


Abstract
The specificities of orphenadrine and methimazole on eight human liver P450 enzyme activities were evaluated by studying the extent of inhibition at different concentrations in two protocols: competitive inhibition and preincubation. In the competitive inhibition protocol, orphenadrine decreased CYP2B6 marker activity up to 45–57% in human liver microsomes and up to 80–97% in cell microsomes containing cDNA-expressed CYP2B6. Orphenadrine strongly decreased CYP2D6 marker activity by 80–90%. Orphenadrine also partially decreased the CYP1A2, CYP2A6, CYP3A4, and CYP2C19 marker activities. In the preincubation protocol, orphenadrine decreased the CYP2B6 activity in cDNA-expressed cell microsomes to completion. In human liver microsomes, orphenadrine strongly decreased the marker activities of CYP2B6, CYP2D6, as well as CYP2C9; and partially decreased the marker activities of CYP1A2, CYP2A6, CYP3A4, and CYP2C19. In the competitive inhibition protocol, methimazole had no effect on the marker activities of CYP2E1 and CYP2A6; slightly decreased CYP2D6 marker activity; partially decreased the marker activities of CYP2C19, CYP2C9, and CYP2B6; and dramatically decreased CYP3A4 marker activity. Methimazole decreased CYP1A2 marker activity at lower concentrations, but not at the highest concentration studied (1 mM). In the preincubation protocol, methimazole was shown to be a potent and nonspecific inhibitor of all the enzyme activities. Marker activities of CYP2C9, CYP2C19, and CYP3A4 were completely inhibited at relatively low concentrations. This study indicates orphenadrine cannot be used as a selective inhibitor of CYP2B6 in human liver microsomes and that methimazole is not a selective inhibitor of the flavin-containing monooxygenase in human liver microsomes.
The identification of specific P4501 enzymes responsible for the metabolism of new chemical entities is important information in the development of potential drugs. Together with the knowledge on the conditions that influence the expression and/or catalytic activity of that enzyme, predictions can be made as to whether or not environmental, medical, nutritional, chemical, or genetic factors could affect the metabolism of the new chemical entity. Predictions can also be made on potential drug–drug interactions (induction or inhibition). The use of chemical inhibitors is one of the approaches developed for such enzyme identification studies (1, 2). Chemical inhibitors are valuable and convenient tools because they are generally commercially available and can be used in intact cells and possibly for in vivo studies. However, the utility of chemical inhibitors is largely dependent on a thorough understanding of both their enzyme specificity and relative potency. Routinely, the selectivity of a P450 enzyme inhibitor was established mainly by inhibition of a particular P450 enzyme reaction and sometimes also by the absence of inhibition for a few other P450 activities. Only recently, the specificity of several widely used P450 enzyme inhibitors was examined by studying the reactions catalyzed by five major human liver P450 enzymes (3).
Orphenadrine, an antiparkinsonian agent, was shown to be a specific P4502B enzyme inhibitor in rat liver (4). The Ki for the inhibition of rat P4502B-catalyzed pentoxyresorufinO-depentylation was 3.8 μM without preincubation in the presence of NADPH and 0.13 μM with preincubation in the presence of NADPH (4). Orphenadrine (0.3 mM) has been used as a diagnostic inhibitor for CYP2B6-catalyzed reaction in human liver microsomes (5). However, the potency and selectivity of orphenadrine may be different between species. Methimazole is a high-affinity substrate for the flavin-containing monooxygenase (6). There have been reports suggesting a metabolism-dependent inhibition of P450 enzymes by methimazole (7-9). However, high concentrations of methimazole (≥1 mM) have often been used to differentiate the role of the flavin-containing monooxygenase from those of P450 enzymes in the catalyses of a particular reaction in liver microsomes (10, 11).
In this study, we examine the specificity of orphenadrine and methimazole toward eight marker P450 enzyme activities in human liver microsomes.
Materials and Methods
Chemicals and Reagents. Orphenadrine, methimazole, nifedipine, trizma base, coumarin, tolbutamide, chlorpropamide, chlorzoxazone, coumarin, 7-hydroxycoumarin, β-NADPH, NADP+, glucose-6-phosphate, and glucose-6-phosphate dehydrogenase were purchased from Sigma Chemical Co. (St. Louis, MO).S-mephenytoin, 4-hydroxy-S-mephenytoin, 4′-hydroxytolbutamide, and 6-hydroxychlorzoxazone were purchased from Ultrafine Chemicals (Manchester, UK). Bufuralol and 1′-hydroxybufuralol were purchased from Gentest Corp. (Woburn, MA). Perchloric acid was purchased from Fisher Scientific (Fair Lawn, NJ). HPLC-grade acetonitrile, methyl-t-butyl ether, isopropanol, methanol, ethyl acetate, and glacial acetic acid were obtained from EM Science (Gibbstown, NJ). All other chemicals are of reagent grade.
Human Liver Microsomes. Fresh human tissues were obtained from organ donors and supplied by the following procurement agencies: Anatomic Gift Foundation, National Disease Research Interchange, and the International Institute for the Advancement of Medicine. A pooled human liver microsomal sample was prepared from five different human liver samples: a 41-year-old White female, a 10-year-old African-American male, a 67-year-old White male, a 14-year-old White female, and a 62-year-old White male. Microsomes were prepared as described (12) and stored in aliquots at −80°C until use. Protein concentration was determined according to Lowry et al. (13) using bovine serum albumin as a standard. P450 contents were determined according to Omura and Sato (14). Microsomes containing cDNA-expressed human CYP2B6 were obtained from Gentest Corp.
Enzyme Assays. CYP1A2 marker phenacetinO-deethylase activity was determined at a phenacetin concentration of 100 μM essentially according to the procedure of Sattler et al. (15). Coumarin 7-hydroxylation was assayed for CYP2A6 as described previously (16) at a substrate concentration of 50 μM, and 7-ethoxytrifluoromethylcoumarin O-deethylation was assayed for CYP2B6 at a substrate concentration of 10 μM (17). CYP2C9 marker tolbutamide 4′-hydroxylation was assayed at a substrate concentration of 500 μM by a procedure slightly modified from Knodell et al. (18). CYP2C19 marker (S)-mephenytoin 4-hydroxylase activity was assayed at a substrate concentration of 200 μM by the published procedure of Meier et al. (19). Bufuralol 1-hydroxylation was assayed at a substrate concentration of 100 μM (20), and chlorzoxazone 6-hydroxylation was assayed for CYP2E1 at a substrate concentration of 500 μM as described previously (21). CYP3A4-catalyzed nifedipine oxidase activity was assayed at a substrate concentration of 200 μM according to the published method of Guengerich et al. (22). The use of 7-ethoxytrifluoromethylcoumarin O-deethylation as a marker activity for CYP2B6 in human liver microsomes has not been well established. Thus, microsomes containing cDNA-expressed CYP2B6 were used to exclude interference of other human P450 enzymes. All of the other substrates have been shown to be mainly metabolized by their corresponding human P450 enzymes (23).
Inhibition Study Design. Orphenadrine and methimazole were introduced to the incubation mixtures in water. Two protocols were used to study the effects of orphenadrine and methimazole on human liver P450 enzymes: a competitive inhibition protocol and a preincubation protocol to monitor for a metabolism-dependent component of enzyme inhibition. For the competitive inhibition studies, orphenadrine or methimazole was mixed at various concentrations with the substrate in the incubation mixture for 3 min at 37°C. The reaction was then initiated by an NADPH-generating system. For the preincubation protocol, orphenadrine or methimazole was preincubated at various concentrations in the presence of human liver microsomes and an NADPH-generating system for 15 min at 37°C. The reaction was then initiated by addition of the substrate.
Data Analysis. For all assays, analyte levels were determined using a standard curve generated from the authentic metabolite. Raw HPLC data was processed using the Millennium Chromatography Manager (version 2.1, Waters, Milford, MA). The human P450 enzyme-mediated activities in the presence of orphenadrine or methimazole were expressed as a percentage of the corresponding zero concentration values. All data points represent the average of duplicate incubations, with the differences between duplicates being <10%. Orphenadrine or methimazole at various concentrations was also incubated in a control without substrate to ensure no interference with the quantitation of respective products in the various assays. IC50 values were estimated using the Grafit software (version 3.0, Erithacus Software Ltd., Staines, UK).
Results and Discussion
The pooled microsomal sample used for this study had a total P450 content of 0.46 nmol/mg protein. The control CYP marker activities in this sample were: phenacetin O-deethylation (CYP1A2), 0.69 nmol/min/mg protein; coumarin 7-hydroxylation (CYP2A6), 0.77 nmol/min/mg protein; 7-ethoxytrifluoromethylcoumarinO-deethylation (CYP2B6), 80 pmol/min/mg protein; tolbutamide hydroxylation (CYP2C9), 0.32 nmol/min/mg protein; (S)-mephenytoin 4-hydroxylation (CYP2C19), 80 pmol/min/mg protein; bufuralol 1′-hydroxylation (CYP2D6), 205 pmol/min/mg protein; chlorzoxazone 6-hydroxylation (CYP2E1), 1.98 nmol/min/mg protein; and nifedipine oxidation (CYP3A4), 3.06 nmol/min/mg protein. The 7-ethoxytrifluoromethylcoumarin O-deethylase activity was 95 pmol/min/mg protein for microsomes containing cDNA-expressed CYP2B6 at a substrate concentration of 10 μM. CYP2A6, CYP2B6, and CYP2E1 activities were reduced by ∼20%, and CYP3A4 activity was reduced by ∼15% when human liver microsomes were preincubated for 15 min at 37°C with NADPH.
The estimated IC50 values of orphenadrine and methimazole for different human P450 enzymes under the two protocols are summarized in table 1. The inhibition of orphenadrine on CYP2B6 and CYP2D6 activities and the inhibition of methimazole on the activities of CYP2C9, CYP3A4, and CYP2C19 (with preincubation) are most evident.
Estimated IC50 values (μM) of orphenadrine and methimazole for different human P450 enzymes
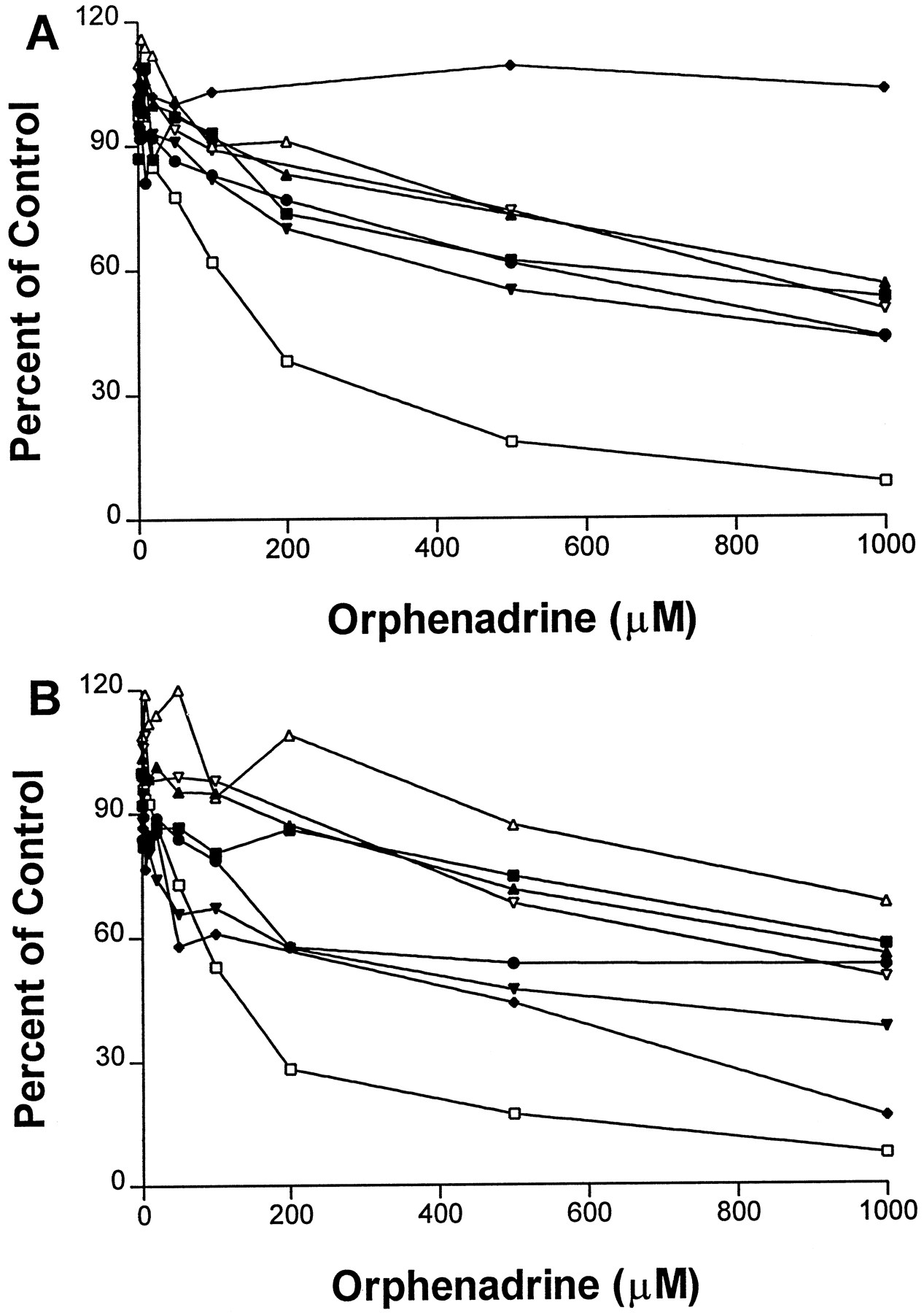
Results for orphenadrine with the competitive inhibition protocol are shown in fig. 1A. Orphenadrine decreased 7-ethoxytrifluoromethylcoumarin O-deethylase activity in human liver microsomes (mainly by CYP2B6) in a concentration-dependent manner, up to 45–57% at 500–1000 μM. At the same concentrations, orphenadrine decreased the 7-ethoxytrifluoromethylcoumarinO-deethylase activity in CYP2B6 cDNA-expressed cell microsomes by 80–97% (fig. 2). Surprisingly, orphenadrine decreased CYP2D6-catalyzed bufuralol 1′-hydroxylation by 80–90% at 500–1000 μM. Orphenadrine had no effect on CYP2C9 activity, but partially inhibited the activities of CYP1A2, CYP2A6, CYP3A4, and CYP2C19 by 40–60% at 1000 μM.

Effect of orphenadrine on human P450 enzyme-catalyzed reactions in human liver microsomes.
(A) Orphenadrine in the competitive inhibition protocol and (B) orphenadrine in the preincubation protocol. Enzyme reactions assayed are: phenacetin O-deethylation for CYP1A2 (▪); coumarin 7-hydroxylation for CYP2A6 (▴); 7-ethoxytrifluoromethylcoumarin O-deethylation for CYP2B6 (▾); tolbutamide 4′-hydroxylation for CYP2C9 (⧫); (S)-mephenytoin 4-hydroxylation for CYP2C19 (•); bufuralol 1-hydroxylation for CYP2D6 (□); chlorzoxazone 6-hydroxylation for CYP2E1 (▵); and nifedipine oxidation for CYP3A4 (▿).

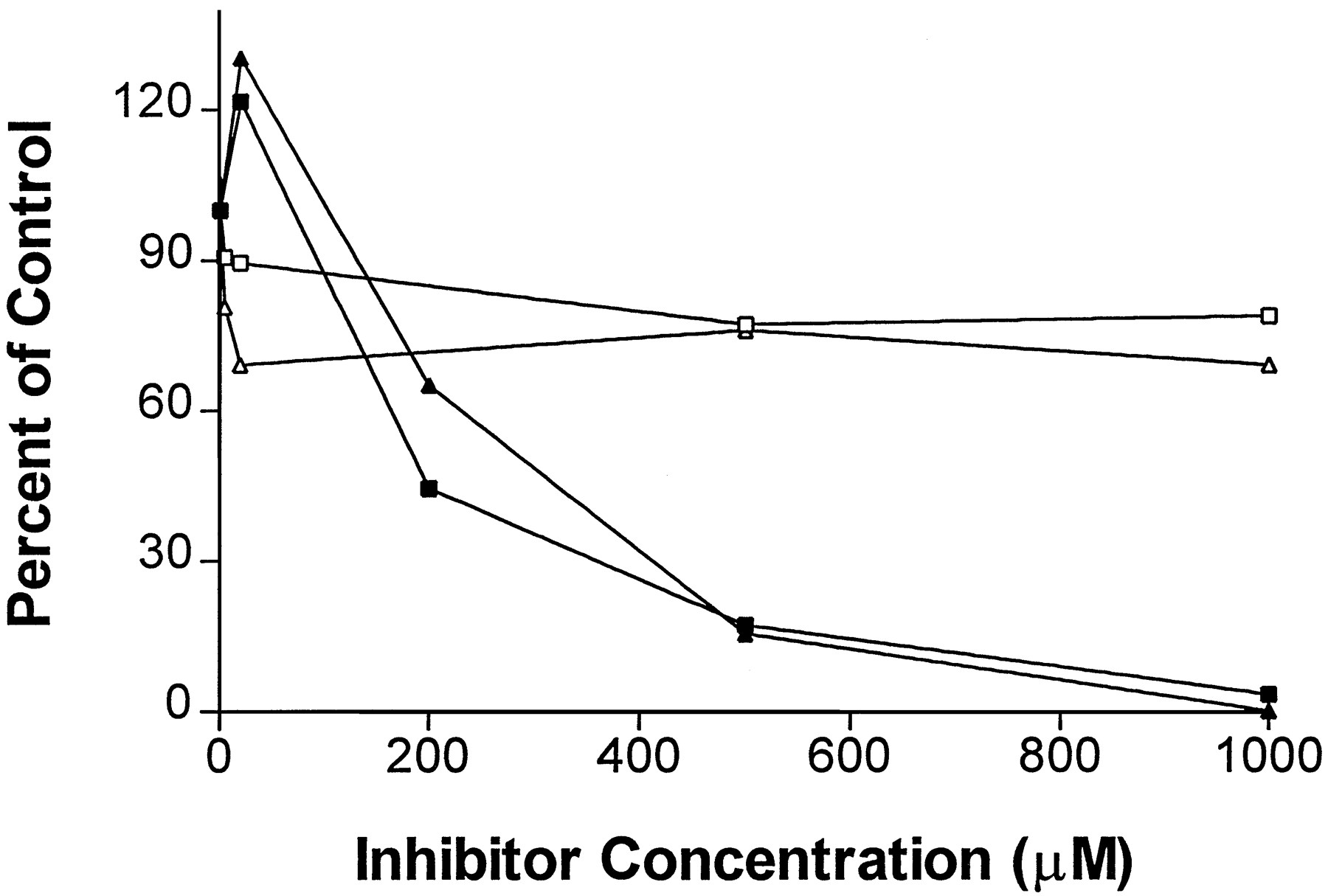
Effect of orphenadrine and methimazole on expressed CYP2B6-catalyzed 7-ethoxytrifluoromethylcoumarin O-deethylation.
▪, Orphenadrine in the competitive inhibition protocol; ▴, orphenadrine in the preincubation protocol; □, methimazole in the competitive inhibition protocol; and ▵, methimazole in the preincubation protocol.
Results for orphenadrine with the preincubation protocol are shown in figs. 1B and 2. In contrast to the competitive inhibition protocol, orphenadrine strongly decreased CYP2C9 marker tolbutamide hydroxylase activity by ∼60–80% at 500–1000 μM. For all other enzyme activities, the results for the mechanism-based inactivation protocol were very similar to those for the competitive inhibition protocol.
Orphenadrine decreased the 7-ethoxytrifluoromethylcoumarinO-deethylase activity in CYP2B6 cDNA-expressed cell microsomes to a much greater extent (fig. 2) than that in human liver microsomes (fig. 1). This may be due to the roles of other human P450 enzymes in catalyzing the reaction.
Results for methimazole with the competitive inhibition protocol are shown in fig. 3A. Methimazole had no effect on CYP2E1 and CYP2A6 marker activities; slightly decreased CYP2D6 activity (<25%); decreased CYP2C19 activity by 25–40% from 10 μM to 1 mM; and decreased CYP2C9 and CYP2B6 activities by 30–60% from 10 μM to 1 mM. Methimazole dramatically inhibited CYP3A4 activity by 30–82% at concentrations of 10 μM to 1 mM. Unexpectedly, methimazole decreased CYP1A2 activity at lower concentrations up to 67% at 50 μM, then the activity recovered at higher concentrations. Methimazole slightly decreased the expressed CYP2B6 activity (∼20%) (fig. 2).

Effect of methimazole on human P450 enzyme-catalyzed reactions in human liver microsomes.
(A) Methimazole in the competitive inhibition protocol and (B) methimazole in the preincubation protocol. Enzyme reactions assayed are: phenacetin O-deethylation for CYP1A2 (▪); coumarin 7-hydroxylation for CYP2A6 (▴); 7-ethoxytrifluoromethylcoumarin O-deethylation for CYP2B6 (▾); tolbutamide 4′-hydroxylation for CYP2C9 (⧫); (S)-mephenytoin 4-hydroxylation for CYP2C19 (•); bufuralol 1-hydroxylation for CYP2D6 (□); chlorzoxazone 6-hydroxylation for CYP2E1 (▵); and nifedipine oxidation for CYP3A4 (▿).
Results for methimazole with the preincubation protocol are shown in fig. 3B. Methimazole decreased all marker activities, and inhibition was more potent when compared with that seen in the competitive inhibition protocol. Activities of CYP2A6 and CYP2E1 were decreased by ∼30%; that of CYP2D6 was decreased by 20–50% at 20 μM to 1 mM; that of CYP2B6 was decreased by ∼80% at 10 μM or higher concentrations; and those of CYP2C9 and CYP2C19 were decreased by 40–90% at 10–200 μM with complete inhibition at >500 μM. CYP3A4 activity was decreased dramatically with >90% inhibition seen at >20 μM. CYP1A2 activity was decreased by 85% at 10–50 μM, and the activity was recovered gradually at higher concentrations. This effect of methimazole on CYP1A2 marker activity was confirmed by measuring the inhibition of CYP1A2-catalyzed theophylline oxidation in human liver microsomes (24) (data not shown). In this protocol, methimazole decreased the CYP2B6 activity in cDNA-expressed microsomes (∼30%) (fig. 2).
The specificity of commonly used human P450 enzyme inhibitors should be established by screening all the major human liver P450 enzyme marker activities (3). For example, 4-methylpyrazole, a presumed selective inhibitor of human CYP2E1, was found to inhibit CYP2D6 activity to a similar extent (3). Specificity of chemical inhibitors of human P450 enzymes also largely depends on the inhibitor concentration used. For example, ketoconazole and 7,8-benzoflavone strongly inhibited their respective P450 enzymes, CYP3A4 and CYP1A2 at low concentrations (<5 μM), but also inhibited several other enzymes at higher concentrations (3). In the present study, orphenadrine was shown to inhibit most human liver P450 enzymes, with stronger inhibition seen with CYP2D6 than CYP2B6. At concentrations higher than 0.5 mM orphenadrine, nearly complete inhibition of the expressed CYP2B6 activity was observed (fig. 2). This indicates that orphenadrine is not a selective inhibitor for CYP2B6 in human liver microsomes (5), even though it has been shown to be a very potent inhibitor of rat P4502B1 (4). This suggests orphenadrine, or other P450 enzyme inhibitors in general, may have different potencies and specificities in different species. Methimazole strongly inhibited a number of human liver P450 enzyme activities. The inhibitory effects of methimazole were much more pronounced in the preincubation protocol, with complete inhibition seen with CYP3A4, CYP2C9, and CYP2C19 at 500 μM or higher concentrations. This indicates that methimazole is not a selective inhibitor of the flavin-containing monooxygenase in human liver microsomes. The stronger inhibition of all eight human liver P450 enzymes in the preincubation protocol suggests that methimazole can be metabolized by the flavin-containing monooxygenase or P450 enzymes to a reactive species that inactivates microsomal enzymes (7-9).
Acknowledgments
We thank Dr. Michel Martinet for his support and useful discussions.
Footnotes
-
Send reprint requests to: Dr. Zuyu Guo, Department of Drug Metabolism and Pharmacokinetics, Rhone-Poulenc Rorer, NW12, 500 Arcola Road, Collegeville, PA 19426.
-
Received July 22, 1996; accepted November 25, 1996.
-
Present address for S. Raeissi: Department of Pharmaceutics, Uppsala University, Uppsala, Sweden.
- Abbreviations used are::
- P450
- cytochrome P450
- CYP
- cytochrome P450
- IC50
- inhibitory concentration of 50%
- The American Society for Pharmacology and Experimental Therapeutics



{kind=link}
{kind=link}
{kind=link}