


Abstract
Although the liver has been considered the most important organ involved in glucuronidation, recent studies have focused on the role of the gastrointestinal tract in the glucuronidation of xenobiotics and endobiotics. Two UDP-glucuronosyltransferase (UGT) isoforms of human intestinal mucosa, which are absent in liver, have been identified by reverse transcriptase-polymerase chain reaction. mRNAs of UGT1A8 and UGT1A10 were detected in both the small intestine and the colon. The corresponding cDNAs for UGT1A8 and UGT1A10 were cloned from ileal RNA and inserted into the mammalian expression vector pcDNA3. Transfection of the cDNAs into human embryonic kidney 293 cells was carried out and stable expression was achieved. Membrane preparations from human embryonic kidney 293 cells expressing either UGT1A8 or UGT1A10 were isolated and the expression of each isoform was analyzed by Western blot. The catalytic activity of stably expressed UGT1A8 toward catechol estrogens, coumarins, flavonoids, anthraquinones, and phenolic compounds was much higher than that of UGT1A10. UGT1A8, but not UGT1A10, catalyzed the glucuronidation of opioids, bile acids, fatty acids, retinoids, and clinically useful drugs, such as ciprofibrate, furosemide, and diflunisal. These studies suggest that human intestinal UGTs may play an important role in the detoxification of xenobiotic compounds and, in some cases, limit the bioavailability of therapeutic agents.
Glucuronidation is an important metabolic process in which many xenobiotics and endobiotics are converted to more hydrophilic compounds, which are more readily eliminated from the body (Dutton, 1980). Glucuronidation reactions are catalyzed by UDP-glucuronosyltransferases (UGTs)1 that are located on the luminal side of the endoplasmic reticulum and in the nuclear envelope of hepatocytes and cells in other organs. Currently, over 40 different UGT isoforms from various species have been identified through molecular cloning techniques. Based on evolutionary divergence, at least two UGT gene subfamilies have been described:UGT1 and UGT2 (Mackenzie et al., 1997). Gene products of the UGT2 subfamily are transcribed from unique genes (Mackenzie and Rodbourn, 1990; Haque et al., 1991). In contrast, members of the UGT1 gene complex are encoded by unique first exons (2nd, 11th, and 12th are pseudogenes with premature stop codon) and four common exons leading to protein products with unique N-terminal regions and identical C-terminal domains (Ritter et al., 1992).
Liver has been widely considered as the major drug-metabolizing organ. However, recent studies (Watkins, 1997) have shown that gastrointestinal metabolism may also play an important role in the elimination of orally administered drugs, potentially toxic compounds ingested with food, or chemoprotective substances such as antioxidants present in the diet. To obtain a better understanding of the role of the gastrointestinal tract in the metabolism of drugs, natural products, and toxins, our studies have focused on UGTs found only in the gastrointestinal tract.
UGT1A8 and UGT1A10 are 94% similar in primary amino acid sequence. These genes have recently been cloned from human intestinal mucosa and expressed by three different laboratories. Strassburg et al. (1998)reported identification of UGT1A8 and UGT1A10 from human colonic mucosa. Although UGT1A10 as transiently expressed in COS-1 cells was capable of glucuronidating certain phenolic compounds, anthraquinones, coumarins, and steroids, UGT1A8 was reported to be inactive toward the substrates tested. UGT1A8 and UGT1A10 were also isolated by Mojarrabi and Mackenzie (1998) and, after transfection in COS-7 cells, both expressed enzymes catalyzed the glucuronidation of benzo(α)pyrene and 2-acetylaminofluorene metabolites. We have recently reported (Cheng et al., 1998a) the cloning of human UGT1A8. On transfection into human embryonic kidney 293 (HK293) cells, human UGT1A8 catalyzed the glucuronidation of coumarins, anthraquinones, flavonoids, phenolic compounds, and certain steroids. In this report we compare the catalytic activity of human UGT1A8 with UGT1A10 toward an extended range of chemical compounds using membrane preparations from HK293 cells. The reactivity and relative enzymatic efficiency of these isoforms were investigated and are reported here.
Experimental Procedures
Materials.
Human gastrointestinal tissue was obtained from organ donors from the University Hospital (Little Rock, AR), according to a protocol approved by the Human Research Advisory Committee of the University of Arkansas for Medical Sciences (A.R.-P.). UDP-[U-14C]GlcUA (311 mCi/mmol) was purchased from ICN Pharmaceuticals (Irvine, CA). Aglycone substrates, saccharolactone, UDP-GlcUA, dithiothreitol, and geneticin (G418) were obtained from Sigma Chemical Co. (St. Louis, MO). [11,12-3H] all-trans RA was purchased from DuPont-NEN (Boston, MA). 5,6-Epoxy-RA and 4-hydroxy-RA were synthesized as described previously (Radominska et al., 1997). Protein assay reagents were obtained from Bio-Rad (Hercules, CA). All other reagents were of analytical grade.
Identification of the UGT1A10 Transcript from Human Intestinal Tract.
The reverse transcriptase-polymerase chain reaction (RT-PCR) was used to identify the transcript of UGT1A10. RNA isolation and cDNA samples were prepared according to methods described by Cheng et al. (1998a). An exon 1-specific sense primer of UGT1A10(5′-CCTCTTTCCTATGTCCCCAATGA-3′) was generated and used in conjugation with an antisense primer of UGT1A10(5′-GTTTTCAAGCTTGGAAATGACTAGGG-3′) located within the 3′ noncoding region of exon 5 for PCR amplification. Thirty-five cycles consisting of reactions at 95°C for 1 min, 54°C for 1 min, and 72°C for 2 min were used to amplify double stranded DNA and human glyceraldehyde-3-phosphate dehydrogenase primers (Münzel et al., 1996) were added to each PCR reaction as a positive control. PCR products were verified by DNA sequencing at the DNA core facility of the University of Iowa.
Isolation and Stable Expression of Full-Length Human UGT1A8 and UGT1A10.
The isolation of full-length UGT1A8 cDNA was described previously byCheng et al. (1998a). Based on the UGT1 gene sequence (access number U39550), primers incorporating the initiation codon ofUGT1A10 (5′-TTTCTAAGCTTCTCGGGCTGCAGTTCTCTCATG-3′) and the stop codon (5′-CTCATGGATCCCACTTCTCAATGGGTCTTGG-3′) of the UGT1Agene complex were synthesized. These primers, which containHindIII or BamHI restriction site, were used to amplify full-length UGT1A10 cDNA from total human ileal cDNA. Thirty-five cycles consisting of reactions at 95°C for 1 min, 62°C for 1 min, and 72°C for 2.5 min were performed to amplify double stranded DNA. The partially HindIII-BamHI digested PCR product was subcloned into theHindIII/BamHI restriction sites of the mammalian expression vector pcDNA3. The UGT1A8 and UGT1A10 expression plasmids were transfected into HK293 cells using the calcium-phosphate method (Chen et al., 1998). Membrane preparations from HK293 cells stably expressing human UGT1A8 and UGT1A10 were prepared according to methods described previously (Chen and Okayama, 1988).
Western Blot.
The total protein in membrane preparations from HK293 cells stably expressing UGT1A8 and UGT1A10 were separated on a 10% SDS-polyacrylamide gel. The separated proteins were electrotransferred to nitrocellulose membranes and probed with a rabbit anti-UGT1A antiserum provided by Drs. Chris Patten and Charles Crespi, Gentest Corporation (Woburn, MA). The proteins were visualized with a goat anti-rabbit horseradish peroxidase-conjugated secondary antibody and SuperSignal substrate detection system (Pierce, Rockford, IL).
Glucuronidation Assays.
Membrane preparations from HK293 cells stably expressing UGT1A8 and UGT1A10 were suspended in 10 mM Tris-buffered saline (pH 7.4) containing 0.5 mM dithiothreitol. Glucuronidation activity toward steroids was assayed using the extraction method described by Matern et al. (1994). Retinoid glucuronidation was measured using the methods described by Little et al. (1997). Opioid glucuronidation assays were analyzed using the methods of Puig and Tephly (1986). All other substrates were analyzed using the thin layer chromatography method ofBansal and Gessner (1980). All assay mixtures (100 μl total volume) contained 50 mM Tris-HCl, 10 mM MgCl2, 8.5 mM saccharolactone, 2 mM UDP-GlcUA, 200 nM UDP-[U-14C]GlcUA (0.1 μCi) and varying concentrations of aglycones. All of the enzyme assays were carried out at pH 7.4 with 0.05 mg of protein at 37°C for 1 h. Glucuronidation activities for the substrates used in the present studies were not detected in nontransfected or pcDNA3-transfected HK293 cell membrane preparations. Glucuronidation rates were calculated assuming that all substrates formed monoglucuronides.
The apparent Km values for the aglycones were determined at the optimal pH, protein concentration, and reaction times, yielding linear product formation. The apparentKm was determined by varying the concentration of aglycones from approximately one-tenth to ten times the estimated Km using 2 mM UDP-GlcUA. Kinetic parameters were calculated using the Enzyme Kinetics program (Trinity Software, Plymouth, NH).
Results
Tissue Distribution and Cloning of Human UGT1A10.
RT-PCR was performed to determine the tissue distribution of UGT1A10 transcripts, and the cDNAs of the correct molecular size, 1089 bp, were observed from human jejunum, ileum, and colon (Fig.1). DNA sequencing analysis confirmed the transcripts found were those of UGT1A10. A cDNA with a complete UGT1A10 coding region was synthesized from total human ileal RNA by RT-PCR and cloned into the mammalian expression vector, pcDNA3. Sequencing revealed that the insert sequence was identical with that published previously (Mojarrabi and Mackenzie, 1997).

Intestinal expression of human UGT1A10.
Using RT-PCR, a 1089-bp product corresponding to human UGT1A10 was observed in adult jejunum (lane 3), ileum (lane 4), and colon (lane 5), but was not expressed in liver (lane 6) and kidney (lane 7). A 299-bp product was also present, which corresponds to glyceraldehyde-3-phosphate dehydrogenase. DNA ladder (1 Kb) is shown in lane 1, and the no template control is shown in lane 2.
Expression of Human UGT1A8 and UGT1A10.
Western blot analysis was carried out on membranes derived from HK293 cells stably expressing UGT1A8 and 1A10. Results in Fig.2 show the 56kDa bands of the two UGT proteins in varying concentrations. The expression level of UGT1A8 in HK293 cells was approximately two-fold that of UGT1A10 as determined by densitometric analysis. Neither of these two UGT proteins was detected in nontransfected HK293 cells.

Western blot analysis of protein expression levels in membrane preparations from HK293 cells stably expressing human UGT1A8 and UGT1A10.
Western blots were analyzed as described in Experimental Procedures. Lanes 2 to 5 represent 50, 25, 12.5, and 6.25 μg of total protein from cell membrane preparations expressing UGT1A8. Lanes 6 to 9 represent 100, 50, 25, and 12.5 μg of total protein from cell membrane preparations expressing UGT1A10. Protein lysate from 50 μg of nontransfected HK293 cells was shown in lane 1.
Glucuronidation of Endobiotics by Stably Expressed Human UGT1A8 and UGT1A10.
The reactivity of human UGT1A10 with certain steroids has been described previously (Strassburg et al., 1998). We have shown that human UGT1A8, as transiently expressed, catalyzed the glucuronidation of catechol estrogens and certain androgens (Cheng et al., 1998a). The data in Table 1 show that stably expressed human UGT1A8 also catalyzes the glucuronidation of testosterone, epitestosterone, and estrogens, such as diethylstilbestrol and 17α-ethynyl estradiol. In contrast, pregnenolone and 17α-hydroxyprogesterone were not substrates for expressed UGT1A8. Interestingly, expressed UGT1A8 also catalyzed the glucuronidation of certain straight-chain saturated fatty acids, bile acids, and retinoids (Table 1). Of the fatty acids tested,n-decanoic acid was the best substrate. Fatty acids with longer or shorter alkyl chains were glucuronidated at lower rates than decanoic acid. Of over twenty endobiotic compounds investigated, stably expressed human UGT1A10 only showed glucuronidation activity toward 4- and 2-hydroxyestrone and dihydrotestosterone.
Glucuronidation of endobiotics catalyzed by stably expressed human UGT1A8 and 1A10
Glucuronidation of Xenobiotics by Stably Expressed Human UGT1A8 and UGT1A10.
A number of flavonoid xenobiotics are substrates for UGT1A8 and UGT1A10 (Table 2). Flavones, such as 7-hydroxyflavone, chrysin, and apigenin; the flavanone, naringenin; and the isoflavone, genistein, were glucuronidated by stably expressed human UGT1A8 and UGT1A10. However, stably expressed UGT1A8 displayed higher glucuronidation rates than UGT1A10 toward this chemical compound class.
Glucuronidation of flavonoids and anthraquinones by stably expressed human UGT1A8 and 1A10
Anthraflavic acid, emodin, alizarin, and other anthraquinone derivatives were glucuronidated by human UGT1A8 and UGT1A10 (Table 2). Of the substrates tested in the current study, alizarin appeared to be the most reactive for UGT1A8 and emodin was the best substrate for UGT1A10. In general, expressed UGT1A10 displayed lower glucuronidation activity toward these anthraquinones than UGT1A8.
Stably expressed human UGT1A8 also catalyzed the glucuronidation of all the phenolic compounds and coumarins tested, whereas stably expressed UGT1A10 only reacted with a limited number of substrates, including eugenol, mycophenolic acid, 4-hydroxybiphenyl, scopoletin, and 4-methylumbelliferone (Table 3). Neither stably expressed human UGT1A8 nor UGT1A10 catalyzed the glucuronidation of aliphatic alcohols, sapogenins, or nonsteroidal anti-inflammatory drugs (NSAIDs).
Glucuronidation of phenols and coumarins by stably expressed human UGT1A8 and 1A10
Membrane preparations derived from HK293 cells stably expressing UGT1A8 protein catalyzed the glucuronidation of opioids and other clinically important drugs or their metabolites (Table4). In contrast, glucuronidation of these compounds was not catalyzed by UGT1A10. Of the opioids tested, naltrexone, naloxone, and nalorphine showed higher glucuronidation rates with UGT1A8 than did morphine and buprenorphine. The observation that morphine was a substrate, but that codeine was not, suggests that expressed UGT1A8 catalyzes only the 3-O-glucuronidation of morphinan opioids.
Glucuronidation of opioids and other drugs catalyzed by expressed UGT1A8 not by UGT1A10
We have shown previously (Cheng et al., 1998a) that UGT1A8 did not catalyze the glucuronidation of NSAIDs. A number of other clinically useful drugs were tested to determine whether they were substrates for expressed UGT1A8. Of two fibrate hypolipidemic agents tested, the glucuronidation rate for ciprofibrate was higher than for clofibrate (Table 4). Other drugs or their metabolites, such as furosemide, diflunisal, 4-hydroxytamoxifen, phenolphthalein, and hexafluoro-2-propanol, were also substrates for expressed UGT1A8. Measurable activity was not detected for acetaminophen, chloramphenicol, (R)- and (S)-propanolol, and probenecid.
Kinetic Analysis of Glucuronide Formation Catalyzed by Stably Expressed Human UGT1A8 and UGT1A10.
Kinetic analysis of a number of representative compounds from the various classes of aglycones is shown in Table5. The glucuronidation rates of naringenin, propofol, 1-naphthol, and opioids were too low or nondetectable for UGT1A10, therefore the Kmand Vmax for these compounds have not been determined. A number of naturally occurring compounds such as scopoletin, alizarin, apigenin, and eugenol were found to have high glucuronidation efficiencies for both UGT1A8 and UGT1A10, suggesting that the contribution of the two intestinal UGTs in the conjugation of these natural products may be significant in vivo. Naltrexone and naloxone, two opioid antagonists, showed relatively low glucuronidation efficiencies for UGT1A8 compared with other xenobiotics (Table 5).
Comparison of kinetics of glucuronide formation in HK293 cells stably expressing UGT1A8 and UGT1A10
Discussion
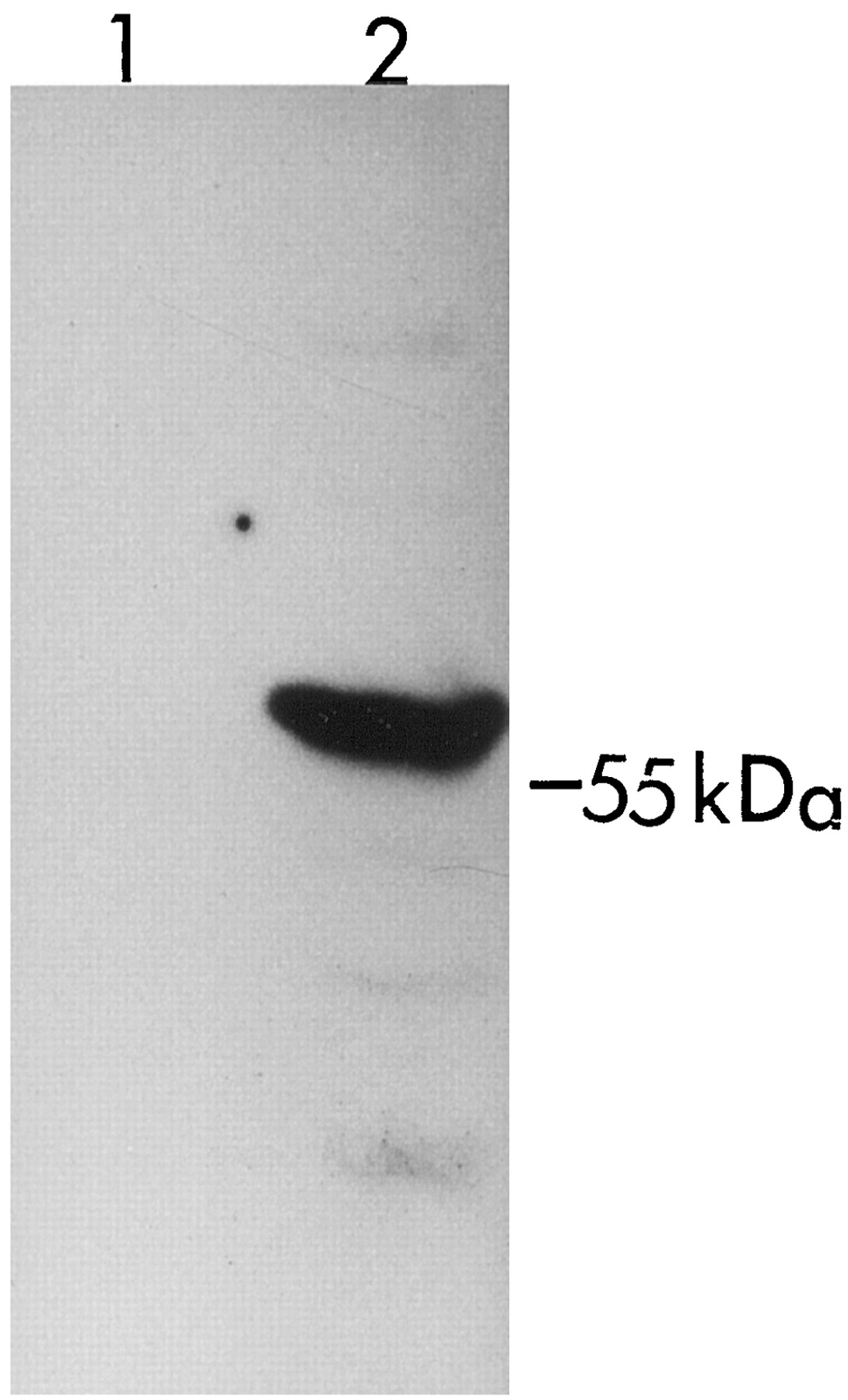
Full-length cDNAs of UGT1A8 and UGT1A10 were cloned from human small intestinal mucosa. Stably expressed UGT1A8 and UGT1A10 catalyzed the glucuronidation of a number of xenobiotics and endobiotics. UGT1A8 and UGT1A10 were previously expressed in Sf cells (Strassburg et al., 1998), transiently in COS cells (Strassburg et al., 1998; Mojarrabi and Mackenzie, 1998), or in HK293 cells (Cheng et al., 1998). COS cells have one or more intrinsic UGT1A isoforms (Fig.3) and endogenous UGT activity in COS cells obscures the detection of glucuronide formation when low rates are observed. Although the expression level of stably expressed UGT1A10 in HK293 cells was only half that of stably expressed UGT1A8, the catalytic activity of UGT1A8 toward anthraquinones, coumarins, flavonoids, and phenolic compounds was much higher than that of UGT1A10. Furthermore, stably expressed UGT1A8 showed glucuronidation activity with some compounds that were not substrates for UGT1A10. This includes bile acids, fatty acids, retinoids, opioids, and several clinically useful drugs or their metabolites such as ciprofibrate, furosemide, 4-hydroxytamoxifen, and phenolphthalein. The amino acid sequences of UGT1A8 reported in the Gen Bank (accession numberU42604), Strassburg et al. (1998), and Mojarrabi and Mackenzie (1998)differ from that which we have reported (Cheng et al., 1998). Based on this information, it would not be unexpected that variants of UGT1A8 exist in the human population. Many variants for other UGTs (e.g., UGT1A1) are already well known (Mackenzie et al., 1997). However, additional studies are needed to determine whether structural differences relate to functional differences with human intestinal UGT1A8.
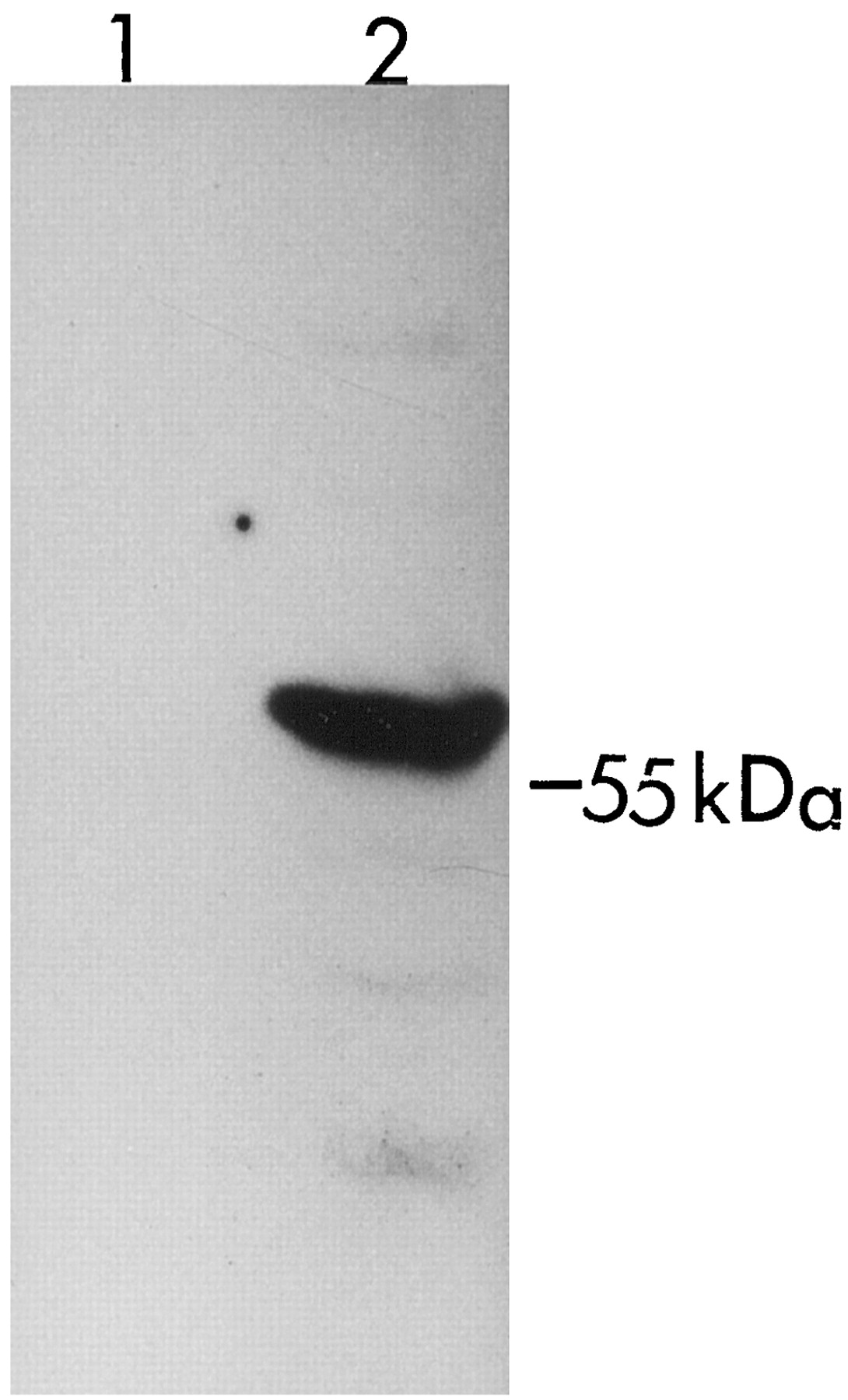
Western blot analysis of UGT1A protein expression in COS cells.
The proteins from whole cell homogenates were separated on a 10% SDS-polyacrylamide gel, transferred to a nitrocellulose membrane, and probed with a rabbit anti-UGT1A antiserum. Fifty micrograms of total proteins from HK 293 cells (lane 1) and COS cells (lane 2) are shown.
The bioavailability of natural products, toxic or carcinogenic compounds, and orally administered drugs may be reduced due to presystemic elimination dependent on UGTs. Although liver is considered to be a main drug-metabolizing organ in the body, the intestinal mucosa often represents the first exposure site to chemical substances and may play an important role in the first-pass metabolism of xenobiotics. Although the oxidative metabolic capacity of the intestinal mucosa is considerably lower than that of the liver, the activity of conjugation reactions in the gut may be close to that of the liver and in some cases may even exceed it (Back and Rogers, 1987). Furthermore, the surface area of the intestinal mucosa is enormous. We have shown that UGT1A8 and UGT1A10 were expressed in the mucosa of both the small intestine and the colon and expressed UGT1A8 and UGT1A10 displayed relatively broad catalytic ability toward both xenobiotic and endobiotic compounds. Therefore, these two UGTs may play important roles in the first step of inactivation and detoxification in vivo. The transcripts of other UGTs, including UGT1A1, UGT1A3, UGT1A4, UGT1A9, and UGT2B7, have also been identified from human intestinal mucosa (Strassburg et al., 1998; Radominska-Pandya et al., 1998). UGT2B7 is an important polymorphic UGT isoenzyme (UGT2B7Y and UGT2B7H), both of which catalyze the glucuronidation of catechol estrogens, hyodeoxycholic acid, androsterone, NSAIDS, and opioids (Ritter et al., 1990; Jin et al., 1993; Coffman et al., 1998). The catalytic efficiency of UGT2B7 with opioid substrates is much higher than that for UGT1A8 reported here. Therefore, UGT2B7 may play a more important role in the glucuronidation of opioids in humans, although UGT1A8 might also contribute to the overall intestinal metabolism of opioids.
UGT1A8 demonstrates activity toward the carboxyl function of RA and RA derivatives such as 5,6-epoxy-RA and 4-hydroxy-RA. In the present study with UGT1A8, only the carboxyl function of 4-hydroxy-RA was glucuronidated. This is the first demonstration that a human enzyme from the UGT1A subfamily glucuronidates retinoids. The major human isoform that glucuronidates retinoids on both carboxyl and hydroxyl functions is UGT2B7 (V. M. Samokyszyn, W. E. Gall, G. Zawanda, M. A. Freyaldenhoven, G. Chen and A.R.-P., unpublished data). UGT2B7 glucuronidates retinoids with a very high activity of 1 to 2 nmols/min/mg protein.
Coumarins are found in green plants, fungi, and bacteria (Murray et al., 1982). These naturally occurring compounds are excellent substrates for UGT1A8. Coumarins possess a variety of biological activities. For example, 4-methylumbelliferone and scopoletin have been found to act as scavengers of hydroxyl free radicals and to scavenge superoxide anions generated in the hypoxanthine oxidase system and esculetin has been shown to strongly inhibit lipid peroxidation (Páya et al., 1992). Glucuronidation of these compounds would be expected to limit their ability to inactivate reactive oxygen species. Our data shows that UGT1A8 can catalyze the glucuronidation of coumarins at high efficiency in the intestinal mucosa and might thereby decrease the bioavailability of these compounds to the body.
Expressed UGT1A8 and UGT1A10 also react with flavonoids found in food products. Naringin is the most prevalent flavonoid in grapefruit juice and imparts the characteristic bitter taste to the juice. Naringin is readily metabolized to the aglycone, naringenin, in the gut. Apigenin is found in parsley, genistein is found in soy foods, and quercetin is found in many different plants (Brown, 1980). Flavonoids have been shown to have many biological functions, including induction and inhibition of drug-metabolizing enzymes, inhibition of neutrophil degranulation, and inhibition of protein tyrosine kinase (Buening et al., 1981; Siess et al., 1989; Limasset et al., 1993). In addition, naringenin has been shown to increase the plasma concentration of many orally administered drugs by the inhibition of oxidative drug metabolism enzymes in the gastrointestinal tract (Ameer and Weintraub, 1997). The high glucuronidation efficiency observed for naringenin suggests that the contribution of UGT1A8 in the metabolism of this dietary compound and the regulation of its biological activity in the intestinal tract may be significant. Considering that the average American dietary consumption of flavonoids is ∼1 g/day (Pierpoint, 1986), possible food-drug interactions for glucuronidation cannot be ignored.
The cytochrome P-450-dependent activation of procarcinogens in the gut could specifically contribute to the formation of intestinal tumors (McKay et al., 1993). Intestinal UGT1A8 and UGT1A10 have been shown to metabolize carcinogenic benzo(α)pyrene and 2-acetylaminofluorene metabolites (Mojarrabi and Mackenzie, 1998). The existence of intestinal UGTs may be important in the detoxification of carcinogenic compounds in the intestine. The activity of most drug-metabolizing enzymes decreases slightly from proximal to distal small intestine, whereas in the mucosa of the large intestine a sharp fall in activity was observed (Peters et al., 1991). A low level of the detoxifying potential in the colon might be important with regard to the high rates of carcinogenesis in the colon. Therefore, induction of the intestinal UGTs could be important in the prevention of cancer formation in colon. Additional studies are needed to investigate the potential regulation of human UGT1A8 and UGT1A10 synthesis in the gastrointestinal tract.
Acknowledgments
We thank Mitchell Green for his advice in the preparation of the manuscript, and Gladys Rios and Chrystal Schroeder for their assistance.
Footnotes
-
Send reprint requests to: Dr. Thomas R. Tephly, 2–9452 Bowen Science Building, Department of Pharmacology, The University of Iowa, Iowa City, IA 52242. E-mail: Thomas-Tephly{at}uiowa.edu
-
This work was supported by National Institutes of Health Grants GM26221 (T.R.T.) and DK49715 and 51971 (A.R.-P.) and a grant from the Monsanto Company (T.R.T).
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- HK293
- human embryonic kidney 293
- RT-PCR
- reverse transcriptase-polymerase chain reaction
- NSAIDs
- nonsteroidal anti-inflammatory drugs
- RA
- retinoic acid
- Received December 31, 1998.
- Accepted June 15, 1999.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}