


Abstract
Afloqualone (AFQ) is one of the centrally acting muscle relaxants. AFQ N-glucuronide is the most abundant metabolite in human urine when administered orally, whereas it was not detected in the urine when administered to rats, dogs, and monkeys. Species differences in AFQ N-glucuronidation were investigated with liver microsomes obtained from humans and experimental animals. The kinetics of AFQ N-glucuronidation in human liver microsomes showed a typical Michaelis-Menten plot. The Km and Vmax values accounted for 2019 ± 85.9 μM and 871.2 ± 17.9 pmol/min/mg protein, respectively. The Vmax and intrinsic clearance (CLint) values of AFQ N-glucuronidation in human liver were approximately 4- to 10-fold and 2- to 4-fold higher than those in rat, dog, and monkey, respectively. Among 12 recombinant human UDP-glucuronosyltransferase (UGT) isoforms, both UGT1A4 and UGT1A3 exhibited high AFQ N-glucuronosyltransferase activities. The Km value of AFQ N-glucuronidation in recombinant UGT1A4 microsomes was very close to that in human liver microsomes. The formation of AFQ N-glucuronidation by human liver, jejunum, and recombinant UGT1A4 microsomes was effectively inhibited by trifluoperazine, a known specific substrate for UGT1A4. The AFQ N-glucuronidation activities in seven human liver microsomes were significantly correlated with trifluoperazine N-glucuronidation activities (r2 = 0.798, p < 0.01). In contrast, the Km value of AFQ N-glucuronidation in recombinant UGT1A3 microsomes was relatively close to that in human jejunum microsomes. These results demonstrate that AFQ N-glucuronidation in human is mainly catalyzed by UGT1A4 in the liver and by UGT1A3, as well as UGT1A4 in the intestine.
Afloqualone (AFQ; Fig. 1), 6-amino-2-fluoromethyl-3-(o-tolyl)-4-(3H)-quinazolinone, is one of the centrally acting muscle relaxants (Tani et al., 1979; Ochiai and Ishida, 1981). It has been reported that AFQ is extensively metabolized in humans (Furuuchi et al., 1983), rats (Otsuka et al., 1983b), dogs, and monkeys (Otsuka et al., 1983a), and there are species differences in the composition of urinary metabolites. The major metabolic pathways of AFQ consist of acetylation of the aromatic amino group, followed by hydroxylation at the methyl carbon of either the acetyl or 2′-methyl residue, whereas the N-acetylated metabolites have not been found in dogs. In addition, it has been reported that when AFQ is administered orally to humans, rats, dogs, and monkeys, the parent AFQ and its metabolites are excreted into urine (human, 20% of dose; experimental animals, 50-70% of dose), in which AFQ N-glucuronide (Fig. 1) is the most abundant metabolite (8% of dose) in human urine (Furuuchi et al., 1983; Otsuka et al., 1983a,b). Although this type of conjugation seems common to primary aromatic amines, the N-glucuronide of AFQ has not been detected in rats, dogs, and monkeys. The N-glucuronidation of AFQ plays an important role in the metabolism of AFQ in humans.

Structures of AFQ and its N-glucuronide.
Glucuronidation is one of the most common phase II biotransformation reactions for therapeutic drugs catalyzed by UDP-glucuronosyltransferase (UGT). To date, 18 functional UGT isoforms have been identified in humans and classified on the basis of primary amino acid sequence into two families of protein termed UGT1 and UGT2 (Miners et al., 2004). The UGTs are not only involved in the metabolism of many drugs (e.g., morphine, paracetamol) but also capable of the biotransformation of endogenous substrates (e.g., bilirubin and ethynylestradiol) and several xenobiotics (Green and Tephly, 1996; Radominska-Pandya et al., 1999). Glucuronidation of amines also represents a less common metabolic pathway among phase II conjugation reactions of a wide variety of endo- and xenobiotics. Aryl- and alkylamines, sulfonamides, and heterocyclic amines have been reported to undergo glucuronidation in vitro and in vivo in a large number of animal species and humans (Chiu and Huskey, 1998). The primary function of N-glucuronidation, similar to that of O-glucuronidation, is believed to be detoxification. Human UGTs are expressed in a tissue-specific manner in hepatic and extrahepatic tissues (Strassburg et al., 1997, 2000). The distribution of individual UGT isoforms is believed to determine tissue-specific metabolism and detoxification. Whereas, in the past, the liver had been thought to be the extraordinarily powerful organ involved in first-pass metabolism, the small intestine has become recognized as a major site of metabolism for drugs, especially when administered orally. In addition to cytochrome P450 and carboxylesterase, intestinal glucuronidation is known to play a major role in first-pass metabolism (Fisher et al., 2001). UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A9, UGT2B4, UGT2B7, UGT2B10, UGT2B11, UGT2B15, and UGT2B17 are expressed in human liver (King et al., 2000; Tukey and Strassburg, 2000; Fisher et al., 2001). UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A8, UGT1A10, UGT2B4, UGT2B7, UGT2B10, and UGT2B15 are expressed in human intestine (King et al., 2000; Tukey and Strassburg, 2000; Fisher et al., 2001). The determination of the identity of the enzymes involved in drug metabolism is important to understand variation in drug effect and metabolism. Moreover, since many drug-metabolizing enzymes (e.g., cytochrome P450 and UGT) are involved in the metabolism of several xenobiotics, the determination of the enzyme involved in the metabolism of a specific drug could provide essential information about potential drug interaction (Bertz and Granneman, 1997; Liston et al., 2001).
The purpose of the present study was, first, to clarify species differences in N-glucuronidation on the metabolism of AFQ using liver microsomes obtained from humans and experimental animals. Second, to identify the human UGT isoform(s) responsible for AFQ N-glucuronidation, the activities and kinetics in commercially available recombinant UGT microsomes from baculovirus-insect cells were determined. Furthermore, inhibition analyses, using human liver and jejunum microsomes, and correlation analyses with typical substrates for UGT isoforms were performed.
Materials and Methods
Materials. AFQ was synthesized at the Medicinal Chemistry Research Laboratories of our company (Tanabe Seiyaku Co., Ltd., Saitama, Japan). UDP-glucuronic acid, alamethicin, trifluoperazine, and genistein were purchased from Sigma-Aldrich (St. Louis, MO). Pooled human liver microsomes from donors (13 males, 4-62 years old; 9 females, 40-74 years old) and microsomes from seven individual human livers (HG32, HG74, HH18, HH47, HH91, HH74, and HK25) were purchased from BD Gentest (Woburn, MA). The glucuronosyltransferase activities of β-estradiol (for the 3-OH position), trifluoperazine, and propofol in these human liver microsomes were provided as typical activities for UGT1A1, UGT1A4, and UGT1A9, respectively, by the manufacturer. Recombinant human UGTs (UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A7, UGT1A8, UGT1A9, UGT1A10, UGT2B4, UGT2B7, UGT2B15, and UGT2B17) expressed in baculovirus-infected insect cells (Supersomes) were also obtained from BD Gentest. Pooled human jejunum microsomes from donors (five males, 19-55 years old; five females, 16-51 years old) were obtained from Tissue Transformation Technologies (Edison, NJ). For animal species comparison, pooled male rat liver microsomes (pool of 82 Sprague-Dawley rats), pooled male dog liver microsomes (pool of five beagle dogs), pooled male monkey liver microsomes (pool of six cynomolgus monkeys), and pooled male rabbit liver microsomes (pool of six New Zealand White rabbits) were purchased from BD Gentest. The protein contents were used as described in the data sheets provided by the manufacturer. All other chemicals and solvents were of the highest grade commercially available.
AFQ N-Glucuronidation Assay. A typical incubation mixture (100-μl total volume) contained 50 mM Tris-HCl buffer, pH 7.5, 8 mM MgCl2, 5 mM UDP-glucuronic acid, 25 μg/ml alamethicin, 0.5 mg/ml human liver microsomes (human jejunum microsomes, recombinant UGTs, or experimental animal liver microsomes), and 100 μM AFQ. AFQ was dissolved in dimethyl sulfoxide. The final concentration of dimethyl sulfoxide in the reaction mixture was 1% (v/v). After preincubation at 37°C for 5 min, the reaction was initiated by the addition of AFQ. The reaction mixture was incubated at 37°C for 45 min (10 min in the case of rabbit liver microsomes) and the reaction stopped with 50 μl of ice-cold acetonitrile. After removal of the protein by centrifugation at 10,000 rpm for 3 min, a 120-μl portion of the supernatant was added to 80 μl of water, and then a 150-μl portion of the sample was subjected to HPLC. Chromatography was performed using a Waters Alliance HPLC system (model 2695; Waters, Milford, MA), equipped with a model 2996 photodiode array detector (Waters), and a Symmetry C18 column (4.6 × 150 mm, 3.5 μm; Waters). The column temperature was 40°C and the eluate was monitored at 300 nm. Mobile phase A was 0.1% formic acid in water (v/v) and mobile phase B was acetonitrile. A linear gradient of 5 to 55% mobile phase B over 10 min was used for the analysis of AFQ and its glucuronide at a flow rate of 1.0 ml/min. The retention times of AFQ N-glucuronide and AFQ were 8.0 and 9.6 min, respectively. Due to the absence of authentic standard for AFQ N-glucuronide, quantification of the glucuronide in the incubation mixtures was accomplished using standard curves for AFQ. The absorbance spectrum of AFQ N-glucuronide was consistent with that of AFQ, for which the wavelength of maximum absorbance of AFQ N-glucuronide and AFQ was 300 nm. The peak area ratio of AFQ to the converted AFQ N-glucuronide in the HPLC conditions (monitored at 300 nm) was 1.0, after which a correction factor was applied to the calculation of AFQ N-glucuronide formed in the microsomal incubation mixture.
Identification of AFQ N-Glucuronide by LC-MS/MS Analysis. The incubation of AFQ with human liver microsomes for structure identification was carried out as described above except for different concentrations of microsomal protein (1.0 mg/ml) and incubation time (for 60 min). Detection of AFQ N-glucuronide was achieved with injection of 5 μl of the centrifugal supernatant onto the LC-MS/MS system. LC-MS/MS analysis was performed using a Q-TOF Ultima API (Waters) coupled to an Agilent model 1100 series HPLC system (Agilent Technologies, Palo Alto, CA). Ionization of the analytes was achieved by electrospray in the positive ion mode. A Symmetry C18 column (2.1 × 100 mm, 3.5 μm; Waters) was used for the LC separation. The HPLC separation was carried out at 45°C using a gradient composed of mobile phase A [0.1% formic acid in water (v/v)] and mobile phase B (acetonitrile). The gradient, expressed as changes in mobile phase B, was as follows: 0 to 1 min, hold at 10% B; 1 to 8 min, a linear increase from 10% to 70% B; 8 to 10 min, hold at 90% B. The mobile phase flow rate was 0.3 ml/min. MS conditions used were as follows: capillary voltage, 2.7 kV; cone voltage, 35 V; cone gas, 50 l/h; desolvation gas, 500 l/h; desolvation temperature, 450°C; source temperature, 100°C. MS and MS/MS spectra were obtained in the range of m/z 100 to 600. Collision-induced dissociation (CID) was performed using argon as the collision gas, and collision energy was 15 eV. The retention times of AFQ N-glucuronide and AFQ were 6.5 and 7.4 min, respectively.
Effects of Organic Solvents on AFQ N-Glucuronidation. The effects of methanol and dimethyl sulfoxide on the AFQ N-glucuronidation in human liver microsomes were determined. Incubations were performed as described above. Each organic solvent was added separately to incubations to give final concentrations of 0.5, 1, and 2%, v/v. Data were compared with activities determined in the presence of 0.5% (v/v) methanol, and the inhibitory effects of organic solvents were indicated as percentage of control activity for duplicate determinations.
Kinetic Analyses. The kinetics studies were performed using the liver microsomes obtained from human, rat, dog, monkey, and rabbit, and human jejunum microsomes, and recombinant UGTs (UGT1A1, UGT1A3, and UGT1A4) expressed in microsomes. In determining the kinetic parameters, the AFQ concentration ranged from 15.6 μM to 4000 μM, except for the case of rabbit liver microsomes, which ranged from 1.56 μM to 400 μM. Kinetic parameters were estimated from the fitted curves using the Prism computer program (GraphPad Software Inc., San Diego, CA), designed for nonlinear regression analysis.
The Michaelis-Menten equation, 
where V is the rate of reaction, Vmax is the maximum velocity, Km is Michaelis constant (substrate concentration at 0.5 Vmax), and [S] is the substrate concentration. The velocity versus substrate concentration can be described by Michaelis-Menten kinetics that gives hyperbolic kinetics.
However, the substrate inhibition model (Houston and Kenworthy, 2000) was selected when a hyperbolic-type curve was apparent at low concentrations and there was no clearly defined plateau at high substrate concentrations, and the velocity decreased as substrate concentrations are further increased: 
where Ksi is the constant describing the substrate inhibition interaction.
When the velocity described sigmoidal kinetics and the corresponding Eadie-Hofstee plots were curvilinear, the Hill equation (Weiss, 1997) was selected: 
where S50 is the substrate concentration resulting in 50% of Vmax (analogous to Km in previous equations) and n is the Hill coefficient.
Autoactivation is characterized by the dependence of clearance on substrate concentration, and an alternative to intrinsic clearance (CLint), calculated as Vmax/Km, is required. For sigmoidal kinetic data, maximum clearance (CLmax) provides an estimate of the highest clearance attained, that is, when the enzyme is fully activated before saturation occurs (Houston and Kenworthy, 2000): 
Inhibition Analysis of AFQ N-Glucuronosyltransferase Activity in Human Liver and Jejunum Microsomes, and Recombinant UGTs. Two compounds were tested for the inhibitory effects on the AFQ glucuronosyltransferase activity. Trifluoperazine is a known, typical substrate for UGT1A4 (Green and Tephly, 1996; Nakajima et al., 2002; Ghosal et al., 2004). Genistein is a substrate for UGT1A3 and UGT1A4 (Green et al., 1998; King et al., 2000). Trifluoperazine and genistein were dissolved in dimethyl sulfoxide, with the solvent being used as the control. The final concentration of organic solvents in the reaction mixture was 1% (v/v). The inhibitor concentrations for IC50 determinations in human liver or jejunum microsomes were 1 to 300 μM. Each incubation contained 0.5 mg/ml microsomal protein. The glucuronosyltransferase activities in human liver and jejunum microsomes, and recombinant UGT1A3 and UGT1A4 at 100 μM AFQ were determined as described above. The IC50 values were estimated graphically.
Correlation Analysis. Correlation analysis between AFQ N-glucuronidation and the other glucuronidation activities was determined by Pearson's moment method. A p value of less than 0.05 was considered statistically significant.
Results
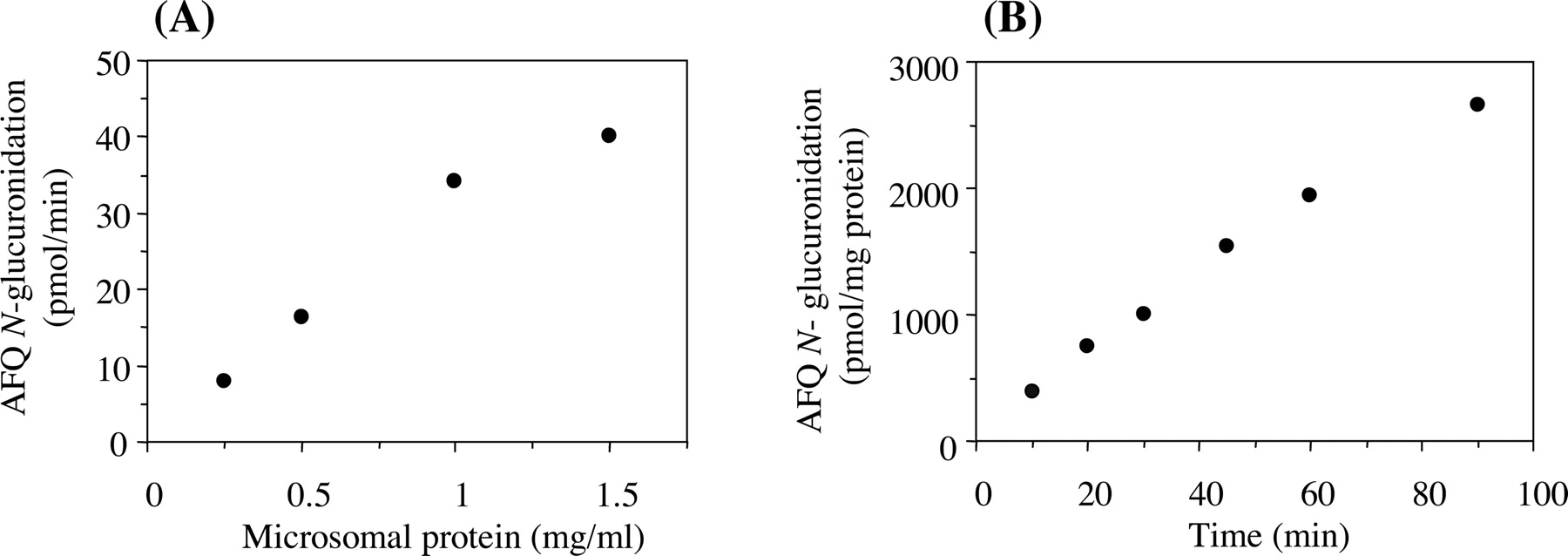
AFQ N-Glucuronidation in Human Liver Microsomes. The electrospray ionization mass spectrum of a peak typically formed by incubation of AFQ with human liver microsomes in the presence of UDP-glucuronic acid had a [M + H]+ ion at m/z 460 corresponding to AFQ N-glucuronide. The product ion spectrum of the peak showed that a loss of the glucuronic acid element (176 Da) yields the protonated aglycon ion at m/z 284. From these observations, it was confirmed that the peak formed by the incubation of AFQ with human liver microsomes and UDP-glucuronic acid was AFQ N-glucuronide. The formation was linear up to 1.0 mg/ml microsomal protein and 60 min of incubation time (Fig. 2). Methanol tested up to 2% (v/v) had no effect on the formation of AFQ N-glucuronide. Dimethyl sulfoxide (0.5, 1, and 2%, v/v) reduced the activities of AFQ N-glucuronidation in human liver microsomes by 8, 10, and 14%, respectively (data not shown). Due to the limited water solubility of AFQ, dimethyl sulfoxide was used as an organic solvent in kinetics and inhibition studies, in which the final concentration of that in the reaction mixture was 1% (v/v).
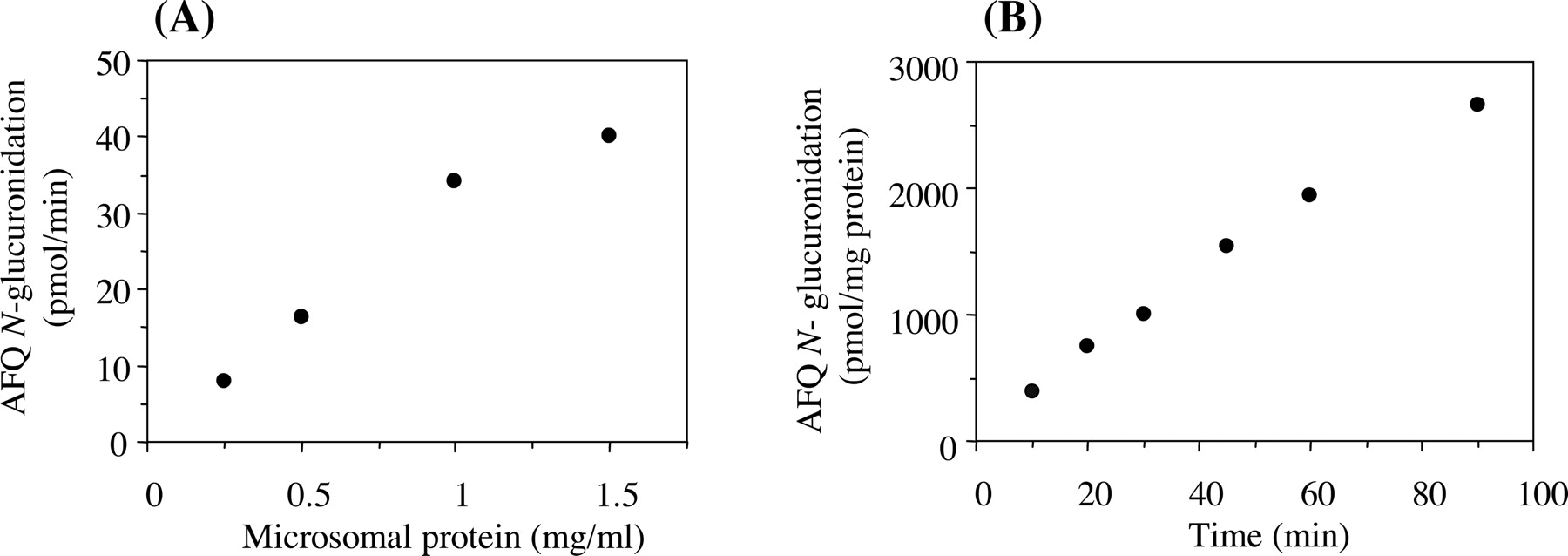
Formation of AFQ N-glucuronide as a function of the protein concentration of human liver microsomes (A) and incubation time (B). Unless specified, the standard incubation mixture contained 0.5 mg/ml microsomal protein, 100 μM AFQ, and 5 mM UDP-glucuronic acid, and was incubated at 37°C for 45 min. Each value represents the mean of duplicate determinations.
Kinetics of AFQ N-Glucuronidation in Liver Microsomes. Kinetic analysis of the AFQ N-glucuronidation was performed in liver microsomes obtained from humans and experimental animals. As shown in Fig. 3, A, B, D, and E, the AFQ N-glucuronidation by human, rat, monkey, and rabbit liver microsomes displayed typical Michaelis-Menten kinetics. However, glucuronidation by dog liver microsomes showed a sigmoidal kinetics, which manifests as a curvilinear Eadie-Hofstee plot (Fig. 3C). Fitting the data points to eq. 1 or 3 yielded the kinetic parameters listed in Table 1. Incubation of various concentrations (15.6-4000 μM for human, rat, dog, and monkey; 1.56-400 μM for rabbit) of AFQ with human, rat, monkey, and rabbit liver microsomes demonstrated that the Km values were 2019 ± 85.9, 611.0 ± 26.3, 732.5 ± 47.2, and 24.8 ± 0.7 μM, respectively. Corresponding Vmax values were 871.2 ± 17.9, 89.2 ± 1.3, 90.1 ± 2.1, and 9375 ± 68.9 pmol/min/mg protein, respectively. Corresponding CLint (Vmax/Km) values were estimated to be 0.432, 0.146, 0.123, and 378.2 μl/min/mg protein, respectively. With respect to dog liver microsomes, the S50 (apparent Km) and Vmax values were 582.1 ± 13.2 μM and 236.9 ± 2.4 pmol/min/mg protein, respectively. The CLmax value was estimated to be 0.219 μl/min/mg protein.
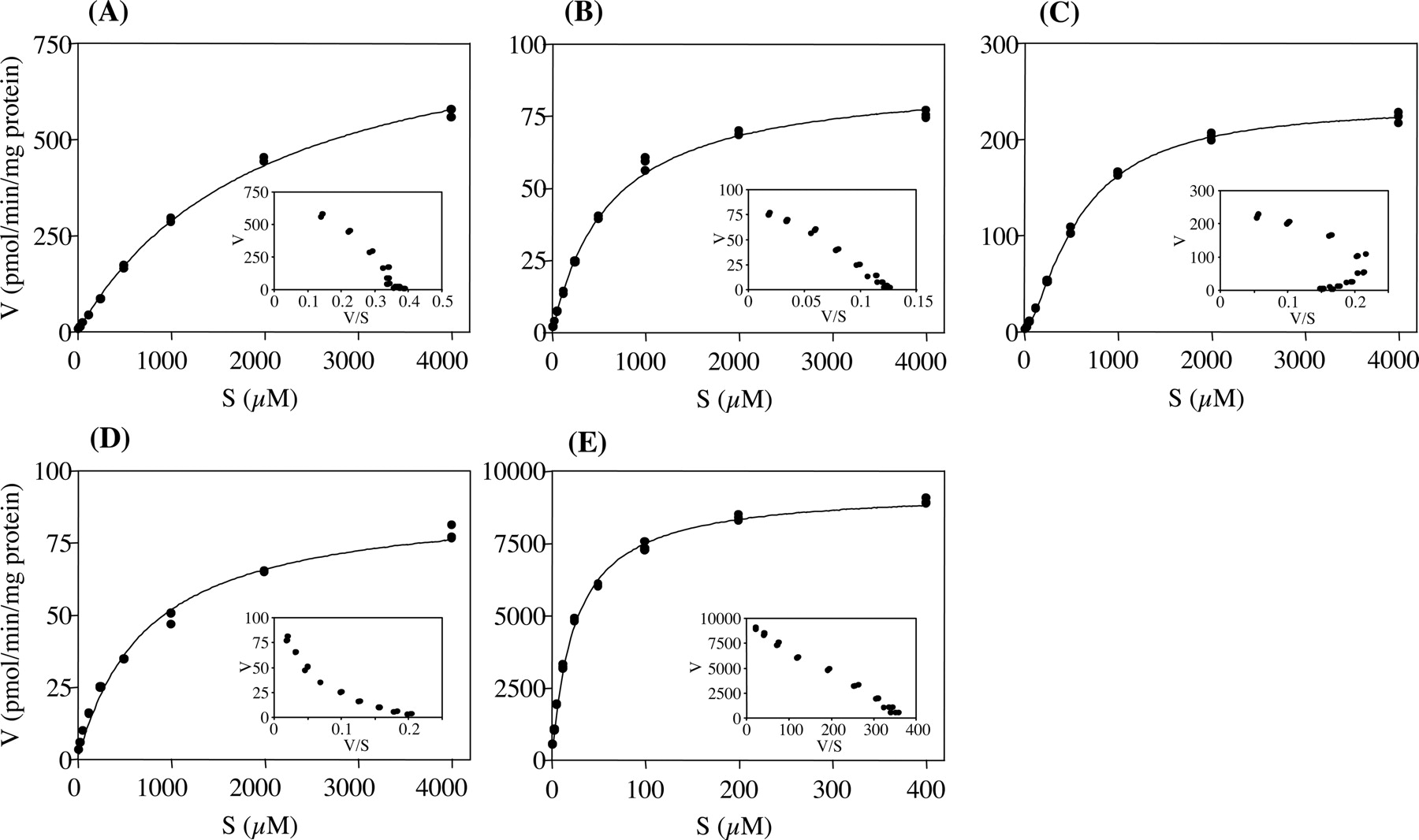
Kinetics of AFQ N-glucuronidation in human (A), rat (B), dog (C), monkey (D), and rabbit (E) liver microsomes. The concentration of AFQ ranged from 15.6 to 4000 μM (A-D) and 1.56 to 400 μM (E). AFQ N-glucuronosyltransferase activities were determined as described under Materials and Methods. Each inset shows the Eadie-Hofstee plot of the experimental data. Each incubation was performed by triplicate determinations.
Kinetic parameters of AFQ N-glucuronidation in liver microsomes from humans and experimental animals
The kinetic parameters were calculated with GraphPad Prism software. Each value represents best-fit values ± S.E. of triplicate points. Kinetic plots are presented in Fig. 3.
AFQ N-Glucuronidation in Recombinant UGT Isoforms. All recombinant UGT isoforms expressed in baculovirus-infected insect cells (Supersomes), which are commercially available, were used to determine their AFQ N-glucuronosyltransferase activities (Fig. 4). UGT1A3 (20.7 ± 0.1 pmol/min/mg protein) and UGT1A4 (28.2 ± 1.2 pmol/min/mg protein) exhibited high AFQ N-glucuronosyltransferase activities. UGT1A1, UGT1A9, UGT1A10, and UGT2B15 also exhibited low AFQ N-glucuronosyltransferase activities (1.2-4.4 pmol/min/mg protein). All other isoforms (UGT1A6, UGT1A7, UGT1A8, UGT2B4, UGT2B7, and UGT2B17) had very low AFQ N-glucuronosyltransferase activities (<1.0 pmol/min/mg protein) or no activities.
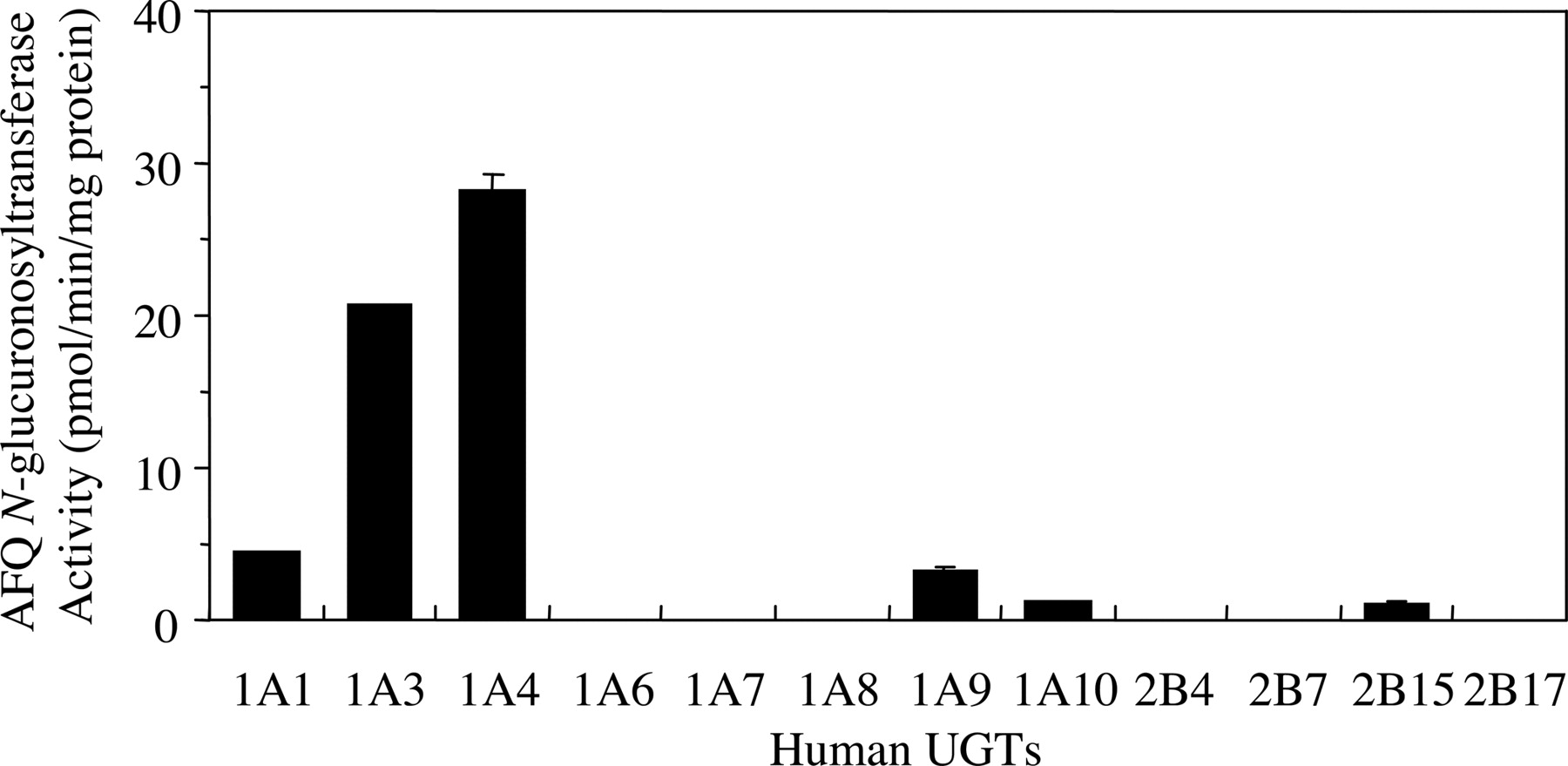
AFQ N-glucuronosyltransferase activity in recombinant human UGTs. AFQ N-glucuronosyltransferase activities in recombinant UGTs were determined at 100 μM AFQ. Each column represents the mean ± S.D. of triplicate determinations. The lower limit of quantitation of the assay under these conditions was 1.0 pmol/min/mg protein.
Kinetics of AFQ N-Glucuronidation in Recombinant UGTs, and Human Jejunum Microsomes. Kinetics analyses of the AFQ N-glucuronidation in recombinant UGT1A1, UGT1A3, and UGT1A4 microsomes were performed. As shown in Fig. 5, A and C, UGT1A1- and UGT1A4-catalyzed glucuronidation followed sigmoidal kinetics, which manifests as curvilinear Eadie-Hofstee plots. In contrast, the kinetics of AFQ N-glucuronidation in UGT1A3 at 15.6 to 1000 μM AFQ fitted to the Michaelis-Menten kinetics. However, when the AFQ concentration exceeded 1000 μM, the AFQ glucuronosyltransferase activity gradually decreased (Fig. 5B). A possible explanation for this phenomenon is substrate inhibition or metabolite inhibition. In addition, kinetic analysis of the AFQ N-glucuronidation in human jejunum microsomes was performed. The AFQ N-glucuronidation by human jejunum microsomes displayed a typical Michaelis-Menten kinetics (data not shown). Fitting the data points to eq. 1, 2, or 3 yielded the kinetic parameters listed in Table 2. Incubation of various concentrations (15.6-4000 μM) of AFQ with human jejunum microsomes demonstrated that the apparent Km, Vmax, and CLint values were 281 ± 17.3 μM, 111 ± 1.9 pmol/min/mg protein, and 0.396 μl/min/mg protein, respectively. The S50 values for UGT1A1 and UGT1A4 were 394 ± 16.6 and 1547 ± 54.3 μM, respectively. Corresponding Vmax values were 36 ± 0.7 and 799 ± 14.5 pmol/min/mg protein, respectively. Corresponding CLmax values were estimated to be 0.047 and 0.304 μl/min/mg protein, respectively. With respect to UGT1A3, when the apparent enzyme kinetic parameters were estimated by fitting to the Michaelis-Menten eq. 1, with the initial velocity values at 15.6 to 1000 μM AFQ, the Km, Vmax, and CLint values were 355 ± 94.0 μM, 120 ± 13.7 pmol/min/mg protein, and 0.337 μl/min/mg protein, respectively. When the apparent enzyme kinetic parameters were estimated by fitting to the substrate inhibition eq. 2, with the initial velocity values at 15.6 to 4000 μM AFQ, apparent Km, Ksi, and Vmax values were 1959 ± 1448 μM, 332 ± 248 μM, and 502 ± 323 pmol/min/mg protein, respectively. The CLint value was estimated to be 0.256 μl/min/mg protein.

Kinetics of AFQ N-glucuronidation in recombinant UGT1A1 (A), UGT1A3 (B), and UGT1A4 (C). A and C, the solid line represents the fitting curves to the Hill equation, 15.6 to 4000 μM). B, the solid line represents the fitting curves to the Michaelis-Menten equation, 15.6 to 1000 μM (closed circles). The dotted line represents the fitting curves to the substrate inhibition equation, 15.6 to 4000 μM (closed and open circles). AFQ N-glucuronosyltransferase activities were determined as described under Materials and Methods. Each inset shows the Eadie-Hofstee plot of the experimental data. Each incubation was performed by triplicate determinations.
Kinetic parameters of AFQ N-glucuronidation in recombinant UGT isoforms and in human liver (HLM) and jejunum microsomes (HJM)
AFQ (15.6-4000 μM) was incubated with microsomes and UDP-glucuronic acid for 45 min. The kinetic parameters were calculated with GraphPad Prism software. Each value represents best-fit values ± S.E. of triplicate points. Kinetic plots are presented in Fig. 5.
Inhibition Analyses of AFQ N-Glucuronidation in Human Liver and Jejunum Microsomes, and Recombinant UGTs. The inhibitory effects of trifluoperazine (UGT1A4) and genistein (UGT1A3 and UGT1A4) on the AFQ N-glucuronosyltransferase activities in human liver and jejunum microsomes were determined. As shown in Fig. 6A, the AFQ N-glucuronosyltransferase activities in human liver microsomes were prominently inhibited by trifluoperazine (IC50 = 59.5 μM) and effectively inhibited by genistein (IC50 = 151.2 μM). In contrast, the AFQ N-glucuronosyltransferase activities in human jejunum microsomes were effectively inhibited by trifluoperazine and genistein (IC50 = 150.0 and 177.8 μM, respectively; Fig. 6B). In addition, to evaluate the selectivity of trifluoperazine and genistein against UGT1A3 or UGT1A4, the inhibitory effects of each compound for UGT1A3 and UGT1A4 were also determined (Fig. 6, C and D). Trifluoperazine inhibited dramatically the AFQ N-glucuronidation activities in UGT1A4 (IC50 = 43.2 μM), with no effect on UGT1A3. In contrast, genistein inhibited the AFQ N-glucuronidation activities in UGT1A3 (IC50 = 117.4 μM) more effectively than those in UGT1A4 (IC50 = 220.5 μM).

Inhibitory effects of trifluoperazine and genistein for UGT isoforms on AFQ N-glucuronosyltransferase activity in human liver (A), jejunum (B), recombinant UGT1A3 (C), or UGT1A4 (D) microsomes. AFQ N-glucuronosyltransferase activities at 100 μM AFQ in pooled human liver, jejunum, UGT1A3, and UGT1A4 microsomes were determined as described under Materials and Methods. Trifluoperazine (UGT1A4) and genistein (UGT1A3 and UGT1A4) were used as inhibitors. Corresponding control activities were 37.0 ± 0.64, 25.9 ± 0.20, 22.3 ± 0.70, and 27.9 ± 0.23 pmol/min/mg protein, respectively. Each value represents the mean ± S.D. of triplicate determinations.
Interindividual Variability of AFQ N-Glucuronosyltransferase Activity from Seven Human Livers, and Correlation Analyses. AFQ N-glucuronosyltransferase activities in microsomes from seven human livers were determined at 100 μM AFQ (Fig. 7A). The interindividual difference in AFQ N-glucuronosyltransferase activity was at most 2.1-fold (25.6 ± 0.6 to 54.7 ± 0.2 pmol/min/mg protein). Correlation analyses were performed between the AFQ N-glucuronosyltransferase activity and β-estradiol (for the 3-OH position) (UGT1A1), trifluoperazine (UGT1A4), or propofol (UGT1A9) glucuronosyltransferase activities provided by the manufacturer. As shown Fig. 7, B and C, the AFQ N-glucuronosyltransferase activities in seven human liver microsomes were significantly correlated with the β-estradiol 3-glucuronosyltransferase activities (r2 = 0.697, p < 0.05) and trifluoperazine N-glucuronosyltransferase activities (r2 = 0.798, p < 0.01). However, AFQ N-glucuronosyltransferase activities did not correlate with the propofol glucuronosyltransferase activities (r2 = 0.260, p = 0.242; Fig. 7D).

Interindividual variability in AFQ N-glucuronosyltransferase activity in microsomes from seven human livers (A), and correlation analyses with enzymatic activities (B, C, and D). AFQ N-glucuronosyltransferase activities in microsomes from seven human livers were determined at 100 μM AFQ (A). Each column represents the mean ± S.D. of triplicate determinations. Correlation analysis between AFQ N-glucuronosyltransferase activity and β-estradiol 3-glucuronosyltransferase activity (B), trifluoperazine N-glucuronosyltransferase activity (C), or propofol glucuronosyltransferase activity (D) in microsomes from seven human livers.
Discussion
It has been reported that when AFQ is administered orally to humans, rats, dogs, and monkeys, the parent AFQ and its metabolites are excreted into urine (Furuuchi et al., 1983; Otsuka et al., 1983a,b). Although AFQ N-glucuronide is the most abundant metabolite in human urine, it has not been detected in rats, dogs, and monkeys. To clarify the species differences in N-glucuronidation on the metabolism of AFQ, we focused on the investigation of AFQ N-glucuronosyltransferase activities using liver microsomes obtained from humans and experimental animals. The Vmax and clearance (CLint) of the AFQ N-glucuronidation in human liver was approximately 4- to 10-fold and 2- to 4-fold higher than those in rat, dog, and monkey, respectively. The major metabolic pathways in AFQ consist of acetylation of the aromatic amino group followed by hydroxylation at the methyl carbon of either the acetyl or 2′-methyl residue and direct conjugation of AFQ with glucuronic acid at the aromatic amino group, whereas the N-acetylated metabolites have not been found in dogs. Dramatic species differences in the urinary excretion rate of AFQ N-glucuronide between humans and experimental animals might be also attributed to other metabolic pathways described above. In rats and monkeys, the major metabolic pathways of AFQ were acetylation of the aromatic amino group followed by hydroxylation at the methyl carbon of the acetyl group (Otsuka et al., 1983a,b). In dogs, AFQ was monohydroxylated at the 2-, 2′-, 3′-, and principally 4′-positions. In addition, these three animal species excreted the novel sulfur-containing metabolites such as the 2-methylsulfinylmethyl and 2-methylsul-fonylmethyl derivatives of AFQ, whereas these metabolites have not been found in humans. Another explanation is that there is high deacetylase activity in human compared with that in rat or monkey. It has been reported that, upon incubation of benzidine with liver slices from rats and humans, different proportions of this diamine were N-acetylated and N-glucuronidated (Babu et al., 1994; Zenser et al., 1998). Little glucuronidation was observed in rats with N,N′-diacetylbenzidine as the major metabolite of benzidine. In contrast, human liver slices demonstrated both extensive N-acetylation and N-glucuronidation activities. Authors have discussed the possibility that differences between rats and humans might be attributed to rapid deacetylation by human liver with N-acetylbenzidine rather than an accumulation of N,N′-diacetylbenzidine. Incidentally, although it has not been determined in an in vivo study in rabbit, it is interesting that the clearance of the AFQ N-glucuronidation in rabbit liver was approximately 1000-fold higher than that in human liver. Chiu and Huskey (1998) have reported that rabbit and guinea pig appear to exhibit the highest capacity of primary amine N-glucuronidation among preclinical species. That finding was consistent with our present results.
The identification of the enzymes involved in a metabolic pathway is important to understand interindividual variation in drug metabolism and drug-drug interaction. A number of approaches have been developed for identification of human UGTs involved in the glucuronidation of endogenous and exogenous compounds in vitro (Ritter, 2000; Tukey and Strassburg, 2000). To identify the human UGT isoform(s) responsible for AFQ N-glucuronidation, we first examined the activities and kinetics in commercially available recombinant UGT microsomes from baculovirus-insect cells. In consequence, UGT1A3 and UGT1A4 possessed the highest AFQ N-glucuronosyltransferase activities among the UGT isoforms examined (Fig. 4). UGT1A1, UGT1A9, UGT1A10, and UGT2B15 also catalyzed the AFQ N-glucuronidation. These findings suggest that multiple UGT isoforms are involved in the glucuronidation of AFQ in human liver and intestine. To clarify the contribution of UGT isoforms for AFQ N-glucuronidation, inhibition studies and correlation analyses with typical substrates for UGT isoforms were performed using human liver and jejunum microsomes. Trifluoperazine, a specific substrate for UGT1A4 (Green and Tephly, 1996; Nakajima et al., 2002; Ghosal et al., 2004), inhibited equivalently the AFQ N-glucuronosyltransferase activities in human liver and recombinant UGT1A4 microsomes, with a similar IC50 value (59.5 versus 43.2 μM). In addition, the activity in human liver microsomes was inhibited almost completely without the effect on the AFQ N-glucuronidation by UGT1A3 (Fig. 6, A, C, and D). It was confirmed that the Km value of AFQ N-glucuronidation in UGT1A4 was consistent with that in human liver microsomes (Table 2). These results suggest that UGT1A4 would mainly catalyze the AFQ N-glucuronidation in human liver. With respect to UGT1A3, its kinetic pattern was different from that of human liver microsomes, in which it was an atypical substrate inhibition. Furthermore, since hepatic UGT1A3 mRNA levels varied, and approximately 20-fold and 5- to 10-fold less than the levels of UGT1A1 and UGT1A4, respectively (Mojarrabi et al., 1996), it is unlikely that UGT1A3 would play a role in the metabolism of AFQ in the liver. The AFQ N-glucuronosyltransferase activities in seven human liver microsomes were significantly correlated with the trifluoperazine N-glucuronosyltransferase activities. In addition, the AFQ N-glucuronosyltransferase activities were significantly correlated with the β-estradiol 3-glucuronosyltransferase activities, which were mainly catalyzed by UGT1A1 (Senafi et al., 1994; King et al., 1996). These results suggest that not only UGT1A4 but also UGT1A1 might catalyze the AFQ N-glucuronidation in human liver. UGT1A1 also possesses significant activity toward AFQ, and it is a major isoform in human liver. However, the contribution of UGT1A1 to the AFQ N-glucuronidation in human liver would be lower than that of UGT1A4, because the Km value of AFQ N-glucuronidation in UGT1A1 was different from that in human liver microsomes (394 ± 16.6 versus 2019 ± 85.9 μM; Table 2).
In contrast, the inhibition effect of trifluoperazine on the AFQ N-glucuronosyltransferase activities in human jejunum microsomes was relatively weak compared with that in human liver (IC50 value, 150.0 versus 59.5 μM). The Km value of the AFQ N-glucuronidation in human jejunum microsomes was different from that in human liver (281 ± 17.3 versus 2019 ± 85.9 μM; Table 2). These results suggest that UGT1A4 and other UGT isoforms would catalyze the AFQ N-glucuronidation in human intestine. Incidentally, the kinetic pattern of UGT1A3 was an atypical substrate inhibition, in which the kinetic constants determined by the substrate inhibition model were inappropriate to describe the kinetics due to the high apparent standard errors (Fig. 5, Table 2). In addition, the maximum plasma concentrations of AFQ have been reported to be about 0.2 μM in humans taking AFQ at therapeutic doses of 60 mg/day (Miura et al., 1985). In clinical use, the concentration of AFQ is unlikely to reach 1000 μM. Therefore, we adopted the kinetic parameter fitted to the Michaelis-Menten equation with <1000 μM AFQ for UGT1A3. It was confirmed that the Km value of AFQ N-glucuronidation in UGT1A3 was relatively close to that in human jejunum microsomes. The AFQ N-glucuronosyltransferase activities in recombinant UGT1A4 were inhibited by trifluoperazine more effectively than by genistein, a substrate for UGT1A3 and UGT1A4 (Green et al., 1998; King et al., 2000). The inhibition pattern of these two compounds in human liver microsomes was similar to that in recombinant UGT1A4, whereas the inhibition effects of these two compounds on the AFQ N-glucuronosyltransferase activities in human jejunum microsomes were apparently comparable. It is considered that this phenomenon would be due to the contribution of UGT1A3 to the AFQ N-glucuronosyltransferase activities. These results suggest that UGT1A4 and UGT1A3 would mainly catalyze the AFQ N-glucuronidation in human intestine. Unfortunately, we could not examine clearly the contribution of UGT1A3 isoform to the AFQ N-glucuronidation in human jejunum microsomes due to the absence of specific substrates or inhibitors for UGT1A3. It has been reported that primary amine glucuronidation was seen with UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A8, UGT1A9, and UGT2B7 (Green et al., 1998; Green and Tephly, 1998; Ciotti et al., 1999; Hiller et al., 1999; Kaivosaari et al., 2002). The present results were almost consistent with these reports. Although it has been extensively reported that, among many UGT isoforms, UGT1A3 and UGT1A4 catalyze exclusively the glucuronidation of tertiary amine compounds to quaternary ammonium-linked glucuronides (Green et al., 1998; Green and Tephly, 1998), it is interesting that UGT1A3 and UGT1A4 play a major role in the metabolism of primary amines such as AFQ.
In conclusion, species differences in AFQ N-glucuronidation using liver microsomes obtained from humans and experimental animals showed that the catalytic activity in human liver was higher than that in the experimental animals except for rabbit. Furthermore, these results are consistent with the clinical data, in which AFQ N-glucuronide is the major metabolite in human urine. We found that the AFQ N-glucuronidation in humans is mainly catalyzed by UGT1A4 in the liver and by UGT1A3 as well as UGT1A4 in the intestine.
Acknowledgments
We wish to express our deep gratitude to Dr. Toru Ishizuka, Director of Exploratory Toxicology and DMPK Research Laboratories, for his interest and encouragement. The technical assistance of Hiroyuki Hashimoto is gratefully acknowledged.
Footnotes
-
doi:10.1124/dmd.104.001925.
-
ABBREVIATIONS: AFQ, afloqualone; UGT, UDP-glucuronosyltransferase; HPLC, high-performance liquid chromatography; LC-MS/MS, liquid chromatography-tandem mass spectrometry; CLint, intrinsic clearance; CLmax, maximum clearance.
- Received August 22, 2004.
- Accepted October 6, 2004.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}