50 is the substrate concentration showing the half-Vmax, n is Hill coefficient, and Ki is the substrate inhibition constant. Data are expressed as the means ± S.D. of three independent determinations.
where V is the velocity of the reaction, S is the substrate concentration, Km is the Michaelis-Menten constant, Vmax is the maximum velocity, S50 is the substrate concentration showing the half-Vmax, n is Hill coefficient, and Ki is the substrate inhibition constant. Data are expressed as the means ± S.D. of three independent determinations.
Effects of Heat Treatment and Detergent on Enzymatic Activities. To investigate the effect of heat treatment, the reaction mixtures containing the cell homogenate were treated at 37, 42, 47, 52, and 57°C for 15 min in the presence or absence of UDPGA or substrate. Immediately, the enzymatic activities were determined as described above with the incubation at 37°C for 15 min. To investigate the effect of the detergent, the incubation mixture was incubated with 0.02% (v/v) Triton X-100 for 15 min on ice. Immediately, the enzymatic activities were determined as described above with the incubation at 37°C for 15 min.
Immunoprecipitation. Rabbit anti-UGT1A6 peptide antibody was conjugated with protein A-Sepharose CL-4B (GE Healthcare Bio-Sciences) using dimethylpimelimidate in PBS containing 0.2 M triethanolamine. The beads were washed with 50 mM ethanolamine in PBS for 5 min and 1 M glycine (pH 3.0) for 20 min. The beads were resuspended in lysis buffer (2 mM EDTA, 1% Triton X-100, 0.1% SDS in PBS). Cell homogenates were lysed with the lysis buffer at 4°C for 60 min. After centrifugation at 13,000g for 30 min at 4°C, the supernatants (0.2 mg) were incubated with the beads at 4°C for 12 h. The beads were washed three times with the lysis buffer. The bound proteins were eluted with 2 M guanidine hydrochloride for 2 h at room temperature. After guanidine hydrochloride was removed using a PAGEprep Advance Protein Clean-up and Enrichment kit (Pierce Biotechnology, Rockford, IL), the immunoprecipitates were subjected to immunoblot analysis.
Statistical Analyses. Statistical significances of the effects of the temperature and kinetic parameters were determined by analysis of variance followed by Dunnett's test. Statistical significance of the effects of detergent was determined by unpaired Student's t test. A value of P < 0.05 was considered statistically significant.
Results
Establishment of Single and Double Expression Systems of UGT1A in HEK293 Cells. To establish the stable cell lines expressing single isoforms of UGT1A1, UGT1A4, UGT1A6, or UGT1A9, we isolated five clones for each isoform. Immunoblot analysis revealed that the expression levels of UGT varied among the clones (data not shown). We selected the clones with the highest UGT1A protein levels for the subsequent analyses. As shown in Fig. 1A, the HEK293 single expression systems of each UGT1A isoform showed a single band at 50 to 55 kDa. In contrast, recombinant UGT1A4, UGT1A6, and UGT1A9 in baculovirus-infected insect cells (Supersomes) showed multiple bands, possibly owing to variability in the glycosylation (Malfatti and Felton, 2004). Based on the UGT1A1 level in HEK293 cells of 1.00 unit/mg protein, UGT1A4, UGT1A6, and UGT1A9 levels were estimated to be 0.94, 0.81, and 0.68 unit/mg protein, respectively (Fig. 1B). The minimal S.D. values of the protein levels (Table 2) revealed the reproducibility of quantification of UGT1As.
Expression levels of UGT1A1, UGT1A4, UGT1A6, and UGT1A9 in single and double expression systems
Data are means ± S.D. of three independent determinations.
Because of the different mobilities, UGT1A1 and UGT1A9, UGT1A4 and UGT1A9, or UGT1A6 and UGT1A9 in each double expression system were separately detected in immunoblot analysis (Fig. 1C). The expression levels of UGT1As in the double expression systems ranged from 0.14 to 0.90 unit/mg protein (Table 2). Although the two expression vectors were transfected with equal content, the ratios of the expression level of UGT1A1 to UGT1A9 in each clone ranged from 0.8 to 2.9, those of UGT1A4 to UGT1A9 ranged from 0.6 to 2.5, and those of UGT1A6 to UGT1A9 ranged from 0.9 to 1.0. The quantification of each UGT1A isoform was reproducible as shown in Table 2.
Thermal Instability of UGT1A1, UGT1A4, UGT1A6, and UGT1A9 in Single Expression Systems. Because the pretreatment of cell homogenate at 37°C for 15 min did not affect any enzymatic activity (data not shown), the activities after a 15-min pretreatment at 37°C were used as controls. Estradiol 3-O-glucuronide formation by the single expression system of UGT1A1 was slightly decreased by heat treatment at 42°C (91% of control) and prominently decreased by treatment at 47°C (17% of control) in the absence of substrate or UDPGA (Fig. 2A). Treatment at 52 and 57°C completely abolished the activity. UDPGA significantly restored the thermal instability at 47°C, yielding 67% of control. In contrast, estradiol had little effect on the restoration. Imipramine N-glucuronide formation by the single expression system of UGT1A4 was decreased by heat treatment at 47°C (80% of control) and at 52°C (25% of control). Treatment at 57°C completely abolished the activity. UDPGA significantly restored the thermal instability at 52°C, yielding 64% of control. The effects of imipramine on the thermal instability could not be determined, because imipramine was unstable when heated. Serotonin O-glucuronide formation by the single expression system of UGT1A6 was decreased by heat treatment at 42°C (73% of control) and at 52°C (31% of control) (Fig. 2C). UDPGA highly restored the activity at 42°C (100% of control), 47°C (94% of control), and 52°C (61% of control). However, serotonin had no effect on the thermal instability. Propofol O-glucuronide formation by the single expression system of UGT1A9 was relatively stable for the heat treatment, yielding 89% of control at 47°C, 53% of control at 52°C, and 40% of control at 57°C (Fig. 2D). UDPGA prominently restored the activity at all temperatures (>91% of control). Because propofol was unstable in heat, the effects of propofol on thermal instability could not be determined.
To verify the thermal instability profile of UGT1As and the effects of UDPGA or substrates, 4-MU O-glucuronide formation by UGT1A1 (Fig. 2E), UGT1A6 (Fig. 2G), and UGT1A9 (Fig. 2H) and trifluoperazine N-glucuronide formation by UGT1A4 (Fig. 2F) were measured. Consequently, it was demonstrated that UGT1A9 is more thermally stable compared with UGT1A1, UGT1A4, and UGT1A6 in the absence of UDPGA and that UDPGA could restore the enzymatic activities. Because trifluoperazine was also unstable in heat, we could not determine the effects of the substrate on the thermal instability of UGT1A4.
We expanded this experiment for recombinant UGTs in baculovirus-infected insect cells (Supersomes) to compare the differences in hosts or expression systems (Fig. 3). The thermal instability properties of UGT1A1, UGT1A4, UGT1A6, and UGT1A9 Supersomes were almost the same as those of UGT1As in HEK293 cells. In addition, it was clearly demonstrated that the enzymatic activities of UGT1A3, UGT1A7, UGT1A8, and UGT1A10 were also prominently decreased at 52 and 57°C.
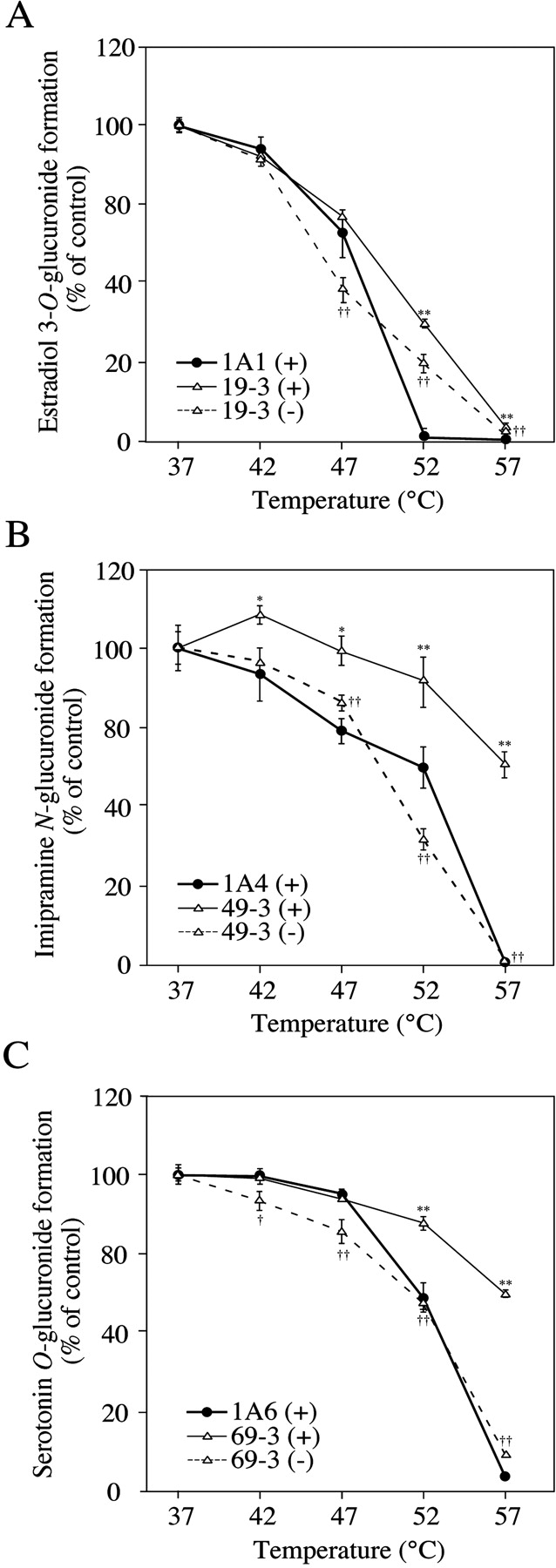
Effects of Coexpression of UGT1A9 on Thermal Instability of UGT1A1, UGT1A4, and UGT1A6. We investigated the thermal instability of UGTs in the double expression systems. In the clone 19-3, estradiol 3-O-glucuronide formation catalyzed by UGT1A1 was restored by the coexpression of UGT1A9 (30% of control in clone 19-3 versus 1% of control in the UGT1A1 single expression system at 52°C) (Fig. 4A). However, the restoration was weak in the absence of UDPGA (21% of control). Clones 19-1 and 19-2 showed similar profiles as the single expression system of UGT1A1 (data not shown). The imipramine N-glucuronide formation catalyzed by UGT1A4 was restored by the coexpression of UGT1A9 (Fig. 4B). In clone 49-3, the activity was restored to 61% of control at 57°C, whereas the activity of the single expression system was only 1% of control. However, the restoration was not observed in the absence of UDPGA. Clones 49-1 and 49-2 also showed the restoration, but the extents were lower than that in 49-3 (data not shown). In clone 69-3, serotonin O-glucuronide formation catalyzed by UGT1A6 was restored by the coexpression of UGT1A9 (58% of control in clone 69-3 versus 3% of control in the UGT1A6 single expression system at 57°C) (Fig. 4C). However, the restoration was weak in the absence of UDPGA (8% of control). Clones 69-1 and 69-2 also showed a profile similar to that for clone 69-3 (data not shown). On the other hand, coexpression of UGT1A1, UGT1A4, and UGT1A6 did not show any effect on the thermal stability of UGT1A9 measured by propofol O-glucuronide formation (data not shown).
Effects of Coexpression of UGT1A9 on the Inhibitory Effects of Triton X-100. The effects of Triton X-100 on the enzymatic activities of UGT1A in the single expression systems were investigated (Fig. 5). The activities of UGT1A1, UGT1A4, and UGT1A6 were abolished to <3% of the control activities by 0.02% Triton X-100, whereas the activity of UGT1A9 showed 66% of the control activity. Thus, UGT1A9 was resistant to the treatment with Triton X-100. We then examined the effects of Triton X-100 on the UGT1A activities in the double expression systems. Clones 19-3, 49-3, and 69-3 showed significantly higher residual activity compared with their single expression systems, suggesting that the coexpression of UGT1A9 could increase the resistance to Triton X-100. Clones 69-1 and 69-2 also showed the higher residual activities, although clones 19-1, 19-2, 49-1, and 49-2 showed residual activities similar to those of the single expression systems of UGT1A1 and UGT1A4 (data not shown). The differences in the three clones from each cotransfection were similar to those for the effects on thermal stability, depending on the relative expression ratio. The effects of Triton X-100 on UGT1A9 activity were not changed by the coexpression of UGT1A1, UGT1A4, or UGT1A6. These results suggested that the coexpression of UGT1A9 altered the properties of UGT1A1, UGT1A4, and UGT1A6.
Coimmunoprecipitation of UGT1A6-UGT1A9. To investigate the direct association between UGT1A9 and the other UGT1A isoforms, immunoprecipitation analysis was performed (Fig. 6). It was confirmed that the anti-human UGT1A6 antibody could immunoprecipitate UGT1A6 protein. When the cell homogenate from the double expression system of UGT1A6 and UGT1A9 was used, both UGT1A6 and UGT1A9 were immunoprecipitated. However, mixing of the single expression systems of UGT1A6 and UGT1A9 could not yield the immunoprecipitant of UGT1A9 (data not shown). The results suggested that UGT1A6 directly interacted with UGT1A9 in a common membrane. Unfortunately, the association between UGT1A1/UGT1A9 and UGT1A4/UGT1A9 could not be investigated, because antibodies for immunoprecipitation are not available.

Effect of heat treatment on the enzymatic activities of recombinant UGT1A1, UGT1A4, UGT1A6, and UGT1A9 in single expression systems. The reaction mixtures were treated at 37, 42, 47, 52, or 57°C for 15 min in the absence (•) or presence of the substrate (○) or UDPGA (▵). Estradiol 3-O-glucuronide (A), imipramine N-glucuronide (B), serotonin O-glucuronide (C), propofol O-glucuronide (D), 4-MU O-glucuronide (E, G, and H), and trifluoperazine N-glucuronide (F) formations were measured at the substrate concentrations of 50 μM, 1 mM, 5 mM, 200 μM, 30 μM, and 50 μM, respectively. The residual activity was determined by comparing with the activity when preincubated at 37°C. Data are the means ± S.D. of three independent determinations. *, P < 0.05; **, P < 0.01, compared with the activities treated in the absence of both the substrate and UDPGA at each temperature.
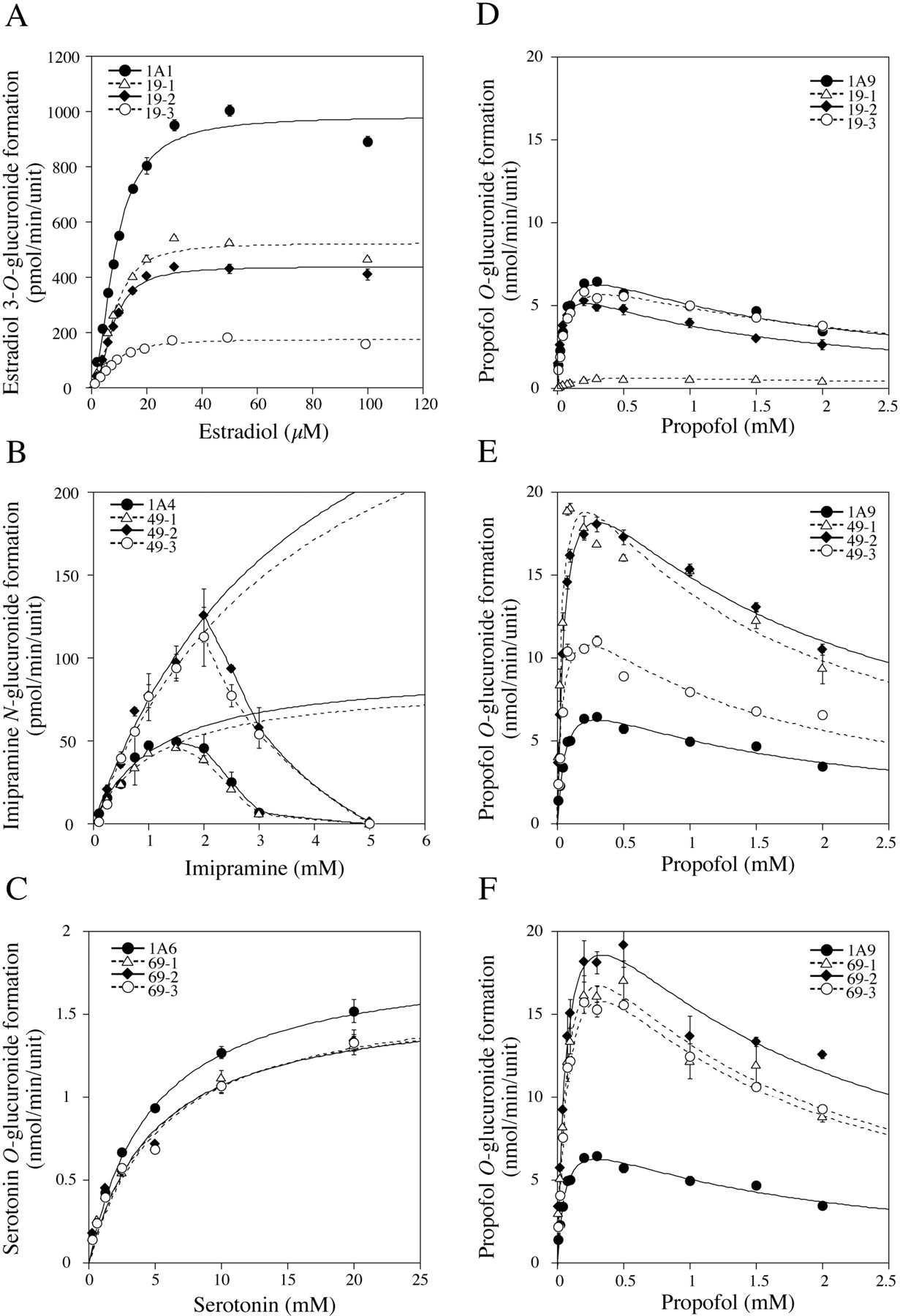
Kinetic Analyses of Estradiol 3-O-Glucuronide, ImipramineN-Glucuronide, SerotoninO-Glucuronide, and PropofolO-Glucuronide Formations. It was confirmed that the estradiol 3-O-glucuronide, imipramine N-glucuronide, serotonin O-glucuronide, and propofol O-glucuronide formations were specifically detected by UGT1A1, UGT1A4, UGT1A6, and UGT1A9, respectively (data not shown). In addition, the reproducibility of the kinetics of each glucuronide formation by the single expression systems was confirmed using multiple preparations. The estradiol 3-O-glucuronide formation by the single expression system of UGT1A1 in HEK293 cells followed the Hill equation (Fig. 7A), yielding S50 = 8.5 ± 0.2 μM, Vmax = 983.7 ± 21.2 pmol/min/unit, Vmax/S50 = 115.7 ± 1.2 μl/min/unit, and Hill coefficient, n = 1.8 ± 0.1 (Table 3). The kinetic parameters in the double expression systems of UGT1A1/UGT1A9 are shown in Table 3. The coexpression of UGT1A9 did not affect the S50 and Hill coefficient, but significantly decreased the Vmax, resulting in decreased Vmax/S50 values. Clone 19-3 with the highest ratio of UGT1A9 to UGT1A1 showed the lowest Vmax value (about one-sixth that of the single expression system).
Kinetic parameters for estradiol 3-O-glucuronide formation by UGT1A1 in single and double expression systems
Data are means ± S.D. of three independent determinations.

Effect of heat treatment on the enzymatic activities of recombinant UGT1A1 (A), UGT1A3 (B), UGT1A4 (C), UGT1A6 (D), UGT1A7 (E), UGT1A8 (F), UGT1A9 (G), and UGT1A10 (H) in baculovirus-infected insect cells (Supersomes). The thermal stability of each isoform was determined as described in Fig. 2. After heat treatment in the presence of UDPGA, estradiol 3-O-glucuronide (A), imipramine N-glucuronide (C), serotonin O-glucuronide (D), propofol O-glucuronide (G), and 7-HFC O-glucuronide (B, E, F, and H) formations were measured at the substrate concentrations of 50 μM, 1 mM, 5 mM, 200 μM, and 50 μM, respectively. Data are the mean of two independent determinations.
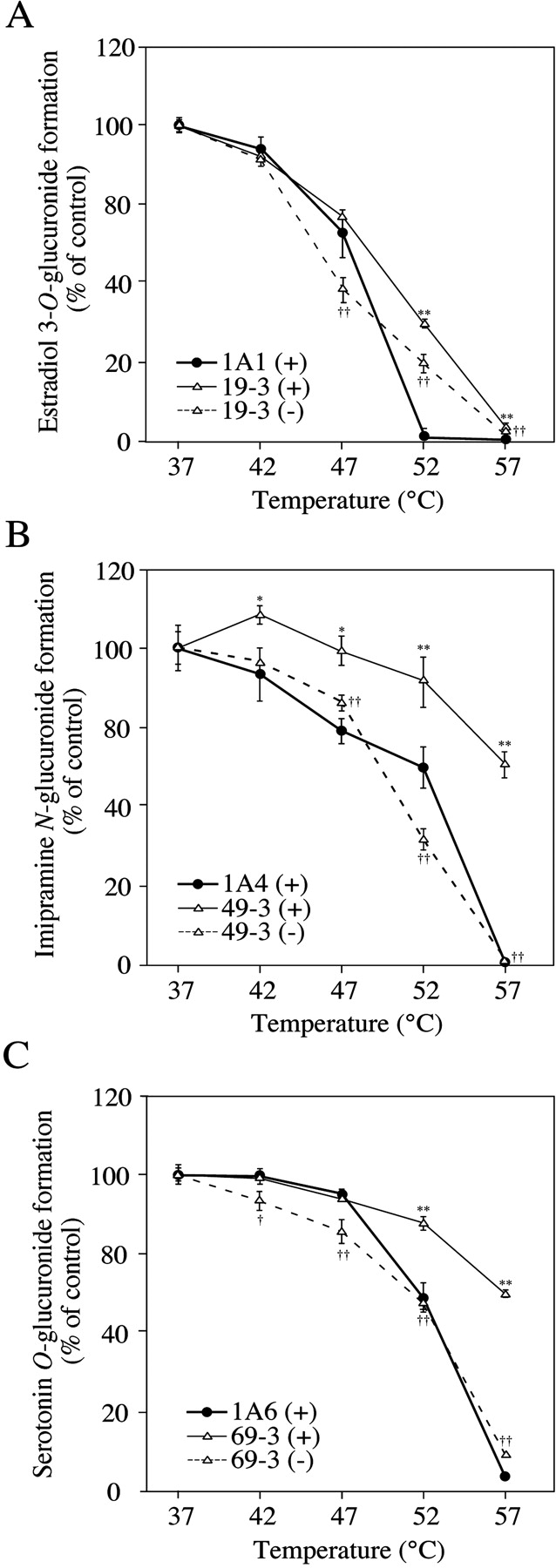
Effects of coexpression of UGT1A9 on thermal stability of UGT1A1, UGT1A4, or UGT1A6. The total cell lysate was incubated at each temperature for 15 min in the presence (+) or absence (–) of UDPGA. Estradiol 3-O-glucuronide (A), imipramine N-glucuronide (B), and serotonin O-glucuronide (C) formations were measured at the substrate concentrations of 50 μM, 1 mM, and 5 mM, respectively. Data are the means ± S.D. of three independent determinations. *, P < 0.05; ** P < 0.01, compared with the residual activities of single expression systems; †, P < 0.05; ††, P < 0.01; compared with the activities of double expression systems incubated in the presence of UDPGA.

Effects of coexpression of UGT1A9 on the inhibitory effects of Triton X-100. The residual activities are shown as percentages of the activity in the absence of Triton X-100. Estradiol 3-O-glucuronide, imipramine N-glucuronide, serotonin O-glucuronide, and propofol O-glucuronide formations were measured at the substrate concentrations of 50 μM, 1 mM, 5 mM, and 200 μM, respectively. Data are the means ± S.D. of independent three determinations. *, P < 0.05, compared with the residual activities of single expression systems.

Coimmunoprecipitation of UGT1A6 and UGT1A9. Coimmunoprecipitation was performed with cell homogenates from Mock cells, single expression systems of UGT1A6 or UGT1A9, and double expression systems of UGT1A6 and UGT1A9 (69-3) using anti-UGT1A6 antibody. The coimmunoprecipitates were subjected to SDS-polyacrylamide gel electrophoresis followed by immunoblot analysis by using anti-UGT1A antibody. As the controls, input proteins were also subjected to immunoblot analysis. IP, immunoprecipitation; WB, Western blot.
Imipramine N-glucuronide formation by the single expression system of UGT1A4 in HEK293 cells showed substrate inhibition at substrate concentrations >1.5 mM, but the plot did not fit to the substrate inhibition kinetics (Fig. 7B). When the kinetics were analyzed by fitting to Michaelis-Menten kinetics with substrate concentrations up to 1.5 mM, Km and Vmax were 1.1 ± 0.1 mM and 91.7 ± 9.5 pmol/min/unit, respectively (Table 4). Coexpression of UGT1A9 significantly increased the Km and Vmax values, resulting in slightly increased Vmax/Km. Clone 49-3 with the highest ratio of UGT1A9 to UGT1A4 showed 4-fold higher Km and Vmax than those of the single expression system (Table 4). In addition, the coexpression of UGT1A9 attenuated the substrate inhibition, shifting the substrate concentration and showing inhibition from 1.5 to 2.0 mM.
Kinetic parameters for imipramine N-glucuronide formation by UGT1A4 in single and double expression systems
Data are means ± S.D. of three independent determinations.

Kinetic analyses of various glucuronide formations by recombinant UGT1As in single or double expression systems. Estradiol 3-O-glucuronide (A), imipramine N-glucuronide (B), serotonin O-glucuronide (C), and propofol O-glucuronide (D–F) formations catalyzed by UGT1A1, UGT1A4, UGT1A6, and UGT1A9, respectively were determined as described under Materials and Methods. Data are the means ± S.D. of three independent determinations.
Serotonin O-glucuronide formation by the single expression system of UGT1A6 fitted to the Michaelis-Menten kinetics (Fig. 7C) with Km = 4.8 ± 0.2 mM and Vmax = 1.9 ± 0.1 nmol/min/unit (Table 5). Coexpression of UGT1A9 significantly increased the Km and decreased the Vmax values, resulting in decreased Vmax/Km. The Vmax/Km value of clone 69-3 with the highest ratio of UGT1A9 to UGT1A6 was 76% of that of the single expression system (Table 5).
Kinetic parameters for serotonin O-glucuronide formation by UGT1A6 in single and double expression systems
Data are means ± S.D. of three independent determinations.
Propofol O-glucuronide formation by the single expression system of UGT1A9 fitted to the substrate inhibition kinetics (Figs. 7, D–F) with Km = 59.8 ± 3.1 μM, Vmax = 8.8 ± 0.4 nmol/min/unit, and Ki = 1.5 ± 0.1 mM (Table 6). Coexpression of UGT1A1 significantly increased the Km value, decreased the Vmax value, and slightly increased the Ki value. Clone 19-1, with the highest ratio of UGT1A1 to UGT1A9, showed a 3-fold higher Km value and 9-fold lower Vmax value than those of the single expression system, resulting in 25-fold lower Vmax/Km. Coexpression of UGT1A4 significantly decreased the Km value and increased the Vmax value, resulting in increased Vmax/Km. Clone 49-1, with the highest ratio of UGT1A4 to UGT1A9, showed a 5-fold higher Vmax/Km than that of the single expression system. Coexpression of UGT1A6 significantly increased the Km and Vmax values, resulting in increased Vmax/Km. It was confirmed that mixing of each single expression system did not influence the kinetics of the enzymatic activities (data not shown). Thus, it is suggested that the coexpressed UGT isoforms functionally interacted with each other in the double expression systems.
Kinetic parameters for propofol O-glucuronide formation by UGT1A9 in single and double expression systems
Data are means ± S.D. of three independent determinations.
Discussion
UGTs play a key role in detoxification by conjugating glucuronic acid to various lipophilic compounds. It has been reported that UGTs form oligomers, but the effects of the oligomerizations on enzymatic activities have not been completely investigated. There is only one report suggesting that UGT hetero-oligomerization results in a change of the substrate specificity (Ishii et al., 2001). The authors reported that the formation ratio of morphine 3-glucuronide/morphine 6-glucuronide by guinea pig UGT2B21 was affected by the simultaneous expression of UGT2B22 in COS-7 cells. Unfortunately, a limitation of the study was that the activity was evaluated by a transient expression system. The expression levels and the expression ratios of two UGT isoforms differ in each transient transfection. Therefore, in the present study, we established stable cell lines expressing double UGT isoforms with various expression ratios to examine the effects of simultaneous expression of UGT1A isoforms on the enzymatic activities.
One approach to understanding protein-protein interactions is to investigate the thermal stability of the proteins. It has been reported that protein shows increased thermal stability when it interacts with another protein (Ruvinov and Miles, 1994). Thus, we investigated the thermal stability of the double UGT1A expression system to analyze the protein-protein interactions and found three interesting results. First, we found that UDPGA could increase the thermal stability of UGTs. This phenomenon is reminiscent of MurG (UDP-N-acetylglucosaminyltransferase from Escherichia coli) and GtfA (TDP-epi-vancosaminyltransferase from Amycolatopsis orientalis), which are in the glycosyltransferase-B superfamily as are UGTs. These enzymes have the C-terminal domains responsible for donor substrate binding and N-terminal domains responsible for aglycon binding like UGT (Hu et al., 2003; Mulichak et al., 2003). Interestingly, X-ray crystal analyses of MurG and GtfA revealed that the binding of a donor substrate causes a conformational change from the open to closed type of N- and C-terminal domains (Hu et al., 2003; Mulichak et al., 2003). Conceivably, the binding of UDPGA may trigger the conformational change to a more stable form by making a closing between the N- and C-terminal domains. Second, UGT1A9 uniquely showed thermal stability. It is interesting that UGT1A7, UGT1A8, and UGT1A10, which have 93 to 95% amino acid homology with UGT1A9, were unstable (Fig. 3). It would be worth analyzing which amino acid(s) is involved in the thermal stability in a future study. Third, UGT1A1, UGT1A4, and UGT1A6 acquired thermal stability by the coexpression of UGT1A9, suggesting that there were protein-protein interactions with UGT1A9. However, the acquisition of thermal stability was weak in the absence of UDPGA. Thus, UDPGA may play roles in the conformational stabilization of UGTs not only for monomers or homodimers but also for heterodimers.
Because UGT in microsomes is latent, various types of detergents increase the enzymatic activities by improving the permeability of substrates or UDPGA. However, high concentrations of detergents over the optimal condition conversely decrease the activity due to progressive disruption of the membrane organization of the UGT (Luukkanen et al., 1997). Kurkela et al. (2003) have reported that UGT1A9 was uniquely tolerable to the inhibitory effects of detergent. In the present study, we found that UGT1A1, UGT1A4, and UGT1A6 acquired tolerance to detergent by the coexpression of UGT1A9. The phenomenon may result from protein-protein interactions. Although the mechanism determining the tolerance to detergent remains to be clarified, particular properties of UGT1A9 per se or interaction with membrane components such as phospholipids might be relevant to the stability against heat and detergent. Finally, the immunoprecipitation assay proved an association between UGT1A6 and UGT1A9. These results strongly demonstrated the existence of protein-protein interactions between UGT1A1/UGT1A9, UGT1A4/UGT1A9, and UGT1A6/UGT1A9 in the double expression systems.
To examine the effects of the interactions on the enzymatic activities of the UGTs, single and double expression systems were used for kinetic analyses. We found that coexpression of UGT1A9 significantly affected the kinetics of the activities by UGT1A1, UGT1A4, and UGT1A6 and vice versa. The decrease in the Vmax value of estradiol 3-O-glucuronide formation by UGT1A1 was 19-1 < 19-2 < 19-3. The increase in Km and Vmax values in imipramine N-glucuronide formation was 49-1 < 49-2 < 49-3. Thus, the effects of UGT1A9 depended on the expression ratio of UGT1A9 toward UGT1A1 or UGT1A4 (Table 2). However, the effects of UGT1A9 on serotonin O-glucuronide formation by UGT1A6 were approximately equal within three clones, because the expression ratios UGT1A6/UGT1A9 were almost the same (Table 5). The expression ratio-dependent effects were also observed in propofol O-glucuronide formation by UGT1A9 (Table 6). These results suggested that the effects of coexpression of other UGT1A isoform on the kinetics were dependent on the expression ratios of each UGT1A isoform.
Extending the results obtained in the present study, comparison of Km or S50 between recombinant systems and human liver microsomes would be useful to understand the enzymatic characteristics, but the Vmax values cannot be compared, because the lack of specific antibodies prevents determination of the expression levels of each UGT1A isoform in human liver microsomes. The present study demonstrated that the Km value of propofol O-glucuronide formation by UGT1A9 was significantly increased from 59.8 to 173.1 μMbythe coexpression of UGT1A1 or UGT1A6 (Table 6). The Km values for recombinant UGT1A9 expressed in insect cell and human liver microsomes were reported to be 37 μM (Soars et al., 2003) and 190 to 213 μM (Shimizu et al., 2003; Soars et al., 2003), respectively. The higher Km values in human liver microsomes might result from protein-protein interactions between UGT1A9 and UGT1A1 or UGT1A6. Imipramine N-glucuronide formation showed substrate inhibition at concentrations >1.5 mM in the single expression system of UGT1A4 but at >2.0 mM by the coexpression of UGT1A9 (Fig. 7B). Interestingly, the kinetics in human liver microsomes also showed substrate inhibition at >2.0 to 2.5 mM (Nakajima et al., 2002; Qian and Zeng, 2006). Thus, the change of the substrate concentration showing inhibition might also result from protein-protein interactions between UGT1A4 and UGT1A9. Although the Km values for estradiol 3-O-glucuronide and serotonin O-glucuronide formations were not changed by the coexpression of UGT1A9, these values were close to that in human liver microsomes (Fisher et al., 2000; Krishnaswamy et al., 2003; Lepine et al., 2004). The Vmax values were decreased or increased by the coexpression of other isoforms. Therefore, to predict the glucuronide formations in human liver microsomes, it might be necessary to understand these UGT-UGT interactions.
In conclusion, we found that UGT1A1, UGT1A4, and UGT1A6 interact with UGT1A9, acquiring the enzymatic characteristics of the other isoform, resulting in a change of the kinetics of the enzymatic activities. Extending the lines of research presented here to the interaction between UGT1A and UGT2B may also increase our understanding of UGT activities in human liver microsomes.
Acknowledgments
We acknowledge Brent Bell for reviewing the manuscript.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.014191.
-
ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; UDPGA, UDP-glucuronic acid; HPLC, high performance liquid chromatography; 4-MU, 4-methylumbelliferone; 7-HFC, 7-hydroxy-4-trifluoromethylcoumarin; PBS, phosphate-buffered saline; HPLC, high-performance liquid chromatography.
- Received December 2, 2006.
- Accepted February 7, 2007.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}