


Abstract
Sixty-four compounds with diverse structures were used in evaluation of intrinsic clearance by various hepatocyte preparations from rats, dogs, monkeys, and humans. Intrinsic clearance (CLint) was calculated from the ratio of the initial amount of the test compound minus the amount remaining after 2 h of incubation and the corresponding area under the concentration versus time curve. The predictive potential of this in vitro model was tested by comparing the intrinsic clearance with in vivo clearance using linear regression analysis. In addition, the intrinsic clearance values obtained from three different types of hepatocytes (cryopreserved, fresh, and sandwich-cultured) from the same species were compared to determine the influence of preservation and culture conditions. It seems that intrinsic clearance determined from human cryopreserved hepatocyte (R2 = 0.87) was the best predictor for the corresponding human in vivo clearance. Dog and rat hepatocyte clearances were also demonstrated to be reasonable predictors (R2 ranged 0.68–0.85 in dogs and 0.65–0.72 in rats) for their corresponding in vivo clearances. Monkey hepatocyte clearance seems to be the worst predictor for the corresponding in vivo clearance (R2 = 0.35–0.67). Comparison of intrinsic clearance generated from cryopreserved and fresh hepatocytes demonstrated a very good correlation in dog (R2 = 0.82) followed by rat (R2 = 0.77), and then by monkey (R2 = 0.70). A similar correlation profile was shown between cryopreserved hepatocytes and sandwich-cultured hepatocytes. These results demonstrated the predictive potential of intrinsic clearance for rat, dog, and human in vivo clearance, but also showed some limitation of the approach for monkey.
In the early phase of drug discovery, animal pharmacokinetic data are used for advancing drug candidates to the clinical phase of drug development. According to a study by Prentis et al. (1988), approximately 40% of drug candidates failed because of poor human pharmacokinetic characteristics. For this reason, numerous attempts have been performed in recent years to find a reliable way of predicting in vivo hepatic clearance in humans. For example, Lavé et al. (1997) demonstrated a correlation between intrinsic clearance in human hepatocytes and hepatic extraction ratio in humans, and used the correlation to classify compounds into low, intermediate, or high hepatic extraction ratio in humans. Obach (1997a)used hepatic microsomal intrinsic clearance to predict human in vivo clearance and Houston and Carlile (1997) compared the use of microsomes, hepatocytes, and liver slices for prediction of hepatic clearance in rats. They showed that both microsomes and hepatocytes might be useful for obtaining the rank order of clearances. Schneider et al. (1999) used multivariate statistical models and artificial neural networks to predict the hepatic drug clearance in humans from in vitro hepatocyte data. These in vitro models yielded a wide range of in vivo-in vitro correlation indexes that, in general, suggest the potential for prediction of in vivo human clearance using in vitro clearance data.
In addition to developing various in vitro physiological models for the evaluation of in vivo-in vitro correlations, considerable effort has been made toward methods for calculation of intrinsic clearance. In some studies, intrinsic clearance was obtained by determining the enzyme kinetic parameters (Vmax andKm). The ratio of these two parameters was then equated with intrinsic clearance. In the second method, referred to as the “in vitro t1/2method”, intrinsic clearance (CLint) was calculated by measuring the first order rate constant for consumption of the substrate at low concentration. In a third method, intrinsic clearance was calculated from the ratio of the initial amount of the test compound and the corresponding area under the curve (AUC0-∞1), which was calculated by extrapolation to time infinity. In addition, some recent studies also take into consideration the hepatic flow rate and the protein binding in plasma (Lavé et al., 1997).
In this study, we assumed that the in vitro clearance process follows first order kinetics and accordingly, developed an equation for calculating the intrinsic clearance from the ratio of the initial amount minus the amount after 2 h of incubation and the AUC between 0 and 2 h. This procedure allows us to rapidly determine the in vitro intrinsic clearance for a large number of compounds without going through extensive extrapolation and linear regression analysis. The usefulness of this calculation method is further demonstrated by satisfactory correlation between in vivo and in vitro clearance in rats, dogs and humans, validating the use of this in vitro screening procedure to provide useful information on human pharmacokinetic behavior.
Materials and Methods
Materials.
The 38 compounds for animal studies were discovery compounds from Schering Plough Research Institute (Kenilworth, NJ), and the 26 compounds for human studies were randomly selected from the literature and obtained from Sigma-Aldrich (St. Louis, MO) with the exception of sildenafil (obtained from Schering Plough Research Institute, Kenilworth, NJ). Cryopreserved hepatocytes were obtained from In Vitro Technologies (Baltimore, MD), and fresh and sandwich-cultured hepatocytes were obtained from Cedra (Austin, TX). Waymouth's 752/1 medium, with l-glutamine and without phenol red was obtained from Mediatech (Herndon, VA).
In Vivo Pharmacokinetic Data.
In vivo clearance data in rats, dogs, and cynomolgus monkeys were generated in-house after intravenous (0.6- to 10-mg/kg dose) administration to three or more animals per compound. Plasma samples were collected at various time points and analyzed. Total body clearances were calculated from the ratio of dose/AUC. Human in vivo clearance values were obtained from the literature.
Preparation of Hepatocytes.
Cryopreserved hepatocytes were thawed in a water bath at 37°C and transferred to a 50-ml tube containing 15 ml of medium at 4°C. The hepatocytes were centrifuged at 750 rpm at 4°C for 5 min. The supernatant was removed and the hepatocytes were washed two more times with medium at 4°C. The hepatocyte pellet was gently resuspended in medium to a final density of 2 million cells/ml. Fresh hepatocyte suspensions were shipped in 50-ml tubes containing a gel matrix. The hepatocyte suspension was thawed in a water bath at 37°C and transferred to a 50-ml tube containing 25 ml of medium at 37°C. The hepatocytes were washed and resuspended in the same way as cryopreserved hepatocyte. The sandwich-cultured hepatocyte plates shipped in a gel matrix were thawed in the incubator at 37°C and washed with medium twice before use.
Cryopreserved animal hepatocytes were received as pooled from five to seven individuals; therefore, no pooling was needed. For the cryopreserved human hepatocytes, each lot was from a single individual; therefore, five or more different lots were pooled before incubation. Because fresh animal hepatocytes were from a single individual, pooling was not possible; therefore, clearance studies were done with two separate lots. The viability of each hepatocyte preparation was determined by trypan blue staining method immediately before use and after 2 h of incubation.
In Vitro Hepatocyte Incubations.
Incubations were carried out at a final test compound concentration of 1 μM for cryopreserved and 2 μM for fresh hepatocytes. Stock solutions of the compounds were prepared in methanol and diluted to the desired concentrations before adding to the hepatocytes. Incubations were carried out with a hepatocyte concentration of 1 million cells/ml in Waymouth's medium. Samples (1 ml) were incubated in 24-well plates at 37°C for 2 h in a CO2 incubator. At 0, 10, 20, 40, 60, and 120 min a 100-μl aliquot was removed and added to a 96-well plate containing 200 μl of acetonitrile with internal standard.
For sandwich-cultured hepatocytes, incubations were carried out at a final test compound concentration of 2 μM and with approximately 2 × 105 viable cells/well (48 wells). A volume of 200 μl of each test compounds was added to each well and incubated at 37°C for 2 h in a CO2incubator. At 0, 10, 20, 40, 60, and 120 min the contents of the wells were removed and added to a 96-well plate containing 200 μl of acetonitrile with internal standard. The wells were then washed with an additional 200 μl of acetonitrile and added to the same 96-well plate. The incubations were done with each type of hepatocyte using at least two different lots.
All samples were sonicated for 3 min and centrifuged at 4000 rpm for 20 min, and the supernatant was transferred into a 96-well plate. The plate was stored at −20°C until analyzed. Standard curves were also prepared in hepatocytes and analyzed with the samples.
HPLC/Atmospheric Pressure Chemical Ionization/MS.
The HPLC system consisted of an LC-10ADvp pump (Shimadzu, Koyoto, Japan) and Leap injector (Leap Technologies, Carrboro, NC). The column used was Genesis C182.0 × 50 mm (Jones Chromatography, Lakewood, CO). The mobile phase was a gradient with A (0.01 M ammonium acetate in 80:20 water/methanol) and B (0.01 M ammonium acetate in methanol). The flow rate was 0.35 ml/min. The initial liquid chromatography conditions were held at 40% B for 0.5 min, followed by a linear gradient from 40 to 95% B over 0.5 min, followed by 95% B for 2 min and back to 40% B over 0.5 min. The samples were analyzed in the positive ion mode using the heated nebulizer interface of a PE/Sciex 3000 (PerkinElmer Instruments, Norwalk, CT) triple quadrupole mass spectrometer with nebulizer probe temperature at 400°C.
Calculation of in Vitro Intrinsic Clearance.
Hepatocyte intrinsic clearances were calculated using the following formula:
Results
HPLC/MS-MS is a very important technique in drug discovery. It provides excellent sensitivity and selectivity, with a short analysis time. It also permits the high-throughput screening of large number of compounds for stability in the support of drug discovery. In the present study, all compounds were run with the same column, mobile phases, and gradient with a 3-min run time. No additional chromatographic method development was necessary for individual compounds. With the excellent selectivity of the mass spectrometer, the compounds could be separated by their characteristic parent and fragment ion masses and quantitated with minimal chromatographic separation.
The viability of each hepatocyte preparation was determined before and after incubation. Data suggested that the viability of the fresh and cryopreserved hepatocytes before incubation ranged from 85 to 95% and 50 to 70%, respectively. The viability remained the same after 2 h of incubation. Plated hepatocytes seem to be almost 100% viable before and after incubation.
The data provided by the supplier for both cryopreserved and fresh hepatocytes showed that phase I and II enzymes were still present and active. In addition, the same hepatocyte preparations have been used for in-house metabolite identification studies, both phase I and II metabolites were observed (data not shown). This indicates that both phase I and II enzymes are active in both fresh and cryopreserved hepatocytes.
The in vitro measurements of the rate of clearance were based on substrate disappearance over a 2-h incubation time. The CLint was calculated using eq. 1 instead of the traditional way of extrapolating to infinity to determine the AUC0-∞. The total in vivo plasma clearance values for rat, dog, and monkey (CLtot) were determined experimentally from in-house i.v. data. In vivo human clearance data were obtained from literature references. Assuming that the drug is metabolized or excreted only by the liver and kidney, CLtot is the sum of the hepatic (CLhep) and renal (CLrenal) clearance. Therefore, hepatic clearance (CLhep = CLtot − CLrenal) was used in the correlation to CLint. A linear regression curve fitting was used for all correlation analyses and the regression square (R2) was determined.
Rat, Dog, and Monkey Correlations.
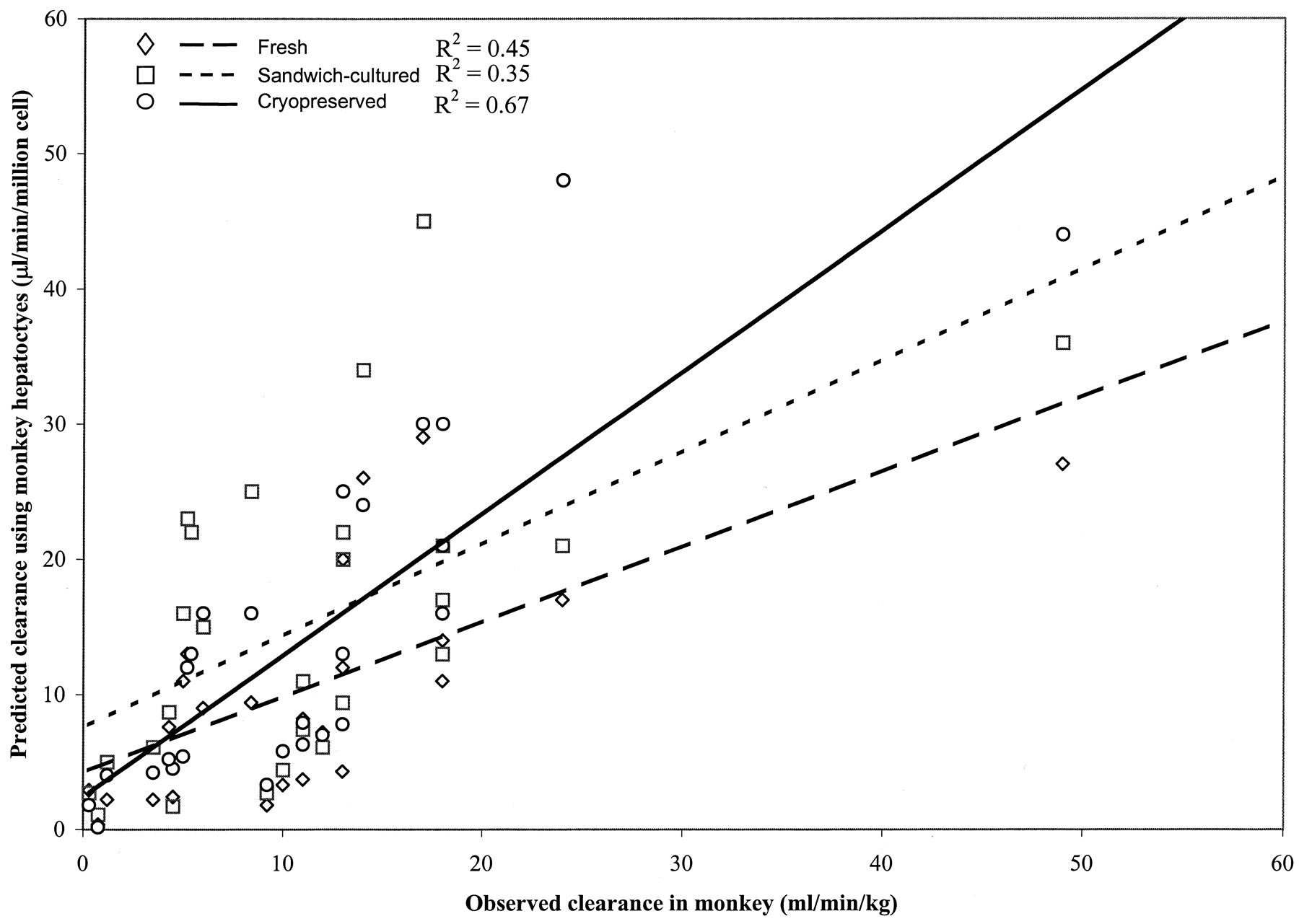
To avoid the extensive use of expensive and time-consuming animal studies, it is more strategic to establish an in vitro system that can screen out compounds with poor animal pharmacokinetics. A total of 27, 18, and 26 Schering compounds for rat, dog, and monkey, respectively, were dosed intravenously to determine their in vivo clearance values. The in vitro clearances for these compounds were determined using cryopreserved, fresh, and sandwich-cultured hepatocytes. The in vitro intrinsic clearances and the in vivo clearances for rats, dogs, and monkeys are summarized in Tables 1,2, and 3. The in vitro intrinsic clearance values are plotted versus actual clearance values in Figs. 1,2, and 3. Good correlations were obtained between in vitro clearance and in vivo clearance values. For rats, the R2value for fresh, sandwich-cultured, and cryopreserved hepatocytes was 0.72, 0.72, and 0.65, respectively. For dogs, theR2 value for fresh, sandwich-cultured, and cryopreserved hepatocytes was 0.85, 0.68, and 0.71, respectively. However, for monkeys, much poorer correlations were seen, with theR2 value for fresh, sandwich-cultured, and cryopreserved hepatocytes only 0.45, 0.35, and 0.67, respectively.
Summary of predicted (in vitro) and observed (in vivo) clearance in rat
Summary of predicted (in vitro) and observed (in vivo) clearance in dog
Summary of predicted (in vitro) and observed (in vivo) clearance in monkey

Correlation of predicted (in vitro) and observed in vivo clearance using fresh, sandwich-cultured, and cryopreserved rat hepatocytes.

Correlation of predicted (in vitro) and observed in vivo clearance using fresh, sandwich-cultured, and cryopreserved dog hepatocytes.

Correlation of predicted (in vitro) and observed in vivo clearance using fresh, sandwich-cultured, and cryopreserved monkey hepatocytes.
Cryopreserved hepatocytes are more readily available than fresh hepatocytes. Therefore, it is necessary to determine whether cryopreserved hepatocytes can be used instead of fresh hepatocytes to provide data that could predict in vivo clearances. The in vitro values obtained from all three animal species using fresh and cryopreserved hepatocytes were compared. The results are shown in Table4. Very good correlations were observed between intrinsic clearance data from fresh and cryopreserved hepatocytes with R2 value equal to 0.77, 0.82, and 0.70, for rat, dog, and monkey, respectively. A similar correlation was also obtained using sandwich-cultured and cryopreserved hepatocytes. Figure 5 contains plots of accuracy of in vitro clearance values using rat, dog, and monkey hepatocytes versus the respective in vivo clearance values. All data points in rats and dogs were within 2-fold error. In monkeys, a few low-clearance compounds fall outside the 2-fold error boundaries, indicating less correlation between the in vivo and in vitro data.
Summary of the correlation of clearances obtained from fresh and EHS sandwich gel versus clearances obtained from cryopreserved hepatocytes

Accuracy of clearance values predicted from rat, dog, monkey, and human hepatocytes versus actual in vivo clearance values.
a, rat hepatocyte studies; b, dog hepatocyte studies; c, monkey hepatocyte studies; d, human hepatocyte studies. Fresh hepatocyte (⋄), sandwich-gel hepatocyte (■), and cryopreserved hepatocyte (○). n is the number of compounds used in the study. Dotted lines signify 2-fold errors between in vitro and in vivo clearance values using in vivo data as reference point. Errors for tenoxicam and warfarin in d are well off scale (2000–6500% error) and are not depicted.
Human Correlation.
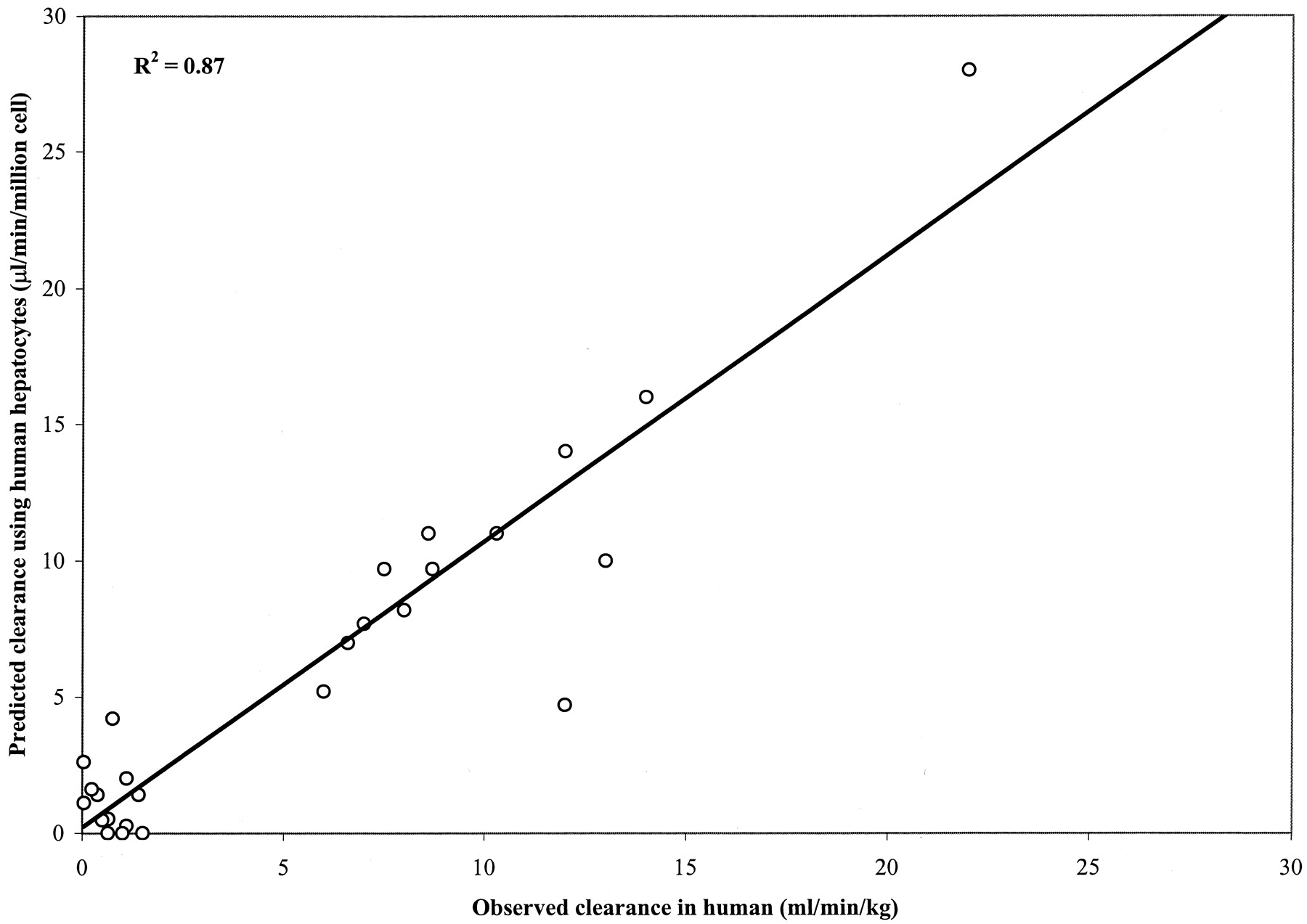
An in vitro-in vivo correlation to human clearance values is particularly important in the screening of new chemical entities in drug discovery because in vitro data are the only human information available before clinical trials. Twenty-six compounds with published in vivo clearance values were randomly selected from the literature and obtained from Sigma-Aldrich. In vitro intrinsic clearance values were determined from the incubation of pooled cryopreserved hepatocytes. A very good correlation was obtained between in vitro and in vivo clearances, with R2 = 0.87. A summary of the human intrinsic clearances obtained in vitro is presented in Table 5. The in vitro intrinsic clearance values are plotted versus actual clearance values in Fig.4. The accuracy of in vitro clearance values using human hepatocytes versus the respective in vivo human clearance values was plotted in Fig. 5d. Only five data points with very low in vivo clearances fall outside the 2-fold error boundary.
Summary of predicted (in vitro) and observed (in vivo) clearance in human using cryopreserved hepatocytes

Correlation of predicted (in vitro) and observed in vivo clearance in human using cryopreserved hepatocytes.
Discussion
With recent advances in combinatorial chemistry, thousands of compounds are going through drug discovery. To decrease the amount of expensive and time-consuming animal studies and be able to exclude compounds with undesirable human pharmacokinetics, a validated in vitro system is needed. The most common in vitro systems used include incubations of compounds in microsomes, hepatocytes, and liver slices. Microsomal studies are simple, cheap, and easy to conduct; however, microsomes only carry out phase I metabolism when supplemented with NADPH and therefore, this system is only predictive when oxidative metabolism predominates over other phase II metabolic routes such as conjugation, reduction, and hydrolysis. Incubations in liver slices are poor predictors of the in vivo state (Houston and Carlile, 1997) due to delayed accessibility of substrate to the cells within the core of the slices. In addition, human liver slices are difficult to obtain and the results are not reproducible. According to Houston and Carlile (1997), freshly isolated hepatocytes show high fidelity and provide accurate predictions of in vivo clearance. In the present study, satisfactory correlations were obtained not only from fresh hepatocyte suspensions but also with sandwich-cultured and cryopreserved hepatocytes, indicating that cryopreserved hepatocytes have predictive power similar to that of fresh hepatocytes. The good correlation to human in vivo clearance values provides the basis for wider use of the more available and accessible human cryopreserved hepatocytes. This allows pooling of hepatocytes from different individuals to eliminate interindividual variability.
The basic assumptions of the in vitro-in vivo clearance correlation model are 1) in vitro enzymatic kinetics is applicable to in vivo kinetic properties, 2) intrinsic clearance follows first order kinetics, and 3) metabolic clearance in the liver is the major route of clearance. These assumptions have been adopted in most studies in the search for better in vitro-in vivo correlation (Houston and Carlile, 1997; Obach, 1999). The general approach (eq. 2) for the calculation of intrinsic clearance was from the ratio of the initial amount of the test compound added to the incubation medium to the corresponding AUC0-∞. AUC0-∞ was calculated using the linear trapezoidal rule and extrapolation to time infinity by using the last few data points in the terminal phase of the log concentration-time curve. This approach requires longer incubation times (up to 72 h) to reach the terminal phase followed by linear regression to extrapolate to infinity time. Obviously over prolonged incubation times, cell viability and consequent nonconstant clearance are important concerns.
In vitro intrinsic clearance can also be obtained from the Michaelis-Menten constant (Km) and maximum velocity of the metabolic reaction (Vmax) parameters for the initial rate of metabolite formation or parent drug loss. Under the first order condition, CLint can be defined asVmax/(Km+ S). Only when S ≪Km does the in vitro intrinsic clearance correspond toVmax/Km. A separate method to derive in vitro clearance values from in vitro half-lives has also been reported (Obach et al., 1997b; Lavé et al., 1999). The in vitro half-life (t1/2) is determined from the rate of the disappearance of the parent compound. The half-life value is then converted to the corresponding intrinsic clearance using the Michaelis-Menten equation (k = 0.693/t1/2, CL = kV). The above-mentioned two methods require linear regression analysis of the raw data for the estimation of CLint. In addition, half-lives are difficult to estimate when compounds have very low intrinsic clearance. In the present study, neither approach was used because both methods involved some extrapolation that may alter the outcome of the correlation.
In the present study, we have implemented a short incubation procedure of just 2 h and developed a mathematical formula (eq. 1) for obtaining intrinsic clearance directly from experimental data with no extrapolations. The potential validity of this new approach has been tested by comparing the intrinsic clearance and in vivo clearance using the hepatocyte system. The good correlations obtained in rats, dogs, and humans suggest that this model represents an accurate approach for predicting in vivo clearance. In a side-by-side comparison, the use of the general approach (eq. 2) yielded anR2 = 0.72 (Schneider et al., 1999) versus R2 = 0.87 obtained in this study using human hepatocytes.
Several recent studies incorporated protein binding to improve the in vitro-in vivo correlation. However, protein binding is significant only for highly protein-bound (>90%) and low-clearance drugs (Oravcová et al., 1996). Because the purpose of this study was to screen for high-clearance drugs, this factor may not be important to early discovery phase. Obach (1997a) also indicated that excluding plasma protein binding resulted in better predictions of clearance from in vitro microsomal data. Data from our own laboratory (data not shown) indicated that compounds highly bound to plasma protein are usually highly bound to the liver microsomes. Therefore, in this study the effect of protein binding was not taken into account.
Although the results demonstrated the predictive potential of intrinsic clearance for rats, dogs, and humans, the approach showed some limitation for predicting clearance in monkey. The correlation for monkey was better for cryopreserved (R2 = 0.668) than for fresh (R2 = 0.450) or sandwich-cultured (R2 = 0.349) hepatocytes. These observations might result from the following reasons: 1) higher interindividual variability occurs in monkeys than other animal species; and 2) more variation in age and size of animal. Fresh and sandwich-cultured hepatocytes are only available from one individual in each incubation. It is possible that high interindividual variability resulted in the lack of correlation when fresh and sandwich-cultured hepatocytes are used. Therefore, it is important to be able to use hepatocytes that are pooled from different individuals.
The compounds for rat, dog, and monkey studies were randomly chosen from nine different programs at Schering-Plough with multiple chemotypes and vast structural differences. Yet good correlations were obtained across these multiple chemotypes, suggesting that the method presented could be applied to other chemotypes and structures. Because the in-house compounds were in early discovery phase, only the general pharmacokinetic properties were determined. In some special cases, the metabolic fate of the lead compounds was determined. Based on these limited metabolic studies, it seems that most of the in-house compounds are eliminated by phase I reaction followed by phase II reaction. In addition, the compounds chosen have reasonable Caco-2 permeability, suggesting that they are not transport limited.
The accuracy of in vitro clearance values from rat, dog, monkey, and human hepatocytes versus actual in vivo clearance values were shown in Fig. 5. Although the correlations for the low-clearance compounds are not as good as for the high-clearance compounds, all predicted clearance values are within 2-fold error for rat and dog. For the monkey, there are a few compounds that the predicted values are more than 2-fold error due to the poor correlation. Likewise, there are also a few compounds in the human study that showed high percentage error. Although the in vitro data overestimated the in vivo clearance of those few compounds, the predicted data still suggested that those compounds were low-clearance compounds.
The commercial compounds used in the human hepatocyte studies were divided into two groups. Group I includes the 16 compounds that are eliminated by phase I reaction and group 2 included 10 compounds by phase II or phase I and II. Separate linear regression analysis showed that group I and group II are very similar in the slopes of the correlation curves, and the R2 values were 0.96 and 0.82, respectively. This analysis suggests that the predictive ability apply to both phase I and phase II compounds. The cytochromes P450 involved in the elimination of those compounds include the following major isoforms: 1A2, 2C9, 2C19, 2D6, 2E2, and 3A4. After carefully examined each data point in Fig. 4, no particular outlier that is eliminated by any particular isoform of cytochrome P-450 was observed. Indicating that none of the major cytochromes P450 is deficient in the human hepatocyte. Hence, the knowledge of metabolic pathways or the cytochrome P450 isoforms involved is not crucial to the predictability of the modeling approaches put forth in this article.
In general, cryopreserved hepatocytes from rat or dog have similar predictive power as the fresh and sandwich gel hepatocytes. Cryopreserved monkey hepatocytes exhibit greater predictive power than fresh or sandwich hepatocytes. Cryopreserved hepatocytes are more readily available particularly in the case of human hepatocytes. Making experimental planning using fresh human hepatocytes ahead of time is not possible. In addition, pooling of fresh hepatocytes was not possible due to the limited availability of fresh liver from more than one individual at one time. Therefore, cryopreserved hepatocytes were our top choice among the three different types. In vitro clearance data obtained using the hepatocyte models presented in this study can only predict in vivo hepatic clearance. It will not be predictive on compounds that are cleared mainly through renal and other extrahepatic clearance.
In conclusion, the equation used in this study for calculating in vitro intrinsic clearance provides a more direct estimation than previously published methods that relied on extrapolation and linear regression analysis. In addition, the extended incubation time that was required for the previously established method is no longer needed in the method presented herein. Therefore, our approach eliminates the necessity for keeping the hepatocytes functionally active for long periods of time. This is especially important because commercially available cryopreserved human hepatocytes do not retain viability for very long under routine tissue culture conditions. The data presented herein indicate that hepatocytes processed and preserved by different methods would generate equally satisfactory correlations with in vivo data. This study provides a basis for wider use of human cryopreserved hepatocyte preparations. Pooled hepatocytes derived from several donors can help eliminate individual variation. In addition, this study presents a method that can rapidly evaluate in vitro intrinsic clearance. Satisfactory correlation between in vitro and in vivo clearance values validates the usefulness of the method in predicting in vivo human clearance. This method presents a way to substantially increase the drug-screening efficiency and provide quantitative information on drug metabolism before pharmacokinetic studies in vivo.
Acknowledgments
We thank Drs. Kathleen Cox, Anther Keung, and Diana Montgomery for critical review of this manuscript.
Footnotes
- Abbreviations used are::
- AUC
- area under the curve
- HPLC
- high-performance liquid chromatography
- MS
- mass spectrometry
- Received February 20, 2002.
- Accepted August 20, 2002.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}