


Abstract
Human hepatic carboxylesterase 1 and 2 (CES1 and CES2) are important for the disposition of ester- and amide-bond–containing pharmaceuticals and environmental chemicals. CES1 and CES2 ontogeny has not been well characterized, causing difficulty in addressing concerns regarding juvenile sensitivity to adverse outcomes associated with exposure to certain substrates. To characterize postnatal human hepatic CES1 and CES2 expression, microsomal and cytosolic fractions were prepared using liver samples from subjects without liver disease (N = 165, aged 1 day to 18 years). Proteins were fractionated, detected, and quantitated by Western blotting. Median microsomal CES1 was lower among samples from subjects younger than 3 weeks (n = 36) compared with the rest of the population (n = 126; 6.27 vs. 17.5 pmol/mg microsomal protein, respectively; P < 0.001; Kruskal-Wallis test). Median cytosolic CES1 expression was lowest among samples from individuals between birth and 3 weeks of age (n = 36), markedly greater among those aged 3 weeks to 6 years (n = 90), and modestly greater still among those older than 6 years (n = 36; median values = 4.7, 15.8, and 16.6 pmol/mg cytosolic protein, respectively; P values < 0.001 and 0.05, respectively; Kruskal-Wallis test). Median microsomal CES2 expression increased across the same three age groups with median values of 1.8, 2.9, and 4.2 pmol/mg microsomal protein, respectively (P < 0.001, both). For cytosolic CES2, only the youngest age group differed from the two older groups (P < 0.001; median values = 1.29, 1.93, 2.0, respectively). These data suggest that infants younger than 3 weeks of age would exhibit significantly lower CES1- and CES2-dependent metabolic clearance compared with older individuals.
Introduction
The human hepatic carboxylesterases (EC 3.1.1.1) are a family of serine esterases that have an important role in the metabolism of numerous chemicals containing ester, amide, or thioester bonds (Satoh and Hosokawa, 2006). Major pharmaceutical classes include anticoagulants, angiotensin-converting enzyme inhibitors, antihyperlipidemic agents, antiviral agents, cancer chemotherapeutics, immunosuppressing compounds, and psychoactive agents, but also several psychoactive drugs of abuse (reviewed in Laizure et al., 2013). Environmental chemicals for which the carboxylesterases are important for detoxication and disposition include the organophosphorous (Maxwell, 1992) and pyrethroid (Ross et al., 2006) insecticides. Of the genes constituting the human carboxylesterase family (Satoh and Hosokawa, 2006), carboxylesterase 1 (CES1) and carboxylesterase 2 (CES2) are the predominant forms expressed in the liver. These two enzymes share 48% amino acid sequence identity but exhibit distinct substrate (Satoh and Hosokawa, 2006) and inhibitor specificities (Parker et al., 2015). Both CES1 and CES2 are found in hepatocyte endoplasmic reticulum and cytosol (Tabata et al., 2004; Xu et al., 2002). For at least CES1, there is evidence that the cytosolic form is missing a putative 18 amino acid N-terminal signal peptide (Tabata et al., 2004). Although the mechanism controlling the dual localization of these enzymes is unknown, there is evidence that processing within the endoplasmic reticulum is necessary to retain activity (Potter et al., 1998).
In the report, Pyrethroids: Evaluation of Data from DNTs and Consideration of Comparative Sensitivity, the United States Environmental Protection Agency opined that observed age-dependent sensitivity to pyrethroids is due largely to pharmacokinetic factors, in particular the maturation of required metabolic processes (Document ID: EPA-HQ-OPP-2008-0031-0028 at http://www.regulations.gov). Because of species differences in the major pharmacokinetic factors involved in pyrethroid metabolism, however, the study of differential sensitivity between juvenile and adult rats will not inform potential human differential sensitivity. Serum esterases exhibit significant activity toward pyrethroids in rats, but not in humans (Crow et al., 2007). Human intestinal CES2 is important for the hydrolysis of some, but not all, pyrethroids, whereas rat intestinal carboxylesterases exhibit little activity (Crow et al., 2007). In the liver, both oxidative and hydrolytic pathways can be important, depending on the pyrethroid, and species differences are observed in the preferred route of metabolism in this organ. Unlike in rats, human CES1 plays a predominant role for several pyrethroids (Ross et al., 2006; Godin et al., 2006). Among pyrethroids for which oxidative metabolism is a major contributor, species differences exist in the relative importance of the cytochromes P450 involved. In rats, CYP1A1, CYP1A2, CYP2C6, CYP2C11, and CYP3A1 are the predominant metabolic contributors, whereas in humans, activity is dominated by CYP2C8, CYP2C19, and CYP3A4 (Scollon et al., 2009). Thus, for each specific pyrethroid, knowledge of relevant human enzyme developmental trajectory is necessary to inform questions regarding juvenile sensitivity, as well as rational pediatric drug dosing and avoidance of drug-drug or drug-insecticide interactions.
Considerable information is available regarding the ontogeny of human CYP2C19 (Koukouritaki et al., 2004) and CYP3A4 (Stevens et al., 2003). In contrast, limited information is available on the ontogeny of CES1 and CES2. Yang et al. (2009a) demonstrated a good correlation between human hepatic CES1 and CES2 mRNA and protein and subsequently showed that expression of both enzymes was similar to adult levels by 18 years of age based on relative mRNA levels. Expression in those 18 years of age or older was significantly higher than in individuals between birth and 10 years of age, which in turn was significantly higher than fetal expression. Consistent with this conclusion, Zhu et al. (2009) found that human hepatic CES1, but not CES2, protein levels and activity were lower in donors younger than 1 year compared with those from older samples. Both studies, however, were limited in sample size at critical ages, interpolated protein levels based on mRNA expression using a limited data set to test this correlation, and/or reported relative levels of CES1 and CES2 expression rather than age-dependent CES1- and CES2-specific content. The latter has much greater utility for extrapolating from in vitro determined intrinsic clearance values to in vivo age-dependent disposition using modeling, an approach increasingly used to predict disposition among sensitive populations for which ethical constraints prevent direct testing.
The objective of this study was to determine age-dependent changes in postnatal human hepatic CES1- and CES2-specific content in both the microsomal and cytosolic compartments, to evaluate interindividual variation in expression, and to assess differences in expression as a function of sex and/or ethnicity or race.
Materials and Methods
Human Liver Tissue Bank.
Anonymized liver tissue samples (N = 165), from donors ranging in age from birth to 18 years, were obtained from the Brain and Tissue Bank for Developmental Disorders, University of Maryland, Baltimore. Samples from individuals likely to have experienced liver disease based on the reported cause of death were excluded. A summary of donor demographics are provided in Table 1. Liver microsome and cytosolic fractions were prepared by differential centrifugation as described in Koukouritaki et al. (2002) and stored at −80°C until use. The collection and described use of these tissue samples were considered exempt by the Children’s Hospital and Health System of Wisconsin Institutional Review Board.
Tissue sample donor demographics
To ensure good age representation and adequate power, target sample sizes for birth to 30 days, greater than 30 days to 1 year, greater than 1–5 years, greater than 5–10 years, and greater than 10–18 years age brackets were developed based on data from the existing literature on drug and toxicant metabolizing enzyme ontogeny. Samples sizes were sufficient to provide at least 80% power to detect a 1 S.D. change in enzyme specific content between age brackets assuming α = 0.05.
Other Materials.
Primary polyclonal antibody raised in rabbits against the carboxy-terminus end of purified human CES1 was obtained from OriGene (Rockville, MD). Primary polyclonal antibody raised in rabbits against a conjugated human CES2 peptide sequence corresponding to a region between amino acids 51 and 469 was obtained from Novus Biologicals (Littleton, CO). Purified recombinant CES1 protein fused to a polyhistidine carboxy-terminal tag was obtained from Novoprotein (Summit, NJ). Purified recombinant CES2 protein with a carboxy-terminal MYC/DDK tag was obtained from OriGene (catalog no. TP303009). Specificity of the primary antibodies was verified by evaluating cross-reactivity of the CES1 and CES2 primary antibodies against the highest concentration of recombinant CES2 and CES1 protein used in the analysis, respectively. Horseradish-conjugated donkey anti-rabbit IgG was obtained from GE Healthcare Life Sciences (Piscataway, NJ). Prestained protein molecular weight markers were obtained from Invitrogen (Grand Island, NY).
Western Blot Analysis.
Aliquots of individual microsomal and cytosolic tissue preparations were fractionated by SDS-PAGE, along with a range of CES1- and CES2-purified, recombinant protein concentrations and molecular weight standards essentially as described earlier (Koukouritaki et al., 2002). Fractionated proteins subsequently were transferred to a nitrocellulose-based membrane (Hybond, GE Healthcare Life Sciences) by electrophoresis. The membrane was incubated with either the CES1 (1:10,000 dilution) or CES2 (1:2,500 dilution) polyclonal antibody, followed by the horseradish peroxidase-conjugated secondary antibody (1:10,000 dilution). Visualization was accomplished using enhanced chemiluminescence (GE Healthcare Life Sciences ECL Plus Detection System) and the ChemiDoc CCD Imaging Analysis system (Bio-Rad, Hercules CA). Linear regression analysis was used to quantify the amount of both CES1 and CES2 protein in each tissue fraction based on the signals observed with the respective purified, recombinant proteins. An r2 ≥ 0.95 was accepted as evidence of assay linearity. The interday coefficients of variation for microsomal and cytosolic CES1 measurements were 7.3% and 6.1%, respectively (n = 5 replicates each). The interday coefficients for variation for microsomal and cytosolic CES2 measurements were 3.2% and 4.1% (n = 5 replicates each).
Statistical Analysis.
Scatter plots of protein-specific content as a function of age were used to evaluate overall trends. Regression-tree analysis using CART version 6 (Salford Systems, San Diego, CA) was used to evaluate possible age threshold effects. Least average deviation from the median was used to define the tree nodes. Statistical comparisons between age groups were performed using Kruskal-Wallis nonparametric tests, followed by stepwise step-down comparisons of all pairs of data sets (IBM SPSS Statistics 19; SPSS Inc., Chicago, IL). Other variables of interest were compared using nonparametric testing. Linear regression was used to test relationships between continuous variables with the strength of the r2 (i.e., proximity to 1) and the P value being considered highly relevant. Analysis of variance (ANOVA) and stepwise regression testing were used to assess the multiple factors potentially associated with enzyme content.
Results
Detection of Microsomal and Cytosolic CES Proteins.
The developmental expression pattern of the human hepatic CES enzymes was determined by SDS-PAGE and Western blot analysis of microsomal and cytosolic samples prepared from a bank of 165 human pediatric liver samples donated by individuals over a wide postnatal age range at the time of death. For CES1, Western blotting revealed a single, major immunoreactive band in both the microsomal and cytosolic fractions with an apparent molecular mass near 61 kDa, corresponding to the intact CES1 protein (Fig. 1, A and B); on reblotting, an apparent molecular mass near 69 kDa, corresponding to the intact CES2 protein (Fig. 1, C and D). No cross-reactivity between the anti-CES1 antibody and CES2 protein (Fig. 1A, lanes 2–6) or between the anti-CES2 antibody and CES1 protein was observed (Fig. 1D, lanes 13–17).

Western blot analysis of human CES1 and CES2 in pediatric hepatic microsomal samples. (A) Western blot with anti-CES1 antibody; lane 1, molecular weight standards as shown (kDa); lanes 2–6, purified recombinant CES2 standards (6.5, 12, 25, 50, and 100 ng); lanes 7–18, microsomal hepatic protein samples (10 µg each). (B) Western blot with anti-CES1 antibody; lanes 1–12, microsomal hepatic protein samples (10 µg each); lanes 13–17, purified recombinant CES1 standards (12.5, 25, 50, 100, 150 ng); lane 18, molecular weight standards as shown (kDa). (C) Western blot with anti-CES2 antibody; lane 1, molecular weight standards as shown (kDa); lanes 2–6, purified recombinant CES2 standards (6.5, 12, 25, 50, and 100 ng); lanes 7–18, microsomal hepatic protein samples (10 µg each). (D) Western blot with anti-CES2 antibody; lanes 1–12, microsomal hepatic protein samples (10 µg each); lanes 13–17, purified recombinant CES1 standards; lane 18, molecular weight standards as shown. Microsomal samples from differing age groups were loaded in a nonordered fashion.
Overall CES Variation and Demographic Variables.
Both CES1 and CES2 were detectable in most of the samples. With outliers included, microsomal and cytosolic CES1 varied 30- and 70-fold, whereas CES2 varied 11- and 8-fold, respectively. Without considering extreme values, however, the overall distribution of CES content for the entire sample varied about 10- and 6-fold for CES1 and CES2, respectively. No relationship was found between the postmortem interval and the specific content of either CES1 or CES2 (linear regression, each r2 ≤ 0.02; each P > 0.05). CES1 and CES2 protein levels in both microsomes and cytosol were similar in male and female samples (data not shown).
When assessed independent of age or any other factors, race or ethnicity was associated with microsomal CES1 and CES2 protein expression (Table 2). Specifically, samples from white donors had greater microsomal CES1 expression than those from African Americans, which had greater expression than those from Hispanics (P ≤ 0.05 and ≤ 0.01, respectively, Mann Whitney U testing). Similarly, microsomal CES2 expression was significantly greater in both African Americans and whites than in Hispanics (P ≤ 0.001; each comparison; Mann-Whitney U testing). Cytosolic CES1 was modestly greater in whites compared with African Americans (P = 0.05), whereas no expression differences in cytosolic CES2 were observed.
Immunodetected carboxylesterase (CES) content from postmortem fractionated hepatic samples
Data are given in median pmol/mg protein (interquartile range). Each enzyme is compared across the three donor race/ethnicity groups.
Human CES1 Ontogeny.
CES1 was readily detected in early life hepatic samples, with expression occurring as early as the first day of life in some, but not all, microsomal and cytosolic samples. Complex relationships were observed between age and both microsomal and cytosolic CES1, with a substantial degree of variability across the entire age range (Fig. 2).

The relationship between human CES1 and age in postmortem microsomal and cytosolic liver samples. (A) Overall relationship with microsomal CES1 in samples from donors aged birth to 18 years (n = 161). The two added vertical lines represent the two nodes, aged 3 weeks (left line) and 6 years (right line), selected by classification regression tree analysis as indicative of appropriate age groupings. The 3-week age classification (solid line) was confirmed on Kruskal-Wallis statistical testing, whereas the 6-year grouping (indicated by a dotted line) was not. (B) Relationship between microsomal CES1 and age in liver samples from the subset of subjects younger than 1 year (n = 102). The added solid vertical line represents the 3-week time point selected by classification trees and confirmed by statistical testing as appropriate age stratification. (C) Relationship between cytosolic CES1 and age in postmortem cytosolic liver samples from birth to 18 years (n = 162). The vertical lines represent the two nodes selected by classification trees as indicative of appropriate age groupings: 3 weeks (left line) and 6 years (right line). Both were confirmed as statistically significant using Kruskal-Wallis testing. (D) Relationship between cytosolic CES1 and age in postmortem cytosolic liver samples from a subset of subjects younger than 1 year (n = 101). The vertical line represents the time point selected by classification trees as an appropriate age grouping (3 weeks).
Based on both classification tree analysis and confirmatory Kruskal-Wallis testing, microsomal CES1 activity was lower among samples from subjects younger than 3 weeks of age compared with the rest of the population. This age differential appeared to be driven by the onset of expression during the first 3 weeks of life, as no subject older than 18 days had nondetectable microsomal CES1 expression (Fig. 2, A and B). A second node at 6 years was identified by the initial tree analysis, but it was not statistically significant (P = 0.13) when considered in conjunction with the 3-week node. Thus, samples from individuals between birth and 3 weeks of age [N = 36, median value; interquartile range, IQR = 6.27 (4.2–13.4) pmol/mg microsomal protein] were lower than those from ages 3 weeks to 6 years [n = 91, median value (IQR) = 16.8 (13.2–20.8) pmol/mg microsomal protein; P < 0.001; Kruskal-Wallis test, Fig. 3], but the CES1-specific content in the later age group was not statistically different from that in the greater than 6 years of age group [n = 34, median value (IQR) = 18.3 (16.7–21.1) pmol/mg microsomal protein; P = 0.13, Kruskal-Wallis test]. For cytosolic CES1, expression was much lower among liver samples from individuals between birth and 3 weeks of age [n = 36, median value (IQR) = 4.7 (0–8.4) pmol/mg cytosolic protein, Fig. 3] compared with those from donors aged 3 weeks to 6 years [n = 90, median value (IQR)= 15.8 (11.5–20.5) pmol/mg cytosolic protein; P < 0.001; Kruskal-Wallis test], which were, in turn, modestly lower compared with those older than 6 years [n = 36, median value (IQR) = 16.6 (14.3–25.3) pmol/mg cytosolic protein; P = 0.05; Kruskal-Wallis test].

Summary of microsomal (white boxes) and cytosolic (gray boxes) human CES1 developmental expression pattern. CES1-specific content as a function of age was grouped using classification tree analysis to minimize differences within while maximizing differences between age brackets. The resulting data are shown as box and whisker plots in which the horizontal bar represents median CES1 content, boxes the upper and lower quartiles, and vertical bars the 5th to 95th percentiles. Outliers, defined as having specific contents outside 1.5 times the 25th to 75th percentiles, are shown as open circles but were excluded from the analyses except for determining absolute ranges in expression. The youngest age group differed significantly from the other two for both matrices (P < 0.001, each comparison; Kruskal-Wallis testing), whereas the middle age group was modestly significantly different from the older group in CES1 cytosolic content (P = 0.05) but did not differ in CES1 microsomal content (P = 0.13).
Considering the population as a whole, as CES1 microsomal content increased, there was a corresponding increase in the cytosolic CES1 content (see trend in Fig. 3); however, when microsomal and cytosolic CES1 content were compared within individuals, only a modest relationship was observed (r2 = 0.207, P < 0.001, linear regression; data not shown). Both microsomal and cytosolic CES1 values were measured in 160 of 165 samples; of these, the enzyme was detectable in one or the other compartment in 154 samples. Microsomal CES1 content was greater than cytosolic CES1 content in 87 of 154 samples, with a median difference of 5.8 pmol/mg protein (range = 0.1–59.5 pmol/mg protein). Cytosolic CES1 content was greater than microsomal CES1 content in 67 of 154 samples, with a median difference of 5.5 pmol/mg protein (range = 0.1–63.3 pmol/mg protein). Taken together, these data argue against microsomal content determining cytosolic content and also suggest that CES1 content in the two matrices is independently regulated.
Human CES2 Ontogeny.
Similar to human CES1, CES2 also increased with age with variation apparent across the age-range spectrum. The most marked changes occurred during the first year of life (Figs. 4). Although CES2 was tested using classification tree analysis separately from CES1, the same age nodes were selected for CES2.
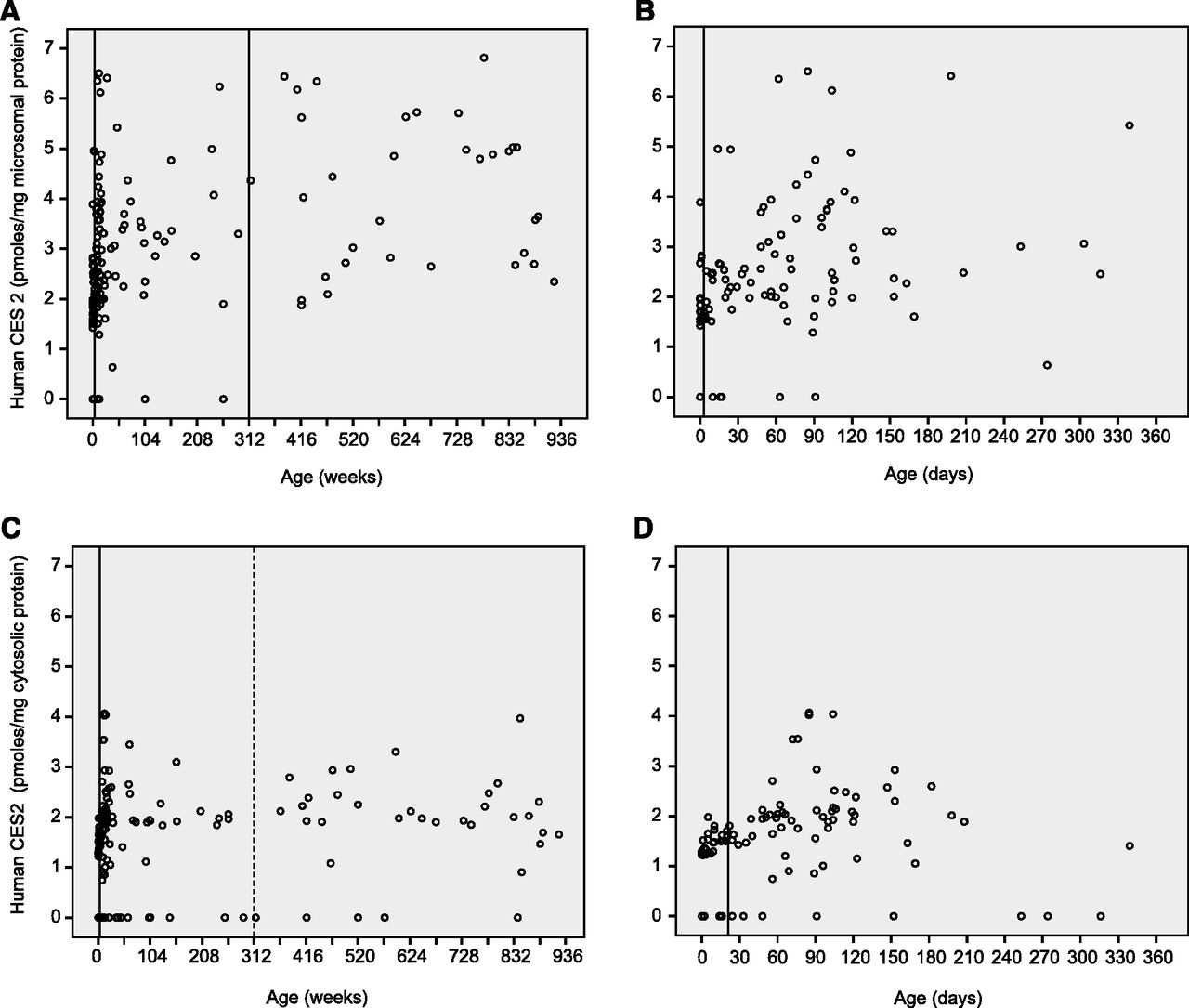
The relationship between human CES2 and age in postmortem microsomal and cytosolic liver samples. (A) The relationship between microsomal CES2 and age in postmortem liver samples from donors aged birth to 18 years (n = 161). The vertical lines represent the two nodes selected by classification trees as indicative of appropriate age groupings: 3 weeks (left line) and 312 weeks or 6 years (right line). Both were confirmed as statistically significant using Kruskal-Wallis testing. (B) The relationship between microsomal CES2 and age in postmortem liver samples from the subset of subjects younger than 1 year (n = 102). The vertical line represents the time point selected by classification trees as an appropriate age grouping (3 weeks). (C) Relationship between cytosolic CES2 and age from postmortem liver samples (n = 162). The two vertical lines at 3 weeks (left) and 6 years (right) of age represent the nodes selected by classification trees. The 3-week age classification (solid line) was confirmed on Kruskal-Wallis statistical testing, whereas the 6-year grouping (indicated by a dotted line) was not. (D) Relationship between cytosolic CES2 and age in postmortem liver samples from a subset of subjects younger than 1 year (n = 101). The vertical line at 3 weeks represents the time point selected by classification trees and statistically confirmed (Kruskal-Wallis) as an appropriate age grouping.
Based on both classification tree analysis and confirmatory Kruskal-Wallis testing, microsomal CES2 was lower in infants from birth to 3 weeks of age [n = 36; median value (IQR) = 1.8 (1.6–2.5) pmol/mg microsomal protein] compared with those between 3 weeks and 6 years of age [n = 91; median value (IQR) = 2.9 (2.1–3.7) pmol/mg microsomal protein; P < 0.001, Kruskal-Wallis test, Fig. 5]. The middle age group, those older than 3 weeks but younger than or equal to 6 years of age, had values that were, in turn, lower than values in the oldest group, children over age 6 years [n = 34; median value (IQR) = 4.2 (2.7–5.2) pmol/mg microsomal protein P < 0.001, Kruskal-Wallis test]. Cytosolic CES2 content was lower in samples from infants from birth to 3 weeks of age [n = 36; median (IQR) = 1.29 (0–1.53) pmol/mg microsomal protein, Fig. 5] compared with the two older age groups [n = 91 and n = 34, 1.93 (1.3–2.1) and 2.0 (1.4–2.5), respectively (P < 0.001, each comparison; Kruskal-Wallis test)]; however, the two older age groups did not differ in cytosolic CES2 content (P = 0.26).

Summary of microsomal (white boxes) and cytosolic (gray boxes) human CES2 developmental expression pattern. CES2-specific content as a function of age was grouped using classification regression trees to minimize differences within age groups while maximizing differences between age brackets. The resulting data are shown as box and whisker plots in which the horizontal bar represents median CES2 content, boxes are the upper and lower quartiles, and vertical bars the 5th to 95th percentiles. Outliers, defined as having specific contents outside 1.5 times the 25th to 75th percentiles, are shown as open circles but were excluded from the analyses except for reporting absolute expression ranges. The youngest age group differed significantly from the other two for both matrices (***P < 0.001, each comparison, Kruskal-Wallis testing), whereas the middle age group was significantly different from the older group in CES1 microsomal content (P ≤ 0.001) but did not differ in CES1 cytosolic content (P = 0.26).
Similar to human CES1, when comparing the amount of microsomal and cytosolic CES2 in individual samples, there was a statistically significant, but numerically modest relationship (r2= 0.08, P < 0.001, linear regression) (data not shown). Microsomal and cytosolic CES2 was measured in 157 of 165 samples and, of these, was detectable in one or the other compartment in 154 samples. Microsomal CES2 content was greater than cytosolic content in 133 of 154 samples with a median difference of 1.42 pmol/mg protein (range = 0.03–6.24 pmol/mg protein). Cytosolic CES2 content was greater than microsomal content in only 21 of 154 samples, with a median difference of 0.84 pmol/mg protein (range = 0.03–11.60 pmol/mg protein).
Simultaneous Consideration of Factors Impacting CES Expression.
Considering age category, sex, and race or ethnicity simultaneously, only the variable “age category” remained significantly associated with human CES1 content in both the microsomal and cytosolic compartments (P < 0.001, each model; ANOVA). Cytosolic CES2 was similar; that is, only the age categorization was significantly associated (P < 0.001, ANOVA). For microsomal CES2, the relationship between enzyme content and sex was ambiguous (P = 0.08, ANOVA) when age was considered simultaneously. Nevertheless, the influence of age on CES2 enzyme content continued to be highly significant, with sex considered simultaneously (P < 0.001, ANOVA). Of note, with age group considered simultaneously, race or ethnic group was no longer significantly associated with CES1 or CES2 in either compartment.
Discussion
The results of this project confirm that human hepatic microsomal and cytosolic CES1 and CES2 expression are developmentally regulated based on the strong association between postnatal age and quantitatively measured protein values. For both enzymes, expression was markedly lower among infants 3 weeks of age or younger compared with older infants and children. Of note, for microsomal CES1, all samples exhibited some expression after 3 weeks, whereas the universal onset of CES2 expression appeared somewhat later, or its expression was suppressed by an unidentified mechanism in a small number of samples. After 3 weeks of age, ongoing developmental changes varied by enzyme and by compartment. Microsomal CES1 and cytosolic CES2 did not exhibit additional age-related differences. In contrast, for microsomal CES2 and cytosolic CES1, the samples from children over the age of 6 years exhibited statistically significantly greater expression than those between 3 weeks and 6 years of age, which suggests that significant developmental changes continue during the window between 3 weeks and 6 years. This finding may be particularly relevant for CES1 given its high expression levels in the liver; however, the rate of change during this time appears modest as reflected in the relatively small numerical change over a wide time interval compared with the relatively steep surge in expression that occurs during the first 3 weeks after birth. Race or ethnicity appeared to be associated with differences in enzyme expression in univariate testing (Table 2), but when this was evaluated with multivariate testing that included age, it was no longer significant; age was the only factor that entered into the model.
These conclusions are consistent with literature reports that have evaluated changes in CES1 and CES2 mRNA levels during early life stages (Yang et al., 2009a), protein expression, and activity data (Yang et al., 2009a; Zhu et al., 2009); however, the limited data sets used in these earlier studies prevented a precise determination of the developmental trajectory, and neither study differentiated between carboxylesterase content in the microsomal and cytosolic compartments. Yang et al. (2009a) demonstrated significant differences in mRNA content and activity only between fetal (12–32 weeks), children (birth to 10 years), and adults (≥18 years), whereas Zhu et al. (2009) were able to demonstrate combined hepatic microsomal and cytosolic CES1, but not CES2, was significantly lower in tissue samples from individuals younger than 1 year compared with older age groups. This finding contrasts to the data presented herein, which demonstrates a significant difference between infants younger than 3 weeks and older children. These data, combined with those from Yang et al. (2009a) and Zhu et al. (2009), convincingly document that both human hepatic microsomal and cytosolic CES1 and CES2 belong to the class 3 group of enzymes involved in drug and toxicant disposition; that is, they exhibit low to no expression in the fetus and that expression increases substantially during the first months to 2 years of life (Hines 2012).
Over the entire population, microsomal and cytosolic CES1 varied about 30- and 70-fold, respectively, whereas microsomal and cytosolic CES2 varied about 11- and 8-fold, respectively. In both compartments, the median CES1 value varied by about 3- to 3.5-fold across the three age groups, whereas the median CES2 value varied by 1.5- to 2.5-fold. Importantly, the within age brackets, variation was substantive. We speculate that this variability within age brackets is likely explained in part by genetic variations. Six CES1 single-nucleotide polymorphisms have been identified that in vitro exhibit various degrees of decreased activity or increased promoter activity or in vivo are associated with altered drug disposition. Three CES2 single-nucleotide polymorphisms have been associated with decreased activity both in vitro and in vivo (reviewed in Merali et al., 2014). Whether any of these genetic variants function through altered protein levels versus changes in enzyme activity is unknown, however. The CES1 rs3785161 variant that is associated with increased promoter activity (Geshi et al., 2005) and the CES1 rs2241409 variant that is associated with decreased transcript levels (Marsh et al., 2004) would be expected to alter enzyme expression levels, but this has not been shown directly. Exposures to environmental factors may also have contributed to the observed variability. Inflammatory cytokines, such as interleukin 6, have been shown to repress CES1 and CES2 expression through a distal regulatory element (Yang et al., 2007) and a nuclear factor-erythroid 2–related factor element at CES1 position −2025 mediates increased expression in response to oxidative stress (Maruichi et al., 2010). Finally, Ghosh and Natarajan (2001) identified three functional peroxisomal proliferator response elements at CES1 positions −176, −779, and −1316 that mediate transcriptional repression by ligand-activated peroxisomal proliferator activated receptor (PPAR) α or PPARγ. Although the pregnane X receptor is involved in the induction of many genes encoding xenobiotic metabolizing enzymes, this receptor is not involved in regulating CES1 or CES2 expression (Yang et al., 2009b).
This study is limited by the absence of CES1 and CES2 activity data for each sample. Such data are desirable if highly specific substrates are available and assays can be developed with sufficient sensitivity to allow an adequate signal to noise ratio with the small amount of protein available for each sample in the tissue bank; this was not the case for these enzymes. Past experiments performed to evaluate the developmental trajectories of other enzymes using this same tissue bank and where highly sensitive and specific activity assays were available demonstrated excellent correlation between enzyme specific content and specific activity (Koukouritaki et al., 2004; Li et al., 2012). This past experience provides some confidence that a similar relationship would be observed for CES1 and CES2. A second limitation is that the sample set did not include tissue from adults (>18 years of age) as a comparator. Indeed, because the tissue bank was designed to have maximum power to detect differences at younger ages, the median age of the entire sample set was 3.5 months and of the samples from donors younger than 3 weeks, 9.6 days. Thus, one might expect the reported median CES1 and CES2 protein levels in the age bracket of donors older than 3 weeks to be less than one would observe in adults (i.e., older than 18 years). Consistent with this expectation, Shi et al. (2011) reported an approximate 2-fold increase in mean relative CES1 protein levels in hepatic S9 fractions from donor samples between 5 weeks and 6.5 months of age and adults (>18 years of age). Similar results have recently been reported for CES2 (Chen et al., 2015).
The data from this project will be used in conjunction with available data on the ontogeny of other relevant enzymes and numerous physiologic variables to develop high-quality physiologically based pharmacokinetic models for specific CES1 and CES2 substrates. Importantly, because of the somewhat unique dual compartment localization of the hepatic CES enzymes, such models should consider the contribution of both compartments to clearance. The anticipated simulations will be valuable for therapeutics but will be critical for assessing the safety of compounds such as the pyrethroids and pyrethrins, which cannot be directly studied in children. As such, these new data represent a significant advance over existing data, which were insufficient for this purpose both because of the poor time resolution and use of relative units of measure.
Acknowledgments
The authors thank Jie He for technical assistance in generating the data for this manuscript.
Authorship Contributions
Participated in research design: Hines, Simpson, McCarver.
Performed data analysis: Hines, Simpson, McCarver.
Wrote or contributed to the writing of the manuscript: Hines, Simpson, McCarver.
Footnotes
- Received December 16, 2015.
- Accepted January 28, 2015.
↵1 Current affiliation: U.S. Environmental Protection Agency, Office of Research and Development, National Health and Environmental Effects Research Laboratory, Research Triangle Park, North Carolina.
↵2 Cary, North Carolina.
This study was funded by the Council for the Advancement of Pyrethroid Human Risk Assessment.
Abbreviations
- ANOVA
- analysis of variance
- CES1
- carboxylesterase 1
- CES2
- carboxylesterase 2
- IQR
- interquartile range
- PPAR
- peroxisomal proliferator activated receptor
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}