


Abstract
UGT2B7 has been cloned and expressed previously in COS cells and HK293 cells. Two forms have been identified: one with a tyrosine and one with a histidine at position 268. UGT2B7 has been shown to catalyze NSAIDs, catechol estrogens, and morphine-3- and -6-glucuronidation. cDNAs for UGT2B7Y268 and H268 were cloned and stably expressed in HK 293 cells. Studies were designed to test each form for reactivity toward a number of opioid compounds, xenobiotics such as menthol, oxazepam, and propranolol, and androgens such as androsterone and testosterone using membrane preparations derived from HK 293 cells. Both UGT2B7Y and UGT2B7H are highly reactive with many opioids, menthol, androsterone, and (R)- and (S)-propranolol, and similar kinetic values were observed. UGT2B7Y and UGT2B7H react poorly with oxazepam and no difference in (R)- or (S)-glucuronidation rate ratios was found. Thus, UGT2B7Y and H cannot account for the variability in the plasma or urine concentrations of these glucuronides in human populations. Our data suggest that UGT2B7 is a major isoform responsible for the glucuronidation of androsterone. Neither UGT2B7Y nor H catalyzes the glucuronidation of testosterone although each catalyzes the glucuronidation of epitestosterone. UGT2B7 seems to be a major human isoform responsible for the glucuronidation of opioids of the morphinan and oripavine class and is capable of catalyzing the glucuronidation of both the 3- and 6-hydroxyl moieties on these molecules. Thus, UGT2B7 plays a major role in the conversion of morphine to morphine-6-glucuronide, the potent analgesic metabolite of morphine.
The metabolic process of glucuronide formation is known to be extremely important for the conversion of xenobiotics and endobiotics to hydrophilic metabolites. These glucuronides are excreted rapidly by the liver and kidney. UDP-glucuronosyltransferases (UGTs,1 EC 2.4.1.17) catalyze this process. UGTs are intrinsic membrane proteins of the endoplasmic reticulum and nuclear envelope and are encoded by multiple genes of at least two families (Mackenzie et al., 1997). Members of the UGT1 gene complex share common second through fifth exons with at least twelve separate exons encoding for specific proteins with unique N-terminal domains (Ritter et al., 1992). Gene products of the UGT2 family are transcribed from unique genes. At this time, over 40 individual UGT isoforms have been identified either by cloning and expression or by purification procedures from hepatic microsomes (Mackenzie et al., 1997). Among the UGTs identified for humans, UGT2B7 is a very important isoform.
In a previous report (Coffman et al., 1997) we showed that stably expressed human UGT2B7 catalyzed the glucuronidation of opioids such as morphine and buprenorphine with high efficiency. Morphine is the most important and widely used opioid analgesic in clinical medicine and its metabolism must be understood, given the influence of its rate of metabolism and the recent appreciation of the potential role of the several metabolites formed in the overall pharmacology of this agent (Christrup, 1997). Clinical studies have shown that morphine- 6-O-glucuronide is 2–3 times more effective as an analgesic than the parent compound, and it has been found to bind with high affinity to opioid receptors (Osborne et al., 1990). In contrast, morphine- 3-O-glucuronide does not bind to the opioid receptors and is devoid of analgesic effects (Oguri et al., 1987, Pasternak et al., 1987). However, morphine-3-O-glucuronide has been shown to counteract the analgesic activity of morphine and morphine-6-O-glucuronide (Smith et al., 1990).
Morphine is a phenanthrene alkaloid with a phenolic 3-hydroxyl group and an alcoholic 6-hydroxyl moiety. UGT2B7 promotes the glucuronidation of both hydroxyls with an efficiency ratio quite similar to the ratio of glucuronides found in human urine, i.e. 7 to 1; 3-OH to 6-OH (Coffman et al., 1997). In addition, codeine, the 3-methoxy derivative of morphine, is efficiently converted to the 6-O-glucuronide by UGT2B7 (Coffman et al., 1997). Codeine-6-O-glucuronide has been found to produce analgesic responses in rats and has been shown to produce less immunosuppression than codeine (Srinivasan et al., 1996). Thus UGT2B7 reactivity with these opioids leads to the production of very important and clinically relevant metabolites.
UGT2B7 has been cloned and expressed previously with a tyrosine or a histidine at amino acid 268 (Jin et al., 1993, Ritteret al., 1989). Indeed, questions concerning the substrate specificity or even the relative reactivity with certain substrates have been raised. UGT2B7Y was reported to be active toward menthol and androsterone glucuronide formation by one laboratory (Jin et al., 1993), while another laboratory has reported that UGT2B7H is inactive with these substrates (Ritter et al., 1989). More recently Patel et al. (1995a) have raised a pharmacogenetic issue for UGT2B7. They proposed that either UGT2B7Y or UGT2B7H accounts for the variability among the human population in the ratio of oxazepam (S)- to (R)-glucuronides in urine and plasma formed from (R, S)-oxazepam. In 10% of human liver microsomal preparations the Km values of oxazepam were abnormally high for the formation of the (S)-glucuronide but remained the same for the (R)-glucuronide formation. They suggest that amino acid 268, either tyrosine or histidine of UGT2B7, accounts for variability of the (S)-oxazepam glucuronide/(R)-oxazepam glucuronide ratio.
The current study addresses the important role of human UGT2B7 in the catalysis of glucuronidation of opioid compounds, agonists, partial agonists, and opioid antagonists, using stably expressed UGT2B7Y and H and optimized experimental conditions. This work shows also the role of UGT2B7 in the glucuronidation of certain androgenic steroids and the xenobiotics, menthol, propranolol, and oxazepam. UGT2B7 is not an important factor in oxazepam metabolism in humans based on its minimal activity toward this substrate, and differences were not found between the reactivities with oxazepam for the two UGT2B7 isoforms.
Materials and Methods
Chemicals.
UDP-[U-14C]GlcUA (319 mCi/mmol) was purchased from ICN Pharmaceuticals, Inc. (Irvine, CA). Morphine sulfate was acquired from Merck and Co., Inc. (Rahway, NJ). Diprenorphine, morphine-3-glucuronide, morphine-6-glucuronide, naloxone, and naltrexone were acquired from Research Biochemicals Inc. (Natick, MA).R- and S-propranolol and (+) and (-) menthol were obtained from Aldrich Chemical Co., (Milwaukee, WI). All other aglycones, L-α-phosphatidylcholine (type XVI-E from egg yolk), dithiothreitol, UDP-GlcUA, and horseradish peroxidase-conjugated rabbit anti sheep IgG were purchased from Sigma Chemical Co. (St. Louis, MO). Protein assay reagents were from Bio-Rad (Richmond, CA). Western blot reagents, SuperBlock and SuperSignal substrate, were from Pierce (Rockford, IL).
Stable Expression of Human UGT2B7Y and UGT2B7H.
The isolation and stable expression of a cDNA coding for human UGT2B7Y has been described previously by Coffman et al. (1997). The cDNA coding for UGT2B7H was obtained from the cDNA coding for UGT2B7Y inserted in Bluescript by exchanging a 5-deoxythymidylic acid for a 5-deoxycytidylic acid using a QuickChange Site-Directed Mutagenesis kit (Stratagene, LaJolla, CA). The base change resulted in a coding for a histidine at position 268 instead of a tyrosine. The UGT2B7H DNA was inserted into the vector DNA3.1 (Invitrogene, Carlsbad, CA) and expressed in HK293 cells as previously described (Coffman et al., 1997). Sequence analysis carried out by the DNA facility of the University of Iowa confirmed the mutation. Membrane preparations from HK293 cells that stably expressed human UGT2B7Y or UGT2B7H were prepared according to the method described by King et al.(1997).
Western Blot Analysis.
Western blots of total protein from membrane preparations were blocked with SuperBlock according to the manufacturer’s instructions. The primary antibody was a sheep anti-rabbit p-nitrophenol IgG as described by Green et al. (1988). The secondary antibody was horseradish peroxidase-conjugated rabbit anti-sheep IgG. After incubation with the chemiluminescent substrate, SuperSubstrate, the blot was analyzed by luminography.
Glucuronidation Assays.
Glucuronidation activity towards opioids in membrane preparations was assayed using the method described by Puig and Tephly (1986). The glucuronides of morphine-3 hydroxyl and morphine-6-hydroxyl were identified and quantified using an HPLC method as described by Coffmanet al. (1997). Analysis of glucuronidation of non-opioids were carried out and analyzed by TLC as described by Green et al. (1994), or in the case of androsterone, testosterone, and epitestosterone by the method of Matern et al. (1994). The glucuronidation of oxazepam was analyzed using the HPLC method of Vreeet al. (1991) and 14C UDP-GlcUA. The radioactivity eluted with glucuronide standards was measured. In each case, optimal pH values and linear product formation were determined and employed for analysis of rates and kinetics. The concentration of UDP-GlcUA in the assays was 2 mM. The Kmfor UDP-GlcUA was determined using 1mM naloxone.
Results
Expression of UGT2B7Y and UGT2B7H
Western blot analysis was carried out on membranes derived from HK293 cells stably expressing UGT2B7Y or H. Results in fig.1 show the 52 Kd bands of the two UGT2B7 isoforms in varying concentrations.
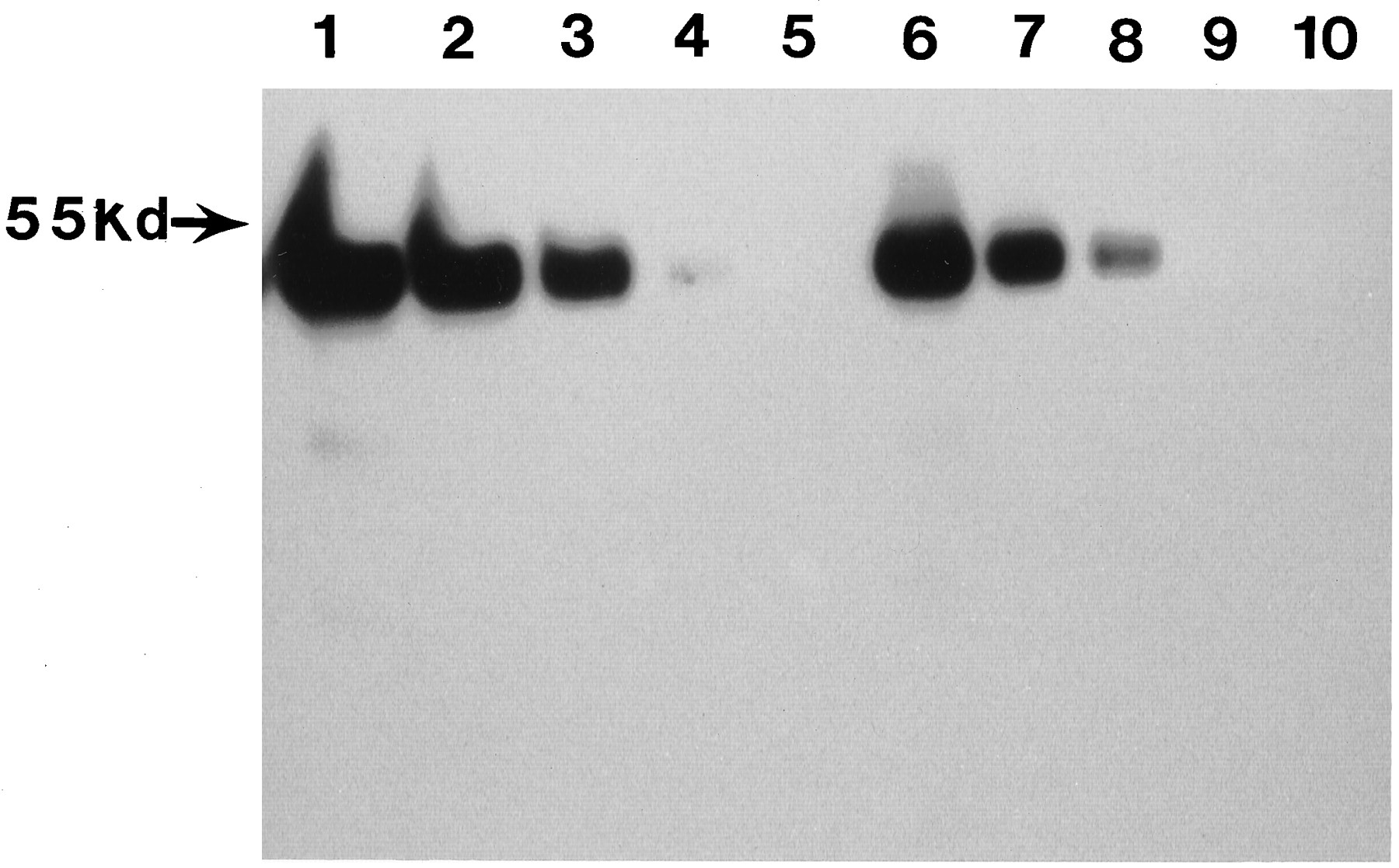
Western blot analysis of protein expression levels in membrane praparations from HK293 cells stably expressing UGT2B7H and UGT2B7Y. Western blots were analyzed as described inMethods. Lane 1–5 represent 25, 10, 5, 2, and 1μg of total protein from HK293 cell membrane preparations expressing UGT2B7H. Lanes 6–10 represent 25, 10, 5, 2, and 1μg of total protein from HK293 cell membrane preparations expressing UGT2B7Y.
Reactivity of UGT2B7Y and UGT2B7H with Opioids
Table 1 shows results obtained when membranes containing stably expressed UGT2B7Y or H were studied for activity toward 18 different opioid derivatives. As demonstrated in a previous study (Coffman et al., 1997), morphine glucuronidation rates were high and both 3- and 6-hydroxyl glucuronides were formed. No differences in rates were found between UGT2B7Y and UGT2B7H. Naloxone and nalorphine were glucuronidated with as high rates as morphine, whereas the oripavine derivatives were glucuronidated at rates nearly 10-fold lower. Studies on various other substituted morphinans are shown. The only difference between UGT2B7Y and H was that normorphine and naltriben seem not to be glucuronidated by UGT2B7H.
Glucuronidation of opioid derivatives catalyzed by expressed UGT2B7
Kinetics of UGT2B7Y and UGT2B7H with Opioid Substrates
The results of kinetic studies are shown in table2. Both UGT2B7 isoforms seem to have similar efficiencies for morphinan substrates, among them morphine, codeine, naloxone, nalorphine, and naltrexone. However, buprenorphine had a 10-fold higher efficiency with UGT2B7Y than for the H form. TheKm for UDP-GlcUA was similar for both UGT2B7 isoforms, as were efficiency values, using naloxone as aglycone.
Kinetics of glucuronidation with various opioid substrates using membrane preparations of HK293 cells stably expressing either UGT2B7Y or H using two different passages of cultured cells
Reactivity of UGT2B7Y and UGT2B7H with Oxazepam, Androgens, and Menthol
Previous studies from other laboratories have reported data that were inconsistent with respect to the reactivities of UGT2B7Y or H with menthol and androsterone (Jin et al., 1993, Ritter et al., 1989). Table 3 shows that UGT2B7Y or H react equally well with (+) or (-) menthol. Furthermore, androsterone and epitestosterone are very good substrates for both isoforms of UGT2B7. Interestingly, testosterone does not react with either isoform under the conditions employed in these experiments. Oxazepam is a poor substrate for either isoform of UGT2B7 and the ratio of (R) to (S) glucuronide is equivalent.
Glucuronidation of several non-opioid substrates catalyzed by UGT2B7
Kinetic studies of UGT2B7Y and UGT2B7H with Propranolol and Androsterone
Since oxazepam is a poor substrate with UGT2B7, another important stereoisomeric drug, propranolol, was studied. For this compound, it was possible to obtain the pure (R)- and (S)- forms. Table 4 shows that both UGT2B7Y and H react with high efficiency toward (R)- and (S)-propranolol. Also, no differences in Kmvalues and efficiencies with androsterone were observed for UGT2B7Y and H.
Kinetics of glucuronidation of R- and S-propranolol and androsterone using membrane preparations of HK293 cells stably expressing UGT2B7Y or H
Discussion
It is clear that UGT2B7 is a UGT isoenzyme of major importance for the glucuronidation in humans of a large number of clinically important opioid compounds. This UGT catalyzes glucuronidation of both the 3- and 6-hydroxyl position of morphinan derivatives, such as morphine, and is also a catalyst of codeine 6-O-glucuronide formation. No other UGT isoform has been shown to catalyze 6-Oglucuronidation, although rat and human UGT1A1 do catalyze the 3-O-glucuronidation of the opioid compounds such as morphinan derivatives, naltrexone, naloxone, and morphine, and the oripavine antagonist/partial agonist, buprenorphine (Coffman et al., 1995, King et al., 1996). UGT2B7H was first cloned and expressed transiently in COS-1 cells by Ritter et al.(1989), who found 3,4-catechol estrogens and estriol to be good substrates but reported that there was no detectable conversion of morphine to a glucuronide. Jin et al. (1989) reported on the cloning and transient expression of UGT2B7Y but this isoform, as expressed in COS-7 cells, was reported to possess no reactivity with morphine. Technique differences between those studies and the one reported here may be the reason that morphine glucuronidation was not detected previously. With respect to opioids, both UGT2B7Y and H, as stably expressed in HK293 cells, have similar reactivity with the opioids studied here. Only a few substrates were glucuronidated with some differences by the two forms of UGT2B7: normorphine, norbuprenorphine, naltriben, and buprenorphine. The efficiency of glucuronidation of buprenorphine was 10 times higher for the Y form. Normorphine and norbuprenorphine seem to possess some different reactivities because these compounds are less active as substrates for the H form than the Y form of the enzyme. Both forms catalyze glucuronidation of morphine and codeine at the 6-OH position, in contrast to rat UGT2B1, the major catalyst of opioid glucuronidation in rats, which catalyzes only 3-O-glucuronidation of opioids (King et al., 1997). For both morphinan and oripavine structures (fig. 2), it can be concluded that aliphatic substitutions at the 17-nitrogen position increase the glucuronidation rate, and that carbon chains allowed for increased efficiency more than the substitution of an alicyclic group, as was found for rat UGT2B1 (King et al., 1997). Other structural differences seem also to play a role for both forms. Hydration of the 7 and 8 positions decreases the catalytic rate 7-fold (morphine/dihydromorphine), and lack of a substituent at the 6-position (levorphanol) yields even lower rates. Also, a propene group at position 6 gives a lower rate than an oxygen substitution (nalmefene/naltrexone). Substitutions at the 14-position are likewise important, a hydroxy group increases rates (oxymorphone/hydromorphone).

Morphinan and Oripavine Structures. a. Morphinan. b. Oripavine. All morphinan aglycones were 4,5 epoxides, except for levorphanol.
The same trends are seen in the kinetic data for selected compounds. Naloxone, naltrexone, and hydromorphone are identical compounds except for the N-substitutions. LowerKm values for the proteins are favored by the aliphatic chain substitution over the cyclic, andKm values are higher where methyl group substitution is present. The presence of a 14-hydroxy group yields values of higher Km , and increases theVmax (oxymorphone/hydromorphone). The methoxy substitution on position 3 seems to influence the efficiency of glucuronidation of the 6-OH position. The efficiency of codeine glucuronidation was one tenth of that of morphine. Codeine has been reported (Yue et al., 1989) to be glucuronidated at a lower rate in the Chinese population when compared with the Caucasian population, but based on our data no differences in codeine glucuronidation between UGT2B7Y and UGT2B7H were seen. The observation of population differences is not likely to be explained by this polymorphism of UGT2B7.
Patel et al. (1995a) have proposed that the two forms of UGT2B7, H and Y, account for differences in two human populations with respect to the apparent differences in the ratio of (R)- and (S)-oxazepam glucuronides excreted in the urine when a racemic mixture of oxazepam was administered orally. We did not observe any differences for glucuronidation by UGT2B7Y and H. Activities were so low for oxazepam that kinetic values could not be obtained accurately. The low rate of reactivity of UGT2B7Y in oxazepam glucuronidation has also been shown by Jin et al. (1993). We believe, based on this data, that UGT2B7 is not an important isoform for glucuronidation of oxazepam. The proponents of the role of UGT2B7 in oxazepam metabolism did not show inhibition of oxazepam glucuronidation by morphine in human liver microsomes (Patel et al., 1995b), although oxazepam glucuronidation was inhibited by various NSAIDs. Because oxazepam was found to be a poor substrate with UGT2B7, the glucuronidations of (R)- and (S)-propanolol were investigated. Propanolol is a clinically important adrenergic β-receptor antagonist and is an excellent substrate with UGT2B7 isoforms. Neither isoform showed stereoselective glucuronidation of (R)- or (S)-propanolol, and there were no differences in Km values with (R)- and (S)-propanolol.
Other laboratories have reported on the glucuronidation of catechol estrogens and NSAIDs by UGT2B7Y or H (Jin et al., 1993,Ritter et al., 1989) but have disagreed on the reactivity of these UGT isoforms toward a few substrates such as menthol and androsterone. The glucuronidations of (+) and (−) menthol were catalyzed by both enzyme forms, and the kinetics for androsterone showed no differences between the isoforms. However, the efficiency of androsterone glucuronidation was found to be so high as to suggest that UGT2B7 is the major isoform catalyzing 3α-hydroxy steroids. This has also recently been reported by Jin et al. (1997). Both isoforms showed activity towards epitestosterone (17 α-ol), but not towards testosterone (17 β-ol).
Human UGT2B7 catalyzes the glucuronidation of a wide variety of substrates, such as catechol estrogens, androgens, opioids, NSAIDs, and propanolol. No differences were seen in the present study with either isoform for most of the compounds tested. The glucuronidation of morphine and codeine is of special interest. Morphine-6-O-glucuronide is known to be a much more potent analgesic than its parent compound in humans (Christrup, 1997, Osborneet al., 1990), and codeine-6-O-glucuronide has also been found to demonstrate high analgesic potency in rats (Srinivasan et al., 1996). Morphine-3-O-glucuronide, the major metabolite of morphine, has no analgesic effect, but has been shown to cause hyperglycemic and neuroendrocrine potentiating effects in rats (Hashiguchi et al., 1995). In addition, morphine-3-O-glucuronide antagonizes the respiratory depression induced by morphine and morphine-6-O-glucuronide (Christrup, 1997, Smith et al., 1990), which might explain why chronic users of morphine become tolerant to otherwise lethal doses of morphine. Since UGT2B7 catalyzes the glucuronidation of both morphine and codeine, the two most widely used analgesic agents of intense pain in humans, our understanding of the reactivities of this isoenzyme becomes most important in understanding the regulation of the metabolic pathway of opioid disposition.
Acknowledgment
We wish to thank Mitchell D. Green for helpful discussions in the preparation of this manuscript.
Footnotes
-
Send reprint requests to: Dr. Thomas R. Tephly, 2–452 Bowen Science Building, Department of Pharmacology, University of Iowa, Iowa City, IA 52242. E-mail: Thomas-Tephly{at}uiowa.edu.
-
This research was supported by NIH grant GM26221.
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- UGT2B7Y
- UGT2B7(Y268)
- UGT2B7H
- UGT2B7(H268)
- UDP-GlcUA
- UDP-glucuronic acid
- NSAID
- non-steroid anti-inflammatory drug
- Received July 31, 1997.
- Accepted October 20, 1997.
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}
{kind=link}