


Abstract
CYP3A4, a cytochrome P450 (P450) isoform metabolizes estrogens, whereas CYP3A7, a fetal liver P450 isoform, is involved in estriol biosynthesis. The goal of this study was to evaluate expression of these enzymes in human uterine tissue during the proliferative and secretory phases of the menstrual cycle. Endometrium and cervix specimens were collected from women undergoing hysterectomy (n = 36). Total mRNA was extracted, quantified, and reverse-transcription polymerase chain reaction (RT-PCR) was carried out using consensus primers for CYP3A. The 453 base pairs PCR product was hybridized with specific internal oligonucleotide probes for CYP3A4 or CYP3A7 end labeled with 32P γ-ATP. The relative intensity of hybridization was determined by autoradiography. Expression of CYP3A7 in endometrium was significantly greater (∼10-fold) in the proliferative phase compared with the secretory phase (p < 0.05). CYP3A4 expression was comparable between the two phases. Expression of both enzymes was minimal in the cervix. Fluorescence in situ hybridization of paraffinized sections indicated localized expression of CYP3A enzymes in the glandular epithelium as well as the stroma. Comparison of relative fluorescence intensity indicated differential expression of CYP3A7 in various phases of the menstrual cycle. These results suggest that CYP3A expression in the endometrium of premenopausal women may vary depending on the menstrual cycle phase.
The P4501 enzymes are a family of hemeproteins that metabolize a number of exogenous and endogenous substrates including steroid hormones (Nelson et al., 1993; Sarkar et al., 1997). Of these, the CYP3A subfamily of enzymes has been established as the most abundant P450 in humans, representing about 30% of all hepatic P450 (Slaughter et al., 1995). So far three major isoforms have been identified in humans, CYP3A4, CYP3A5, and CYP3A7, of which CYP3A4 is the predominant hepatic isoform (Sarkar et al., 1997) and CYP3A7 is a human fetal liver form, first isolated as a 16-α-dehydroepiandrosterone-sulfate (DHEA-s) hydroxylase (Kitada et al., 1987). The role of several P450 isoforms in catalyzing the hydroxylation of cortisol, testosterone, androstenedione, estradiol, and progesterone has been established (Berliner et al., 1956; Lipman et al., 1962; Tukey et al., 1985; Guengerich, 1988). However, it is not clear whether these reactions in the liver have any physiological importance, if any, or simply serve as accessory elimination pathways. Recent evidence from our lab has shown that the uterus, specifically endometrium and cervix, expressed differential levels of CYP1A1 and CYP1B1 mRNA (Satya et al., 1998). These observations suggest that extrahepatic P450 may participate in tissue-specific biotransformations, which might be of physiologic relevance.
While several P450 isoforms have been indicated in estrogen biotransformation, CYP3A4 is probably one of the major isoforms involved in several pathways (Aoyama et al., 1990; Brian et al., 1990). CYP3A7 has been found to catalyze 16-α-hydroxylation of DHEA sulfate (Kitada et al., 1987), which is believed to be a precursor step in estrogen biosynthesis (Pritchard et al., 1992). Also in high concentrations, DHEA-s inhibits cell proliferation (Herrington et al., 1990), interferes with cellular respiration (Mohan et al., 1989), promotes cervical ripening (Sakyo et al., 1987), and inhibits placental progesterone biosynthesis (Powell et al., 1986). Therefore, CYP3A7 might be indirectly involved in several physiologically relevant processes in the uterus.
Menses is generally regarded as ischemic necrosis of a functional layer caused by contraction of spiral arteries depending on sex hormone concentrations. The endometrium undergoes its cycle of proliferation, differentiation, and desquamation, based on levels of either estrogen alone or of both estrogen and progesterone. Since the endometrium undergoes a constant dynamic growth phase, and cyclic variability of other enzyme expressions has been noted in literature (Otsuki et al., 1994) it is possible that P450 might also undergo similar cyclic variability. The goal of this research was to study the expression of CYP3A4 and CYP3A7 in the endometrium and cervix. The effect of the menstrual phase and possible cyclic variability on expression of these isoforms was also studied. The regional distribution of CYP3A7 enzyme expression was examined using fluorescence in situ hybridization.
Materials and Methods
Tissue Procurement.
Uterine tissue was obtained from women (n = 36) undergoing hysterectomy for benign indications. The protocol was approved by the West Virginia University Institutional Review Board for the Protection of Human Research Subjects. The tissues were collected within 30 min of surgery from two different regions of the uterus and carefully sectioned to maintain sampling consistency. Endometrium was obtained by scraping about 200 to 500 mg of tissue from the posterior surface of the inner lining of the uterine fundus. A thin layer of tissue was excised from the cervical canal at the 6 o'clock position, to include the endocervix and squamocolumnar junction. Tissues were collected in separate containers and stored at −70°C.
Endometrial Dating.
Endometrial dating was accomplished by a pathologist, using a light microscope, according to the technique of Hertig and classified as either proliferative (n = 20) or secretory (n = 14).
Patient Demographics.
Patients ranged in age from 25 to 48 years and in weight from 112 to 277 pounds. Patient demographics are described in Table1. None of the patients were taking medications known to induce P450 activity. One of the patients (identification number E19) was diagnosed with squamous cell carcinoma of the cervix, but still included in the analysis since samples were obtained from regions of the cervix, which appeared to be normal.
Patient demographics
RNA Extraction.
Total RNA was extracted from the tissues by homogenization in the presence of liquid nitrogen and placing it in 3 ml of RNAzol (Tel-Test Inc., Friendswood, TX). A standard extraction protocol included with the RNAzol kit was used. Typical yields were 50 to 100 μg RNA from 50 to 300 mg of tissue. RNA concentrations were measured at 260 nm on a Beckman UV spectrophotometer (Beckman Coulter, Inc., Fullerton, CA) and the Abs260/280 was >1.5 for all the samples.
Reverse-Transcription Polymerse Chain Reaction (RT-PCR) of P450 mRNAs.
The RT-PCR reaction was carried out using consensus primers for CYP3A. The target cDNA was synthesized from total RNA by RT-PCR reaction catalyzed by Superscript II Rnase H− reverse transcriptase enzyme (Invitrogen, Carlsbad, CA) and carried out in a PerkinElmer, Thermal Cycler system 2400 (PerkinElmer Life Sciences, Boston, MA). First strand cDNA was synthesized in a total volume of 20 μl of reaction mixture containing 2 μg of total RNA in the reverse transcription reaction buffer (Invitrogen), 10 mM dithiothreitol, 25 μg/ml of oligo(dT), 0.5 mM of each dNTP, 25 mM MgCl2, and 200 units of Moloney murine leukemia virus reverse transcriptase (Invitrogen). Samples were incubated at 42°C for 50 min, and the reaction was terminated by heating at 95°C for 5 min., followed by quick chilling on ice. The reaction samples were further incubated with 100 units/ml of RnaseH for 20 min at 37°C. Target sequences were amplified by hot start PCR at 94°C in a 20-μl reaction mixture containing 2 μM of each primer and 1 unit ofTaq polymerase (Promega, Madison, WI)). The sample tubes (0.2 ml) were preincubated at 94°C for 1 min before addition ofTaq polymerase and followed by 40 cycles of amplification using a thermal cycle program consisting of 94°C for 20 s of denaturation, 54°C for 20 s annealing, and 72°C for 1 min of extension. The PCR reaction products (10 μl) were separated by 2% agarose gel electrophoresis and visualized by ethidium bromide staining. Sizes of the PCR products were estimated by comparison to the migration of the DNA size markers (2000 kilobase ladder; Bio-Rad, Hercules, CA) run concurrently.
Control RNA [891-bp chloramphenicol acetyltransferase derived from previous experiments in the lab] was included to verify the PCR reaction. RNA quality was examined by electrophoresis of 5 μg of RNA in 1.2% denaturing agarose gel (formaldehyde gel) and also evaluated through the expression of β-actin gene. The primers for β-actin were obtained from BD Biosciences Clontech (Palo Alto, CA). β-Actin was positive for all tissue samples. Contamination with genomic DNA was ruled out from the negative results obtained when the RT-PCR was performed without the reverse transcriptase. Direct sequencing of PCR products did confirm the target sequences with 99% accuracy except for a 1% base pair mismatch.
Primers
The sequence for the forward and reverse PCR primers were as follows:
CYP3A forward primers, 619–636, 5′-GCC TAC AGC ATG GAT GT-3′
Reverse primers, 1055–1072, 5′-TGG ACA TCA GGG TGA GT-3′
The probes used for hybridization were as follows:
CYP3A4, 759–779 5′-CTT AAA AAA TTT GTA ACT TC-3′
CYP3A7, 642–662 5′-CGA ATG GAT CTA ATG GAT TA-3′.
The probes were obtained from Invitrogen and end labeled at the 5′ end with γ 32-P dATP (Redivue; Amersham Biosciences Inc., Piscataway, NJ) using the 5′-End labeling system (Promega). The forward reaction was carried out on 10 pmol of oligo probes obtained (Invitrogen) with T4 polynucleotide kinase at 37°C for 30 min. Unincorporated radionucleotides were separated by using G-50 Sephadex columns (Roche Diagnostics, Indianapolis, IN). The specific activity obtained for the probes was 137,000 to 156,000 cpm per 10 pmol.
Hybridization Analysis.
After electrophoresis, the gels were denatured and blotted to Zeta probe Nylon membrane (Bio-Rad), transfer was done by alkaline transfer method using 0.4 N sodium hydroxide solution. Membranes were hybridized with the specific internal probes for CYP3A3/4 and CYP3A7. After initial prehybridization with hybridization buffer at 65°C for 2 to 4 h, the membranes were incubated overnight with probes (25 μl of final volume) in 8 ml of hybridization buffer. Repeated washes were done with buffer I (5 × SSC, and 0.1% SDS) for 15 min at 34°C, followed by buffer II (25 mM NaHPO4, 1 mM EDTA, 0.1% SDS) at 29.5°C for 15 min each. A final wash was done with buffer II at room temp for 15 min. The probes hybridized with PCR fragments were visualized on a PhosphorImager (Amersham Biosciences Inc.) screen, after overnight exposure of the membranes.
ImageQuant Analysis.
The relative abundance of hybridization was measured using ImageQuant software (Amersham Biosciences) under uniform parameters of equal area, pixels, color intensity values etc. and after correcting for background intensity with relative areas in the same lane of fragment movement in respective gels. Relative mRNA levels were determined as a ratio of CYP3A expression with respect to the internal housekeeping gene β-actin. The values generated were statistically analyzed using JMP software (SAS Institute, Cary, NC).
Fluorescence in Situ Hybridization.
Paraffin sections were dewaxed in xylene and hydrated through graded ethanol concentrations to PBS. Permeabilization was carried out for 2 min in 0.01% Triton X-100 in PBS. Sections were then treated with proteinase K (1 μg/ml in 20 mM Tris-HCL, 5 mM EDTA) for 30 min at 37°C. The reaction was stopped by postfixation in buffered 4% paraformaldehyde for 15 min, followed by rinsing in PBS for 5 min. Sections were then acetylated with 0.25% acetic anhydride in 0.1 M triethanolamine for 10 min, dehydrated through graded ethanol solutions, and air dried.
Hybridization was carried out in a buffer containing 50% formamide, 4× SSC, 1× Denhardt's solution, 10% dextran sulfate, 0.24 μg/μl yeast RNA, 0.5 μg/μl salmon sperm DNA, 1% sarcosyl, 2.4 mg/ml Na2HPO4, and 10 pmol/ml biotinylated oligonucleotide probe (CYP3A4, 71–91 bp, 5′-CCT TTC AGC TCT GTG TTG CTC-3′ and CYP3A7, 1930–1949 bp, 5′-GCT TAA TAT AAA GCT TAC TA-3′). Prior to hybridization 0.1 M dithiothreitol was added to the hybridization buffer. Hybridization buffer (500 μl) was added to the coverglass chambers containing the cells and incubated in humidified chambers at 37°C overnight. After three washes with 5× SSC at room temperature, nonspecific sites were blocked with 500 μl of blocking solution (PBS containing 5% gelatin, 1% normal goat serum, and 0.1% Triton X-100). Positive hybridization was detected by incubation with rhodamine-conjugated streptavidin (1:200 dilution in 1% goat serum in PBS). Excess rhodamine conjugate was removed by three washes with PBS and the coverglass with the adhered cells was mounted on slides in n-propyl gallate. Affinity labeling was visualized using the Meridian Instruments OPTIMA scanning (Okemos, MI) confocal fluorescence microscope with 516 nm laser excitation and dual wavelength detection of emission. Control hybridizations were carried out using sense oligonucleotides as well as without any probe to estimate background fluorescence and nonspecific binding.
Data Analysis.
Statistical analyses were carried out using JMP Software (SAS Institute). Comparisons were made using a multi-factor analysis of variance. The differences in optical density of the PCR products obtained by computerized image analysis were evaluated between different regions of the uterus with respect to menstrual phase.
Results
The patient demographics are shown in Table 1. Patient age ranged from 25 to 48 years, and weight ranged from 112 to 277 pounds. None of the patients were on any comedications that could affect P450 levels. The patient charts were specifically reviewed for any drugs that could likely induce CYP3A isoforms (e.g., rifampin or rifabutin). The endometrium was classified as either being in the (a) secretory or luteal phase (n = 14) or (b) proliferative or follicular phase (n = 20). Two patients excluded from the analysis and not shown in the table had no measurable CYP3A consensus primer RT-PCR product and thus could not be processed for hybridization.
Expression of CYP3A4 and CYP3A7 in different regions of the uterus from the same patients is shown in Fig. 1, A and B. The mRNA for both enzymes was lower in the cervix (Fig. 1B) compared with the endometrium (Fig. 1A). Transcripts encoding for CYP3A7 seem to be greater in the endometrium obtained from patients in the proliferative phase compared with the secretory phase (Fig. 1B). These observations were further confirmed for other patients as well, wherein higher expression of CYP3A7 in proliferative phase (Fig.2A) was observed compared with the secretory phase (Fig. 2B). The CYP3A4 mRNA seemed to be comparable between the two phases. A semiquantitative analysis of the autoradiography by the ImageQuant software yielded relative intensities, which were compared.
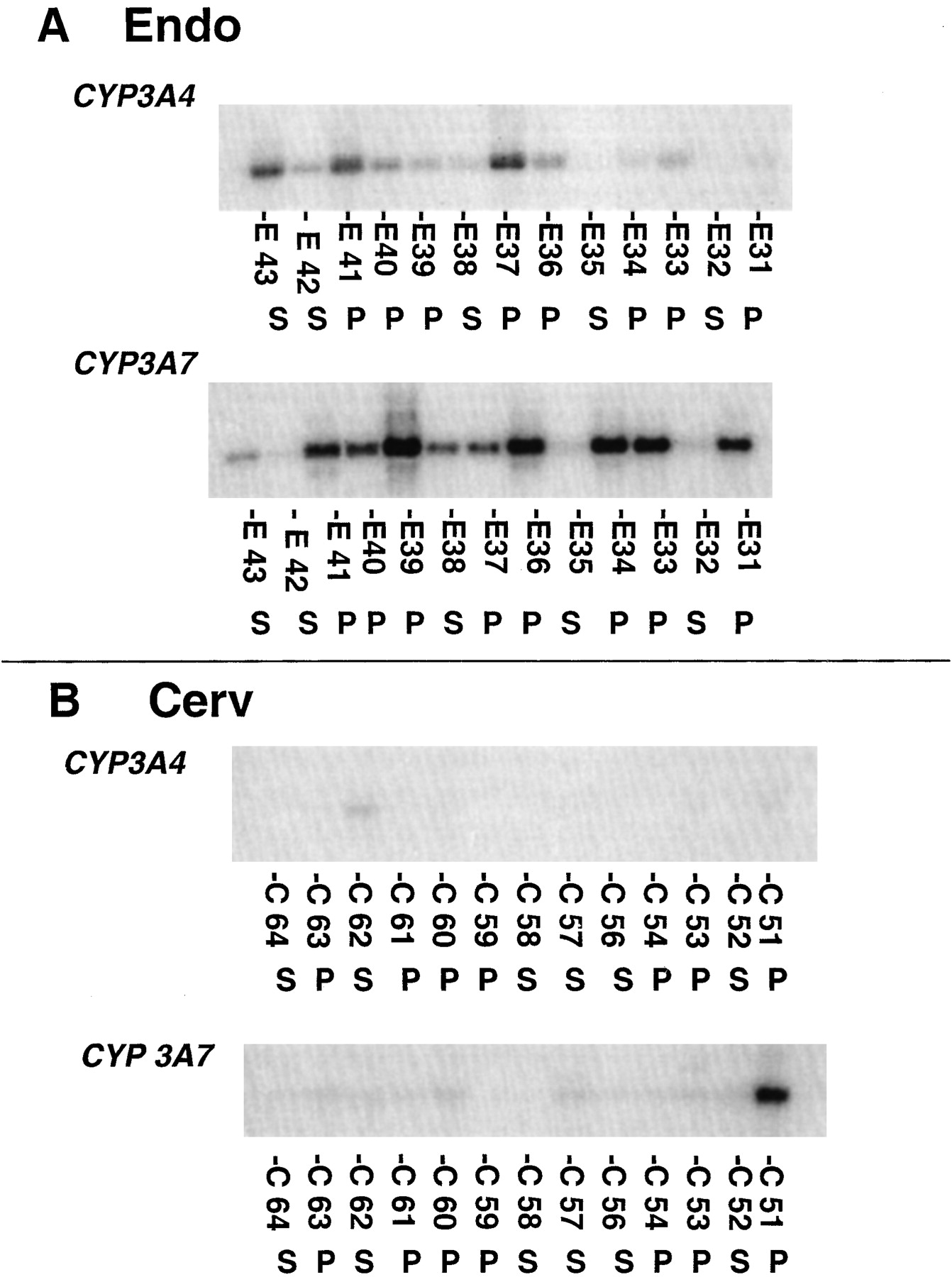
Expression of CYP3A mRNA in human (A) endometrium (Endo) and (B) cervix (Cerv) obtained from women in either proliferative (P) or secretory (S) phase of the menstrual cycle.
CYP3A mRNA expression was determined by RT-PCR using a consensus primer as described under Materials and Methods and the CYP3A4 and CYP3A7 mRNA generated by hybridization with isoform-specific oligonucleotide probes. The patient demographics, clinical diagnosis, and comedication information are provided in Table 1.

Expression of CYP3A mRNA in human endometrium tissue obtained from women in the (A) secretory and (B) proliferative phase of the menstrual cycle.
The RT-PCR products were generated using the CYP3A consensus primer as described under Materials and Methods and hybridized with CYP3A4 and CYP3A7 isoform-specific oligonucleotide probes. The patient demographics, clinical diagnosis, and comedication information are provided in Table 1.
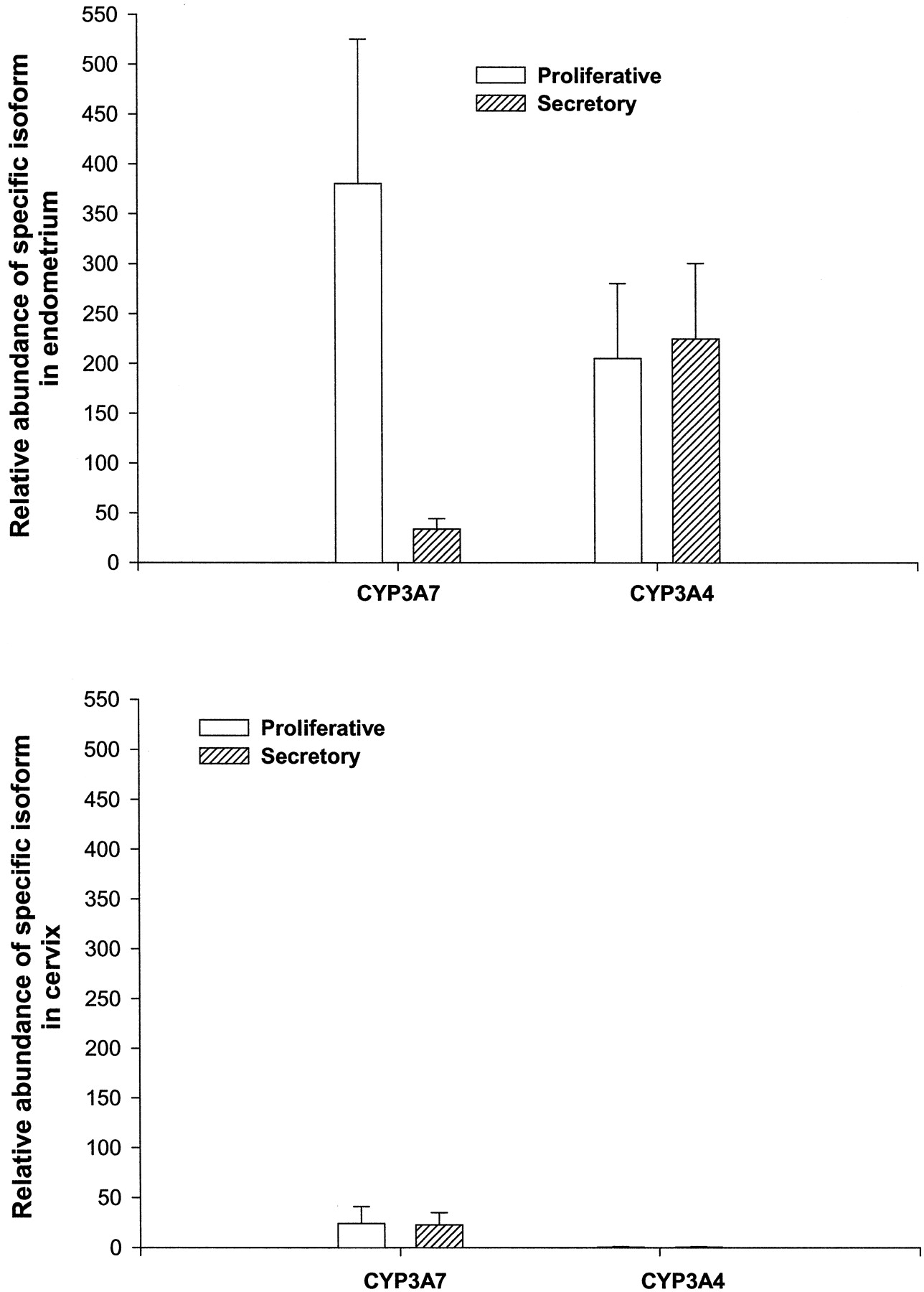
The expression of CYP3A4 and CYP3A7 as measured by the relative abundance of hybridization was considerably higher in the endometrium than in the cervix (Fig. 3, A and B). The endometrium from patients in proliferative phase had highest CYP3A7 expression levels (Fig. 3A), which were significantly greater than the secretory phase (p < 0.05). The relative intensity of hybridization for CYP3A4 in the endometrium was measurably lower as well as similar during the two menstrual cycle phases (Fig.3B).

The relative expression of CYP3A4 and CYP3A7 mRNA in (A) endometrium and (B) cervix in uterine tissue obtained from women in the proliferative phase (n = 20) or secretory phase (n = 14) of the menstrual cycle.
The results are expressed as (mean ± standard error) relative abundance of hybridization with respect to β-actin using an ImageQuant software under uniform parameters as described underMaterials and Methods.
Cyclic expression of CYP3A4 and CYP3A7 in the endometrium was further observed by hybridizing oligonucleotide probes specific for CYP3A4 and CYP3A7 mRNA on paraffin embedded, formalin-fixed sections, from women in the secretory (n = 3) and proliferative phase (n = 3). A representative sample is shown in Fig.4 where significantly higher fluorescence intensity is observed when hybridized with a CYP3A7 probe compared with the control that was hybridized with a sense oligonuculeotide. The fluorescence intensity is also significantly greater in the proliferative endometrium compared with the secretory tissue. The increased fluorescence suggests that greater amounts of biotinylated oligonucleotide probe were available to bind with the rhodamine-conjugated streptavidin. The relative abundance of fluoresence intensity seems to suggest that CYP3A7 expression is localized within the glandular epithelium as well as to some extent in the stroma surrounding the glandular structures. There was evidence of reduced staining in the endometrium from the patients who were in the secretory phase relative to the sections from endometrium from the proliferative phase. Data for CYP3A4 was not included since there were no differences between the proliferative and secretory endometrium.
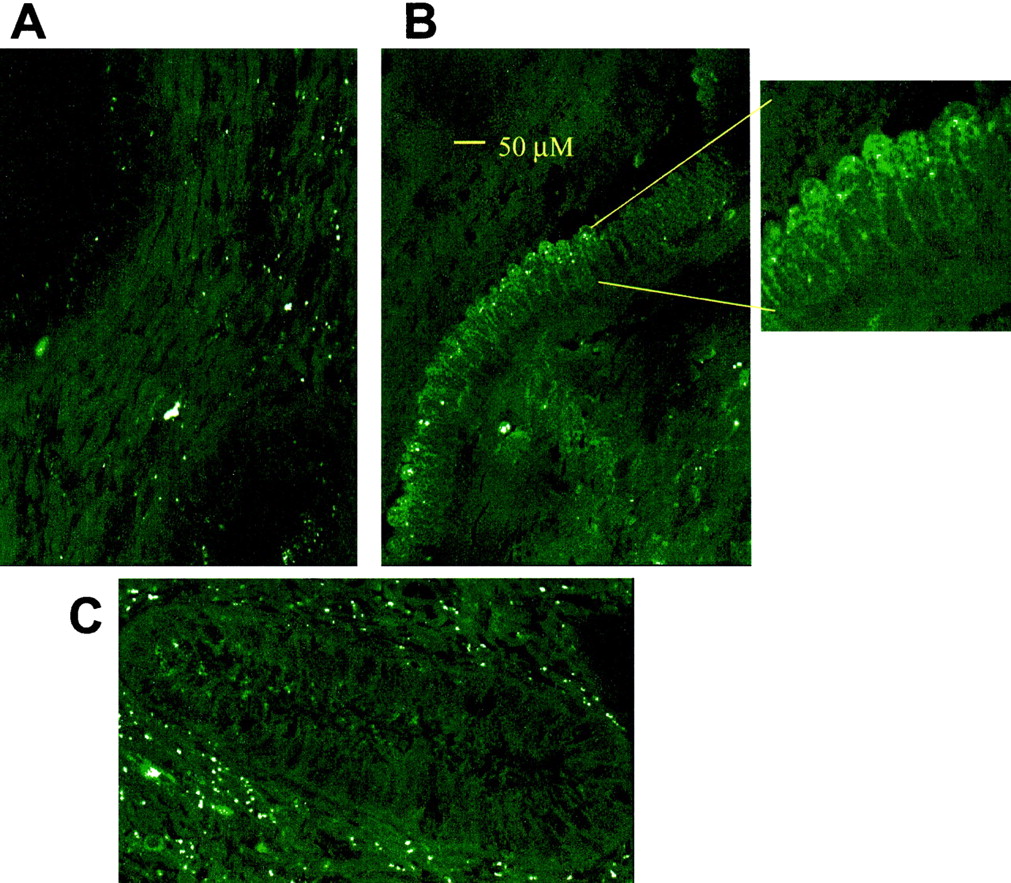
Fluorescence in situ hybridization of CYP3A7 mRNA, control slide hybridized with a sense oligonucleotide sequence (A), proliferative endometrium (B), and secretory endometrium treated with probe (C) all carried through the hybridization process.
Paraffin embedded formalin-fixed sections of human endometrium obtained from two representative patients from the proliferative and secretory phase of the menstrual cycle were hybridized with CYP3A7 mRNA specific probes. Rhodamine conjugated to streptavidin was visualized using the Meridian Instruments OPTIMA scanning confocal fluorescence microscope with 516 nm laser excitation and dual wavelength detection of emission.
Discussion
This study has demonstrated that human endometrium as well as cervix from women undergoing hysterectomy for benign conditions expresses CYP3A4 and CYP3A7. Our observations support a previous report of CYP3A7 mRNA in endometrium from a small number (n = 5) of pregnant women (Schuetz et al., 1993). The appearance of CYP3A isoforms is documented by RT-PCR sequence analysis as well as fluorescence in situ hybridization techniques. It appears that the cervix expresses minimal CYP3A isoforms; however, significant expression is seen in the endometrium that is highly variable. This variability could possibly be attributed to cyclic expression of one of these enzymes. Particularly, the CYP3A7 mRNA seems to be greater in the proliferative phase compared with the secretory phase. Identification of CYP3A4 and CYP3A7 mRNA expression in human endometrium suggests that it may be an important site for intermediary steroid metabolism. Human endometrial proteins are known to undergo cyclic changes in expression during the normal menstrual cycle (e.g., tubulin, keratin, β-galactoside binding lectin etc.) are maximally synthesized in the proliferative phase endometrium (Byrjalsen et al., 1995).
The CYP3A family is known to be inducible, for example CYP3A mRNA increases in response to administration of synthetic and naturally occurring glucocorticoids (Scheutz et al., 1984; Molowa et al., 1986), macrolide antiobiotics (Watkins et al., 1986), rifampin and rifabutin (Oesch et al., 1996), and environmental agents such as polychlorinated biphenyls and organochlorine pesticides (Scheutz et al., 1986). We observed an increase in CYP3A7 mRNA in the endometrium associated with the proliferative phase. The possibility of enzyme induction cannot be ruled out, particularly by exposure to polychlorinated biphenyls, dichlorodiphenyltrichlorethane and other persistent lipophilic environmental agents that are virtually ubiquitous in the general US population (Stehr-Green et al., 1986). However, the increase in CYP3A7 mRNA occurred consistently in the proliferative phase in most patients, suggesting that modification of endogenous substrates and hormones may be affecting CYP3A expression in the endometrium. The reported 3- to 4-fold increase in endometrium tissue concentrations of DHEA-s, a known substrate for CYP3A7 (Kitada et al., 1987), in the secretory phase of premenopausal women (Bonney et al., 1984) coincides with the decreased levels of CYP3A7 observed in this study.
Steroid hormones and their receptors have been implicated to play a role in the regulation of CYP3A enzymes. For example, transcription of the rat liver CYP3A isoform, CYP3A1, is controlled by a nonclassical glucocorticoid receptor-mediated process (Burger et al., 1992). Moreover, the synthetic glucocorticoid agonist, dexamethasone, induces CYP3A7 mRNA in the human hepatoblastoma HEPG2 (Beach et al., 1992) through transcriptional activation of CYP3A7. The 11 β-hydroxysteroid dehydrogenase type II enzyme, a potent inactivator of glucocorticoids, is significantly lower during the proliferative phase compared with the secretory phase of the menstrual cycle (Smith et al., 1997), probably resulting in higher tissue levels of glucocorticoids. Moreover, endometrial mRNA levels of corticosteroid binding globulin, which binds steroid hormones and plays a role in their transportation, were significantly lower in the proliferative phase compared with the secretory phase (Misao et al., 1994). These reports suggest that higher tissue specific levels of free glucocorticoids might exist in the endometrium during the proliferative phase, accounting for up-regulation of CYP3A7 due to its susceptibility to glucocorticoid-mediated regulation. Further investigations are necessary to clearly elucidate whether the increased mRNA levels of CYP3A7 in the proliferative phase are due to up-regulation or lower mRNA levels in the secretory phase are due to down-regulation.
The fluorescence in situ hybridization studies further demonstrated the presence of CYP3A7 mRNA in the endometrium. These studies also confirmed our observations that the CYP3A7 expression was indeed higher in the proliferative phase compared with the secretory phase. Furthermore, CYP3A7 mRNA seems to be localized specifically in the glandular epithelium of the endometrium. There was a remarkably increased fluorescence in the cytosol of the glandular epithelium with some fluorescence intensity evident in the stromal region. Although CYP3A7 is considered a human fetal liver form, our observations suggest that it is also expressed in the endometrium of mature adults. CYP3A7, first isolated as a DHEA-s hydroxylase, might be playing a significant role in the maintenance of physiologic levels of steroids. The uterus is a very sensitive organ that undergoes dramatic connective tissue turnover associated with endometrial tissue breakdown and subsequent regrowth during each menstrual cycle in response to changes in steroid levels. During the proliferative phase, blood estrogen levels are at their peak, followed by a peak in progesterone levels during the secretory phase. It is possible that the cyclic variability observed in the CYP3A7 levels reflect the physiologic need to eliminate the localized-increased steroid exposure.
Footnotes
-
This work was supported by a grant from National Institutes of Health, National Cancer Institute Grant R29-CA62369-01A1.
- Abbreviations used are::
- P450
- cytochrome P450
- DHEA-s
- 16-α-dehydroepiandrosterone-sulfate
- RT-PCR
- reverse-transcription polymerase chain reaction
- bp
- base pair(s)
- SSC
- standard saline citrate
- PBS
- phosphate-buffered saline
- Received June 19, 2002.
- Accepted September 12, 2002.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}