


Abstract
The elucidation of the individual contributions of the fourCYP3A genes to the overall CYP3A activity has been hampered by similarities in their sequence and function. We investigated the expression of CYP3A mRNA species in the liver and in various other tissues using gene-specific TaqMan probes. CYP3A4 transcripts were the most abundant CYP3A mRNA in each of the 63 white European livers tested and accounted on average for 95% of the combined CYP3A mRNA pool. CYP3A5 and CYP3A7 each contributed on average 2%, whereas CYP3A43 contributed 0.3% transcripts to this pool. Fourteen percent of livers exhibited an increased share of CYP3A5 transcripts (range 4–20%). These livers were either heterozygous for the marker of the CYP3A5 polymorphism, the CYP3A5*1A allele, or expressed very low levels of CYP3A4 mRNA. The CYP3A7 expression was bimodal, and it was increased in 15% livers. CYP3A4 was the dominant CYP3A in the intestine, followed by CYP3A5. CYP3A5 and CYP3A7, but not CYP3A4, were also expressed in the adrenal gland and in the prostate, whereas only CYP3A5 was detected in the kidney. These three tissues were shown to express much lower levels of pregnane X receptor mRNA than the intestine, indicating possibly a different mode of regulation of CYP3A expression. Expression of CYP3A genes was undetectable in peripheral blood lymphocytes. In summary, these assays and results should aid in our efforts to further dissect the regulation and the physiological and pharmacological significance of CYP3A isozymes.
The human cytochrome P450 3A (CYP3A) gene family comprises 4 genes, CYP3A4, CYP3A5, CYP3A7, and CYP3A43, which are arranged in tandem at a locus on chromosome 7 (Gellner et al., 2001). Together, protein products of CYP3A genes account for the largest portion of cytochrome P450 proteins in the liver (Thummel and Wilkinson, 1998). They are involved in the metabolism of a large proportion of all currently used drugs and many other compounds such as steroid hormones, toxins, and carcinogens (Li et al., 1995a; Evans and Relling, 1999). The hepatic expression and activity of CYP3A isozymes varies dramatically from individual to individual and also within the individual subject in a time and tissue-dependent manner (Ozdemir et al., 2000). This results in pharmacokinetic variability of drugs that are CYP3A substrates and significantly affects efficacy and adverse events of many medications. It has also been postulated that the variable expression of CYP3A could affect the predisposition of an individual to cancers caused by environmental procarcinogens that are bioactivated by CYP3A isozymes (Kirby et al., 1993).
Despite considerable efforts, our understanding of factors regulating CYP3A activity and expression remains limited. An important reason is the incomplete and in part conflicting estimates of the expression levels and activities of the four known CYP3A genes. For example, most analyses conclude that CYP3A4 forms the bulk of the hepatic CYP3A protein and activity (Thummel and Wilkinson, 1998), but this prevailing view has recently been challenged (Kuehl et al., 2001). The CYP3A5 protein has been reported expressed in between 10 and 97% of human livers at levels varying by up to an order of magnitude (Aoyama et al., 1989; Wrighton et al., 1990; Schuetz et al., 1994; Boobis et al., 1996;Jounaidi et al., 1996; Kuehl et al., 2001). We have reported recently that a several-fold difference in the calculated expression of CYP3A5 can be caused by the change of the antibody and protein standard used (Hustert et al., 2001). The third CYP3A, CYP3A7, was originally described in the human fetal liver where it accounts for about 50% of the total P450 protein (Wrighton et al., 1988). More recent reports suggest constitutive or induced expression of CYP3A7 in adult human livers (Schuetz et al., 1994; Greuet et al., 1996; Pascussi et al., 1999; Tateishi et al., 1999), but a final verdict has been hampered by the lack of specific antibodies. Similarly, no protein expression data are available for the recently identified fourth member of the family, CYP3A43 (Domanski et al., 2001b; Gellner et al., 2001; Westlind et al., 2001).
We set out to assess the variability in CYP3A expression on the mRNA level. Such an assessment should be sufficient to detect transcriptional activation of CYP3A genes in response to drugs (induction). All CYP3A genes have been reported to undergo induction by many drugs and environmental or dietary substances, and this phenomenon can lead to clinically relevant adverse drug interactions (Thummel and Wilkinson, 1998). In addition, since CYP3A genes appear to be regulated mostly at the transcription level (Telhada et al., 1992; Schuetz et al., 1993; Li et al., 1995b), mRNA quantification should give a fair estimate of the variability of their expression.
Interestingly, some, albeit not all, workers in the field have reported the expression and even the induction of CYP3A genes in peripheral blood leukocytes (Janardan et al., 1996; Hukkanen et al., 1997; Sempoux et al., 1999; Starkel et al., 1999; Krovat et al., 2000;Nakamoto et al., 2000; Asghar et al., 2002; Finnstrom et al., 2002;Nowakowski-Gashaw et al., 2002). These results have raised the exciting prospect of prediction of hepatic CYP3A expression of an individual based on an assay performed on a blood sample.
Several efforts have been undertaken to determine the expression of the mRNA derived from the individual CYP3A genes, but a comprehensive analysis has not been reported. This task has become easier following the recent elucidation of the sequence and structure of the human CYP3A locus (Finta and Zaphiropoulos, 2000; Gellner et al., 2001). In this paper, we describe the results of our efforts to develop real-time quantitative PCR2 methods, which are based on reverse transcription followed by PCR and detection based on fluorescence quenching using the TaqMan chemistry. These assays were developed for highly specific quantification of each of the four homologous CYP3A mRNA species. We also devoted significant efforts to develop procedures for normalization of mRNA concentrations, in order to enable comparisons of the absolute expression levels of the individual CYP3A genes in liver and various other tissues.
Materials and Methods
Samples.
Blood and matching liver samples were collected during surgical interventions conducted at the Department of Surgery, University Medical Center Charité, Humboldt University in Berlin, Germany. The donors were white Europeans. The liver samples included nontumorous tissue surrounding primary liver tumors and metastases of various tumors or liver material surgically removed for other reasons. Medical histories were evaluated in respect of treatment with known CYP3A inducers (see also Discussion). The study was approved by the local ethics committee and conducted in accordance with the Declaration of Helsinki. For the preparation of high quality RNA, a small tissue piece of 5 mm maximal side length was cut out immediately from each liver specimen and immediately transferred into a vial containing 1 ml of RNAlater (Ambion, Austin, TX). The samples were first kept at 4°C for 24 h to promote inactivation of RNases by the reagent and then stored frozen at −80°C for maximally 2 months until total RNA was isolated. Pools of mRNAs derived from extrahepatic tissues were purchased from CLONTECH (Palo Alto, CA).
RNA Isolation and cDNAs Synthesis.
Following homogenization, total RNA was isolated using RNeasy kit (Qiagen GmbH, Hilden, Germany). Concentration of total RNA was determined photometrically. cDNA from liver samples as well as from the CLONTECH tissue panel was synthesized from 1 μg of total RNA using random hexamer primers (0.1 A260 units), dNTPs (0.3 mM), and 50 U of SuperScript reverse transcriptase (Invitrogen, Carlsbad, CA) in a total volume of 30 μl using the buffer and reaction conditions provided by Invitrogen. cDNA samples were diluted to a final concentration of 10 ng/μl of the total RNA taken for reverse transcription, aliquoted, and stored at −80°C.
Preparation of CYP3A and hPXR Standards and Calibration Curves.
Plasmids containing CYP3A4, CYP3A5, CYP3A7, CYP3A43, and human pregnane X receptor (hPXR) cDNAs were obtained by PCR amplification and subcloning of reverse-transcribed liver mRNA using gene-specific primer pairs. Following linearization, sense mRNAs were obtained by in vitro transcription using SP6 or T7 RNA polymerase (Promega, Mannheim, Germany). Plasmid DNA was removed through digestion with RQ-DNase (Promega), and transcribed RNA was purified by phenol/chloroform extraction and precipitation. RNA yield was determined photometrically, and the number of transcripts per microgram of in vitro RNA was calculated based on the length of the respective RNA. cDNAs species were then synthesized as described above and serial 10-fold dilutions from 106 to 100 copies were used as calibration curves.
TaqMan Assays.
Expression levels were quantified by real-time quantitative PCR using the Applied Biosystems (Foster City, CA) Sequence Detection System 7700 (SDS7700). Oligonucleotides and TaqMan probes were developed using Primer Express software (Applied Biosystems). TaqMan probes were labeled on 5′ end with the reporter dye 6-carboxy-fluorescein and on 3′ end with the quencher 6-carboxy-tetramethylrhodamin. Usually, cDNA derived from 10 ng of total RNA was used for one TaqMan reaction. In case of low copy numbers, the amount was increased to 40 ng. TaqMan PCR amplifications were performed using the Universal Mastermix from Applied Biosystems or the TaqMan reaction system from Eurogentec (Seraing, Belgium). The conditions consisted of 40 cycles of 15 s at 95°C and 1 min at 60°C, following an initial step of 10 min at 95°C. Transcript numbers were then calculated using linear regression analysis from their respective calibration curves. The expression levels of 18S rRNA, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), hypoxanthine-guanine phosphoribosyl transferase (HPRT), TATA-box binding protein (TBP), and β-glucuronidase (GUS) were determined using commercially available, predeveloped TaqMan assays (Applied Biosystems). Normalized expression data were obtained by dividing CYP3A or PXR transcript numbers by those of 18S rRNA, GAPDH, HPRT, TBP or GUS. All assays were performed in triplicates. Analyses of most samples were performed at least twice with independent reverse transcription and TaqMan analysis.
Statistics.
The significance of the differences between the mean transcript levels was assessed using a Student's t test. The level of significance was 0.05.
Results
Development and Basic Characteristics of CYP3A and PXR TaqMan Assays.
We developed one assay for each CYP3A5, CYP3A7, CYP3A43, and PXR as well as two assays for CYP3A4, henceforth referred to as CYP3A4-A and CYP3A4-B assays. To achieve the highest possible gene specificity, primers and probes for CYP3A assays were derived from regions with most differences to the corresponding portions in the other CYP3A genes. The uniqueness of primers and probes within the available human genomic sequences was verified by Basic Local Alignment Search Tool sequence comparisons against the high-throughput genomic sequences section of Genbank. The sequences of primers and probes are given in Table1. Gene specificity of the assays was then assessed experimentally by performing each assay on cDNAs derived from the other members of the human CYP3A gene family. The assay specificity was expressed for each gene pair as ratio of amplification of the target gene of the assay and of one of the other CYP3A genes. In 12 of 15 possible combinations of assays and gene pairs, the specificity was >106-fold higher with the appropriate cDNAs than with any of the respective other three cDNAs (Table 2). The CYP3A4/CYP3A5 ratio of the two CYP3A4 assays developed was 5 × 104 for the CYP3A4-A assay and 5 × 103 for the CYP3A4-B assay. The CYP3A5/CYP3A7 ratio of the CYP3A5 assay was 103. The CYP3A7 assay was the most specific one, since it did not detect any other CYP3A cDNA within the concentration range tested (from 100 to 107 copies of in vitro RNA).
Sequences of oligonucleotides comprising CYP3A and PXR TaqMan assays
Specificities of CYP3A TaqMan assays
The linear range of all CYP3A assays (as well as of the PXR TaqMan assay) was from 100 to 107 copies of in vitro RNA. The mean difference between two independent measurements was 13% (range 0–40%; n = 23). Finally, we determined the “recovery” of a known number of in vitro generated CYP3A4 transcripts that were added to otherwise identical preparations of liver RNA. The recovery was close to 100% (data not shown).
Evaluation of Two Different CYP3A4 TaqMan Assays.
Different TaqMan assays toward the same gene would be expected to yield similar results, but experimental evidence for this assumption is limited and theoretically, sensitivity of mRNA to endonuclease and exonuclease cleavage may differ in different regions. We investigated CYP3A4 expression in 17 different human liver samples using two different CYP3A4 TaqMan assays (Table 1). The amplicon generated in the CYP3A4-A assay contains portions of exons 1 and 3 and the entire exon 2. The amplicon of the CYP3A4-B assay contains portions of exons 2 and 4 and the entire exon 3. CYP3A4 transcript numbers obtained with these two assays using one cDNA preparation differed on average by 40%. All the following data were obtained with the CYP3A4-A assay.
Data Normalization.
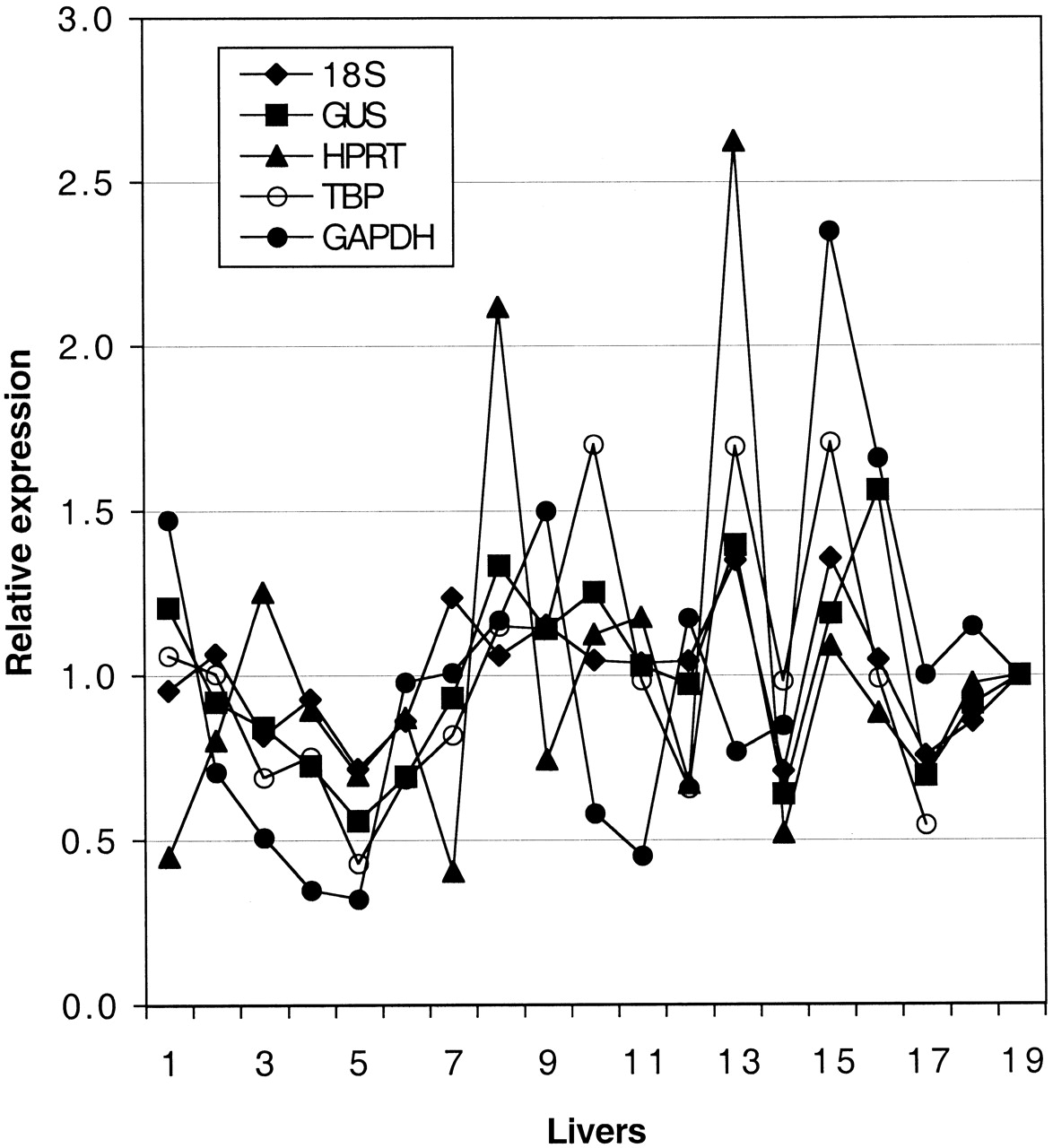
Data normalization, which is necessary to allow for comparisons between different samples and tissues, should be performed using a gene that is expressed at similar levels in these samples and tissues. We examined four housekeeping genes (GAPDH, HPRT, TBP, and GUS), which are frequently used for normalization purposes, using commercially available reagents developed by Applied Biosystems. To facilitate comparisons, the expression of every gene in each sample was expressed as a fraction of the mean expression of the respective gene in the entire sample set (Fig. 1). All four genes showed significant interindividual differences in expression levels, which were however not of the same direction and magnitude in all samples. For example, GAPDH levels, which differed 7-fold among the 18 liver samples tested, were elevated in samples that had low levels of other control genes, or vice versa. This showed that the result of normalization would be highly dependent on the choice of the particular control gene and that none of the housekeeping genes tested would be suitable to normalize expression data. The differences between liver samples were unlikely caused by degradation of RNA, since no signs of degradation were observed by inspecting the RNA quality. In addition, such degradation would be expected to have a similar effect on all genes.

Expression levels of GUS, HPRT, TBP, and GAPDH mRNA species and of 18S rRNA in a set of 18 liver samples.
The expression of every gene in each sample is expressed as a fraction of its mean expression in the entire sample set.
As next, we investigated the suitability of 18S rRNA as a basis for normalization. 18S rRNA is a highly abundant RNA that provides a good estimate of the total RNA content. The values for 18S rRNA varied less than 2-fold in the same set of 18 livers (i.e., it showed the lowest variability of all RNA species tested). Therefore, 18S rRNA expression was taken as a basis for normalization in all subsequent experiments.
Expression of CYP3A Genes in Human Livers.
Expression of mRNAs of the individual CYP3A genes was investigated in 63 liver samples obtained by surgery from individuals of white European origin. CYP3A4 expression varied by a factor of 118, and it was in the range between 11,000 and 1.3 × 106copies/ng of total RNA (Fig. 2). An average liver expressed 479,000 CYP3A4 copies/ng of total RNA. CYP3A5 mRNA expression varied by a factor of 32 (2,000–63,000 copies/ng, average 9,100 copies/ng of total RNA, n = 63). CYP3A7 expression varied by a factor of 333 with an average of 9,500 copies/ng of total RNA (range 170–56,600 copies/ng of total RNA,n = 63) and CYP3A43 by a factor of 50 (100–5,000 copies/ng of total RNA, on average 1,300 copies/ng of total RNA,n = 63).

Distribution of the expression levels of CYP3A mRNA species in 63 human livers.
Data shown in thousands of transcripts/ng of total RNA.
The expression of the CYP3A5 protein is bimodal, and this bimodality has been recently linked to a single nucleotide polymorphism in intron 3 (CYP3A5*3A allele) that adversely affects the splicing and stability of the gene transcripts (Hustert et al., 2001; Kuehl et al., 2001). We have analyzed the expression of CYP3A5 mRNA as a function of CYP3A5 genotype. Among the 63 liver samples investigated, 55 were homozygous for the CYP3A*3A allele whereas eight carried the CYP3A5*1A/CYP3A5*3A genotype. The *3A/*3A samples expressed on average 5,100 transcripts/ng of total RNA (range between 2,000 and 11,000 copies), compared with 41,200 transcripts expressed by the samples with *1A/*3A genotype (range 21,500 to 62,600 transcripts/ng of RNA; P < 0.002).
The correlation coefficient (r) values were below 0.14 for all six possible pairs of CYP3A genes. We then calculated the relative contributions of the individual genes to the combined CYP3A transcript pool. CYP3A4 was the dominant CYP3A in every liver sample screened (Fig. 3). On average, CYP3A4 transcripts accounted for 95% of all CYP3A transcripts (range 71–99.5%). CYP3A5 and CYP3A7 gene transcripts accounted on average for 2.5% of all CYP3A transcripts each. The highest share in the CYP3A transcript pool for either gene was 20% (Fig. 3). All eight samples with a CYP3A5*1A/CYP3A5*3A genotype were within the group of 10 samples with the highest share of CYP3A5 in the CYP3A transcript pool (range 4–20%). The remaining two samples in this group had a CYP3A5*3A/CYP3A5*3A genotype and low absolute numbers of CYP3A5 transcripts of 3700 and 3200 transcripts/ng of total RNA. The higher shares of CYP3A5 transcripts in the CYP3A transcript pool in these samples (5 and 20%) were caused by a low expression of CYP3A4 (69500 and 11400 transcripts/ng of total RNA, respectively). CYP3A43 transcripts accounted on average for only 0.3% of all CYP3A transcripts (range 0.02–1.2%).

Relative contributions of CYP3A genes to the combined CYP3A transcript pool in an average liver and in livers with the highest expression of CYP3A4, CYP3A5, and CYP3A7.
Expression of CYP3A Genes and PXR in Human Tissues.
Expression of CYP3A genes was investigated in pooled RNA preparations from selected extrahepatic human tissues. The expression patterns of CYP3A4, CYP3A5, and CYP3A7, shown in Fig.4 reveal similarities as well as unexpected and functionally, highly interesting features of tissue specificity. All three genes were expressed in the intestine. CYP3A5 and CYP3A7, but not CYP3A4, were found in the adrenal gland and in the prostate, whereas CYP3A5 was the only CYP3A detected in the kidney. We then quantified the expression of the PXR, which is involved in induction of CYP3A transcription. Consistent with previous reports (Bertilsson et al., 1998; Lehmann et al., 1998), PXR mRNA was detected in the liver (data not shown) and in the intestine. The TaqMan assay used detects both the full-length PXR and the splice variant PXR.2 that has 111 nucleotides deleted from the ligand-binding domain (Dotzlaw et al., 1999). A semiquantitative reverse transcription-PCR analysis revealed a very low contribution of this splice variant to the PXR expression in these tissues (data not shown). The expression of the gene in the adrenal gland, in the kidney, and in the prostate was, in comparison, negligible (Fig. 4). The results for CYP3A43 were below the lower limit of the linear range of the assay (not shown).

The expression of CYP3A4, CYP3A5, CYP3A7, and PXR in selected extrahepatic tissues.
Data shown in thousands of transcripts/ng of total RNA. The numbers in parentheses depict the number of organs from which total RNA was isolated and pooled by the manufacturer of the panel.
Expression of CYP3A Genes in Human Peripheral Blood Lymphocytes.
The expression of CYP3A4 was investigated in total RNA isolated from lymphocytes of 26 healthy volunteers prior to and 24 h after a 5-day treatment with the CYP3A inducer rifampin. CYP3A4 transcripts could not be detected in the peripheral blood mononuclear cells under either condition. Similar results were obtained with CYP3A5, CYP3A7, and CYP3A43 assays (data not shown).
Furthermore, we investigated the expression of CYP3A in a set of 11 liver biopsies and matching leukocytes (Hoffmeyer et al., 2000). CYP3A4, CYP3A5, and CYP3A7 were detected in all 11 liver biopsies. CYP3A4 was expressed at an average of 88,000 transcripts/ng of total RNA (range between 33,000 and 148,000 transcripts/ng of total RNA). The expression of CYP3A5 was between 200 and 500 transcripts/ng of total RNA for homozygous CYP3A5*3A samples and between 2200 and 4200 transcripts/ng of total RNA in two livers with the CYP3A5*1A/CYP3A5*3A genotype. The expression of CYP3A7 was between 100 and 15600 transcripts/ng of total RNA in this sample set. The lower expression levels of CYP3A4, CYP3A5, and CYP3A7 in these livers in comparison with livers shown in Fig. 2 were most likely due to the different methods of total RNA isolation. In contrast, no expression of CYP3A4 or other CYP3A genes was detected in total RNA isolated with the same method from matching leukocytes. These results indicated that for all practical purposes, CYP3A expression cannot be reliably assessed in peripheral leukocytes.
Discussion
The differentiation between CYP3A isozymes at the level of protein expression or activity has been hampered by lack of specific antibodies and activity probes. Therefore, we attempted to quantify CYP3A expression at the mRNA level by means of quantitative real-time reverse transcription-PCR assays. First of all, we turned our attention to the normalization of assay results, which is necessary to compare expression levels of different genes and among different samples. Working with frozen samples of solid tissues practically precludes the calculation of the expression results per cell. Typical other possibilities include normalization using either total RNA content or mRNA content of the samples. However, these options do not allow correction for sample-to-sample variations in RNA quality or cDNA synthesis. Similarly to most other investigators, we first used the so-called “housekeeping genes”, which are assumed by many to be expressed at constant and comparable levels. However, all of the housekeeping genes investigated showed large interindividual differences in the expression levels. These observations are in agreement with the increasing number of reports describing regulatory elements in the promoter regions of such housekeeping genes as β-actin, GAPDH, and HPRT (for a review, see Bustin, 2000). For the following reason, we chose to normalize our results using a TaqMan assay for 18S rRNA. Due to its abundance, 18S rRNA is expected to provide a good estimate of the total RNA content of the sample. In agreement with this assumption, 18S rRNA showed the lowest (<2-fold) variability of all RNA species tested. In comparison with normalization using the photometrically determined RNA content, normalization with 18S rRNA has the advantages of taking into account the integrity of the RNA molecules and any differences in the efficiency of the reverse transcription step.
Several lines of evidence suggest that the results obtained with our assays accurately reflect the expression of CYP3A transcripts in the samples investigated. The recovery is close to 100%, as revealed by analysis of samples differing by a defined number of transcripts. Furthermore, the average expression of CYP3A4 (479,000 transcripts/ng of total RNA) is very similar to that obtained by an independent group using a different quantitative PCR method [300,000 transcripts/ng of total RNA (Rodriguez-Antona et al., 2000)]. Lastly, two separate CYP3A4 assays gave comparable results.
A reliable and affordable test for hepatic and/or intestinal CYP3A expression would be beneficial to drug development and for drug therapy. Several laboratories have investigated the expression of CYP3A in peripheral blood leukocytes or their subtypes, but the obtained results are equivocal (Janardan et al., 1996; Hukkanen et al., 1997;Sempoux et al., 1999; Starkel et al., 1999; Krovat et al., 2000;Nakamoto et al., 2000; Asghar et al., 2002; Finnstrom et al., 2002;Nowakowski-Gashaw et al., 2002). With the specific and sensitive TaqMan assays we developed, none of the CYP3A transcripts was detectable in peripheral blood leukocytes. This result was verified in several ways. Leukocytes with matching liver samples obtained from the same individuals revealed no expression, although the transcript numbers of the liver samples were within the range of other liver samples tested. Second, in a series of leukocytes obtained from individuals treated with rifampin at a regimen that induces hepatic CYP3A4 expression, we found no discernible effect on CYP3A mRNA expression in leukocytes. Therefore, although CYP3A genes may be expressed at low levels in peripheral blood leukocytes, these cells cannot be applied to predict their hepatic expression levels.
None of the CYP3A mRNAs exhibits a truly unimodal distribution in the liver set analyzed. The most pronounced bimodality was apparent in the histograms of CYP3A5 and CYP3A7. The histograms of CYP3A5 and CYP3A7 expression revealed that approximately 10 and 15% samples, respectively, express much higher transcript numbers of either gene. In the case of CYP3A5, all these samples are heterozygous for the CYP3A5*1A allele, which has been linked to the increased expression of the CYP3A5 protein (Hustert et al., 2001; Kuehl et al., 2001). The expression of CYP3A7 was originally thought to be restricted to fetal livers (Wrighton et al., 1988), but its detection in adult livers in this study is in agreement with more recent reports (Schuetz et al., 1994; Greuet et al., 1996; Pascussi et al., 1999; Tateishi et al., 1999). CYP3A7 gene transcripts reach one-fifth of the combined CYP3A transcript pool, but in most samples their contribution remains low. In some samples, CYP3A expression may have been affected by drug exposure prior to surgery. Thus an analysis of donor medical histories revealed exposure to omeprazole and pantoprazole in two samples expressing high levels of CYP3A5 mRNA and to nifedipine and omeprazole in two samples expressing high levels of CYP3A7 mRNA. The least important CYP3A appears to be CYP3A43, at least as judged from its low expression levels.
CYP3A4 transcripts account for 95% of all CYP3A transcripts in an average liver. Higher contributions of CYP3A5 or CYP3A7 in some livers do not necessarily reflect high transcript numbers for these genes but can be also caused by low CYP3A4 transcript numbers. This is consistent with the relatively low degree of correlation of the expression levels among the CYP3A family members. This observation was a little surprising, considering that these genes share transcriptional regulators, with the orphan nuclear receptor PXR being the most prominent example (Bertilsson et al., 1998; Lehmann et al., 1998).
On the other hand, CYP3A genes exhibit a degree of tissue-specific expression, which could reflect gene-specific differences in their regulation, at least in the extrahepatic tissues. In support of this possibility, CYP3A5 and CYP3A7, but not CYP3A4, are expressed in the adrenal gland and in the prostate, whereas CYP3A5 alone is detected in the kidney. Considering the relatively low frequencies of hepatic CYP3A5 and CYP3A7 polymorphisms, the failure to detect these genes in some organs could have been caused by the relatively low numbers of individuals screened.
The low level of PXR expression in the adrenal gland, prostate, and kidney demonstrates that expression of these CYP3A isozymes does not strictly depend on PXR in these three tissues. Instead, the CYP3A expression could be mediated by Vitamin D receptor (Schmiedlin-Ren et al., 2001) or by the constitutively activated receptor (Sueyoshi et al., 1999; Pascussi et al., 2001). This possibility remains to be verified in further studies.
Footnotes
-
↵1 Present address: Department of Clinical Pharmacology, Georg-August University Goettingen, Robert-Koch-Str. 40, D-37075 Goettingen, Germany. E-mail:Leszek.Wojnowski{at}med.uni-goettingen.de
-
Parts of this work were supported by Grants 01GG9846 and 01GG9848 from the German Federal Ministry for Education and Science (BMBF) and by the Robert Bosch Foundation, Stuttgart, Germany.
- Abbreviations used are::
- PCR
- polymerase chain reaction
- cDNA
- complementary DNA
- PXR
- pregnane X receptor
- rRNA
- ribosomal RNA
- GAPDH
- glyceraldehyde 3-phosphate dehydrogenase
- HPRT
- hypoxanthine-guanine phosphoribosyl transferase
- TBP
- TATA-box binding protein
- GUS
- β-glucuronidase
- Received March 20, 2002.
- Accepted July 2, 2002.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}