


Abstract
3′-Azido-3′-deoxythymidine (AZT) is frequently prescribed to patients infected with the human immunodeficiency virus. After absorption, AZT is rapidly metabolized into 3′-azido-3′-deoxy-5′-glucuronylthymidine by UDP-glucuronosyltransferase (UGT) enzymes. Using labeled [14C]UDP-glucuronic acid and microsomal preparations from human kidney 293 cells stably expressing the different human UGT2B isoenzymes, it was demonstrated that AZT glucuronidation is catalyzed specifically by human UGT2B7. The identity of the metabolite formed was confirmed as AZT-G by liquid chromatography coupled with mass spectrometry. UGT2B7 is encoded by a polymorphic gene and kinetic analysis of AZT glucuronidation by the two allelic variants UGT2B7(H268) and UGT2B7(Y268), yielded apparent Km values of 91.0 and 80.1 μM, respectively. Normalization to protein levels yielded glucuronidation efficiency ratios (Vmax/Km) of 21.3 and 11.0 μl · min−1 · mg protein−1for UGT2B7(H268) and UGT2B7(Y268), respectively. It remains possible that other UGT enzymes are also involved in AZT conjugation; however, the glucuronidation of AZT by UGT2B7, which is a UGT2B protein expressed in the liver, is consistent with previous findings and supports the physiological relevance of this enzyme in AZT conjugation.
Drug therapy for HIV and AIDS infection generally includes treatment with nucleoside analogs, which act as HIV-reverse transcriptase (HIV-RT)2inhibitors (Patterson et al., 1997). 3′-Azido-3′-deoxythymidine (AZT) is the most studied member of the 2′,3′-dideoxynucleoside family of compounds, which has demonstrated clinical efficacy with decreased mortality and morbidity in patients infected with HIV (Fischl et al., 1987; Patterson et al., 1997). The mechanism of HIV-RT inhibition is based on the catabolic phosphorylation of the molecule, generating AZT-5′-triphosphate (AZT-TP), which is an analog of thymidine-5′-triphosphate, and acts as a competitor for the HIV-RT and a DNA-chain terminator (Mitsuya et al., 1990). However, toxic effects such as bone marrow suppression, vomiting, nausea (Mitsuya et al., 1990), circulating erythrocytes and neutrophil suppression (Moore et al., 1995), and mitochondrial myopathy (Mitsuya et al., 1990; Izuta et al., 1991), which are thought to be due to the interaction of AZT-TP with mammalian DNA polymerases β and γ (Izuta et al., 1991), are associated with this agent.
Before it can exert its antiviral effect by inhibiting HIV-RT, AZT must be phosphorylated by cellular enzymes to azidothymidine triphosphate (Fig. 1) (Lavie et al., 1998). This activation is catalyzed by successive kinases, beginning by the formation of AZT-monophosphate by thymidine kinase. The bottleneck of AZT activation lies in the second phosphorylation step, catalyzed by thymidylate kinase (Lavie et al., 1998). Cells treated with AZT accumulate the toxic AZT-monophosphate whereas only low concentrations of the active AZT-triphosphorylated (AZT-TP) are measured (2–10% of total phosphate metabolites) (Barry et al., 1996). The alternate pathway of AZT elimination from the body occurs primarily by glucuronidation (Good et al., 1990; Moore et al., 1995) with UDP-glucuronosyltransferase (UGT) enzymes (EC 2.4.1.17), which catalyze the transfer of the glucuronide moiety from UDP-glucuronic acid (UDPGA) to small hydrophobic molecules. The resulting compounds are generally more water-soluble, less toxic, and easier to excrete into the bile or urine (Dutton, 1980).

Structure and dual metabolism pathway of AZT (adapted from Patterson et al., 1997).
Based on divergent evolution, the mammalian UGT proteins have been categorized into two major families, UGT1 and UGT2 (Mackenzie et al., 1997). It has been thought that UGT1A enzymes were mainly involved in bilirubin and drug metabolism. However, several recent studies demonstrate that UGT1 can also glucuronidate steroid hormones (Albert et al., 1999). The UGT2 family is divided into two subfamilies, UGT2A, which are found in the olfactive epithelium (Jedlitschky et al., 1999) and UGT2B. Enzymes of the UGT2B subfamily catalyze the glucuronidation of several endogenous compounds, including bile acids, steroids, fatty acids, and carboxylic acids (Mackenzie et al., 1997), and exogenous substrates such as phenolic compounds and drugs (Coffman et al., 1998). To date, six human UGT2B cDNA clones have been isolated and characterized. UGT2B10 and UGT2B11 (Jin et al., 1993b; Beaulieu et al., 1998) demonstrate no reactivity after expression in human kidney 293 (HK293) cells, whereas UGT2B4, UGT2B7, UGT2B15, UGT2B17, and their allelic variants show overlapping substrate specificities, which could include AZT (Beaulieu et al., 1996; Lévesque et al., 1997, 1999;Coffman et al., 1998). In light of the interindividual variability that has been observed in the circulating level of AZT, it is important to identify the enzymes involved in AZT metabolism (Marchbanks et al., 1995; Fletcher and Balfour, 1996). At present, it is not known if the variable level of AZT in different patients is a result of altered phosphorylation and/or glucuronidation. Identifying the UGT enzyme(s) involved in AZT conjugation is required to assess their ability to influence the plasma and intracellular level of the drug. It will also be important to determine whether polymorphic variants of UGT proteins can influence the metabolism and elimination of AZT in different individuals harboring the different alleles. In this study, we report the identification of an AZT-glucuronidating enzyme, UGT2B7, and the characterization of AZT glucuronidation kinetics by human UGT2B7(H268) and its allelic variant UGT2B7(Y268).
Experimental Procedures
Materials.
UDPGA and eugenol were obtained from Sigma Chemical Co. (St. Louis, MO); AZT and 3′-azido-3′-deoxy-5′-glucuronylthymidine (AZT-G) were provided by Dr. Keith Gallicano (Bureau of Drug Research, Health Canada, Ottawa, Ontario, Canada). [14C]UDPGA was obtained from NEN Life Science Products Inc. (Boston, MA) and had a specific activity of 292.8 mCi · mmol−1 (10.8 GBq · mmol−1). HK293 cells were obtained from the American Type Culture Collection (Rockville, MD). Human liver microsomal preparations were obtained from Human Cell Culture Center, Inc. (Laurel, MD).
cDNA Isolation and Stable Expression of Human UGT2B.
cDNA isolation and HK293 cell lines stably expressing human UGT2B4, UGT2B7, UGT2B15, and UGT2B17 have been reported previously (Beaulieu et al., 1996; Lévesque et al., 1997, 1999; Coffman et al., 1998).
Microsomal Proteins Isolation.
Homogenization of 8 × 106 stably transfected HK293 cells was performed in 5 ml of homogenization buffer (0.1 M K2HPO4, pH 7.4, glycerol 20%, 1 mM EDTA, 1 mM dithiothreitol) and centrifuged at 12,000g, 4°C for 20 min. The supernatant was centrifuged at 105,000g for 1 h at 4°C. The microsomal pellets were resuspended in 0.5 ml of homogenization buffer.
Western Blot Analysis.
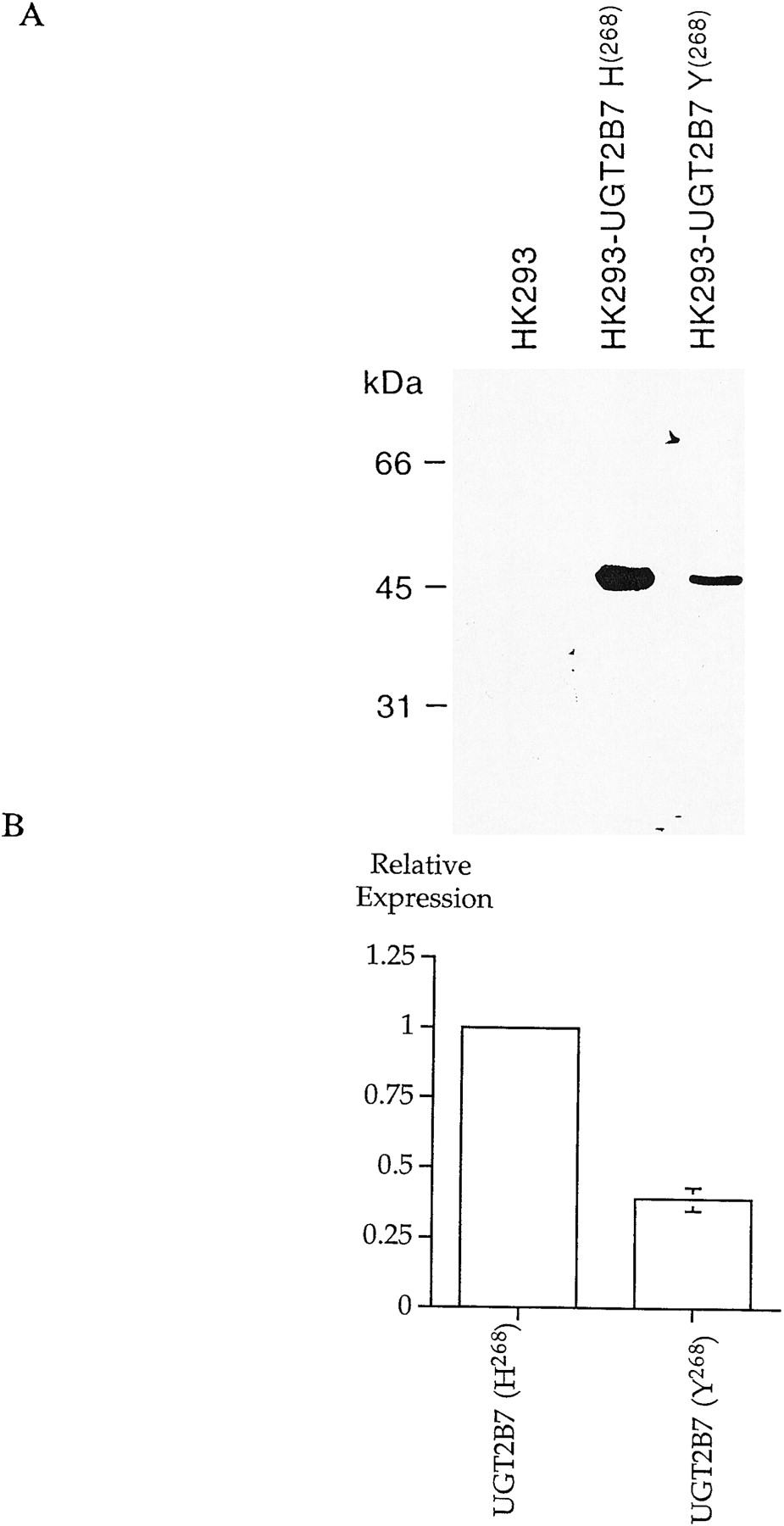
Microsomal proteins (20 μg) from HK293 cells and HK283 cells stably expressing UGT2B7(H268) and UGT2B7(Y268) were separated by 10% SDS-polyacrylamide gel electrophoresis. The gel was transferred onto nitrocellulose membrane and probed with anti-UGT2B17 EL-93 antisera (1:3000 dilution) (Guillemette et al., 1997). An anti-rabbit IgG horse antibody conjugated with horseradish peroxidase (Amersham Corp., Oakville, Canada) was used as the second antibody, and the resulting immunocomplexes were visualized using an enhanced chemiluminescence kit (Rennaissance, Quebec, Canada) and exposed on hyperfilm for 2 min (Kodak Corp., Rochester, NY) and quantified by BioImage Visage 110s (Genomic Solution, Inc., Ann Harbor, MI).
Glucuronidation Assay and KmDetermination Using Microsomal Proteins.
Screening assays were performed using 7.5 μM [14C]UDPGA, 92.5 μM unlabeled UDPGA, 200 μM AZT, and 20 μg of microsomal proteins from human liver and stably transfected HK293 cell lines in 50 mM Tris-HCl (pH 7.5), 10 mM MgCl2, 100 μg/ml phosphatidylcholine, and 8.5 mM saccharolactone in a final volume of 100 μl. Assays were performed for 16 h at 37°C, and were terminated by adding 100 μl of methanol. Chromatographic analysis and formation of glucuronide were estimated as described previously (Beaulieu et al., 1996).
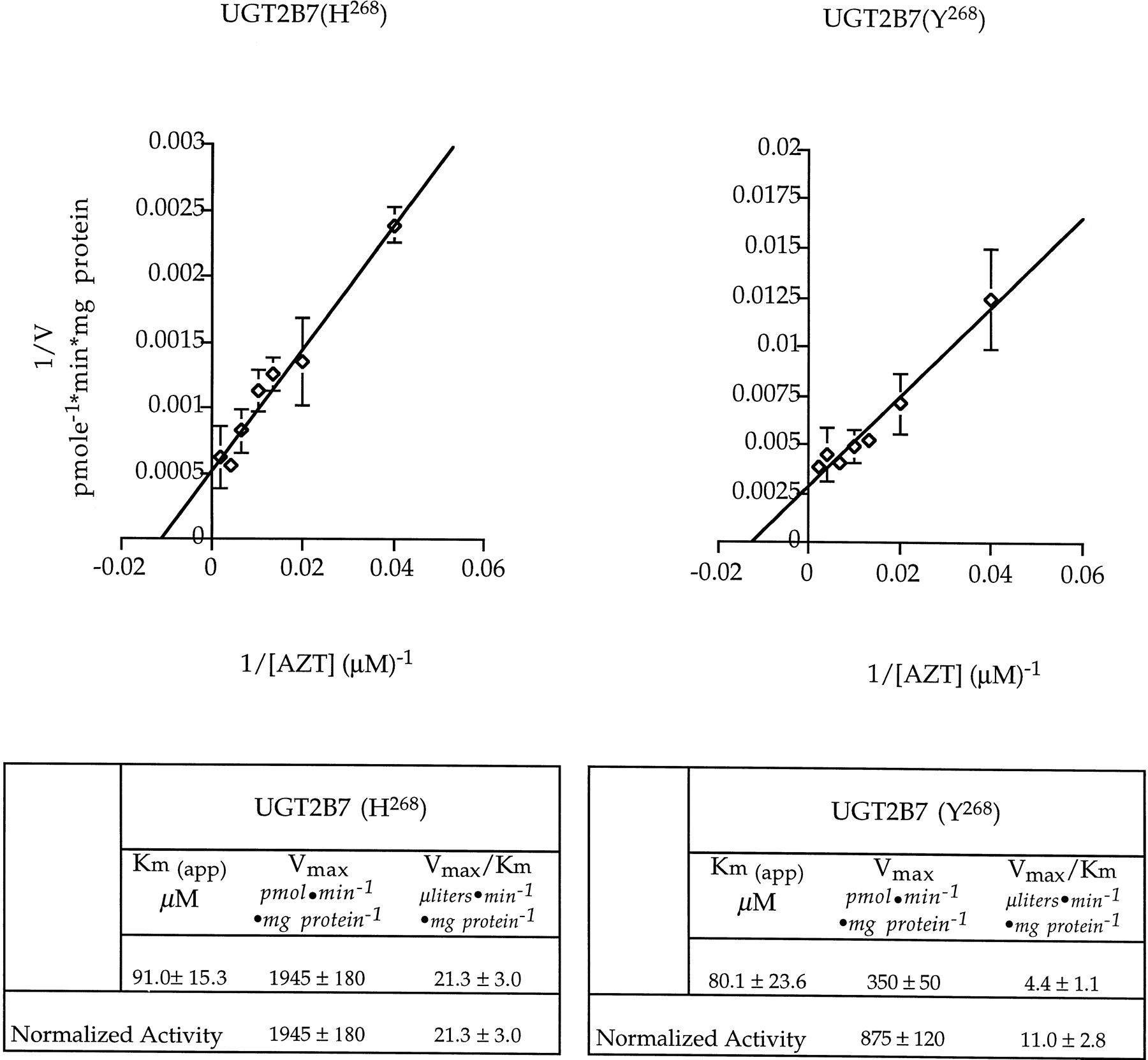
Proteins that demonstrated conjugation activity with AZT in the screening assay were subsequently reassayed in 50 mM Tris-HCl (pH 7.5) for 15 min to 12 h at 37°C in the presence of 7.5 μM [14C]UDPGA, 492.5 μM unlabeled UDPGA. Results demonstrated that AZT glucuronidation by both polymorphic UGT2B7 variants is linear for 4 h (data not shown). Kinetic analyses were realized for 30 min, using microsomal proteins in the same conditions with AZT concentration ranging from 100 to 10 mM as a first determination, and from 10 to 500 μM to obtain precise apparentKm and Vmaxvalues. The values, obtained by double reciprocal plots (Lineweaver-Burk), represent the mean of three independent experiments.
Liquid Chromatography Coupled with Mass Spectroscopy.
Increasing amounts of microsomal proteins from HK293 cells stably expressing UGT2B4, UGT2B7(H268), UGT2B7(Y268), UGT2B15, and UGT2B17 were assayed in the presence of 200 μM AZT and 500 μM unlabeled UDPGA, in the same conditions as screening assays. AZT and AZT-G standards or products from the activity assay (25 μl) were separated by HPLC on a zorbax C18 column (Hewlett-Packard, Boston, MA) using an Alliance 2690 system (Waters, Milford, MA). Elution was performed with a gradient of 1 mM acetonitrile-ammonium formate, from 2% in water-ammonium formate (1 mM) to 100% in 10 min (1 ml/min). Mass spectrometry, used to determine the molecular weight of compounds isolated by chromatography, was conducted using a Ion-Trap Mass Spectrometer, LCQ detector in negative electrospray ionization mode (Finnigan, San Jose, CA). Conditions of ionization were: 1) 225°C capillary temperature, 2) −45.00 V as capillary voltage, 3) 4kV as voltage source, 4) 80 U of sheath gas flow, 5) 1 U of auxiliary gas flow, 6) 500 ms as maximum injection time, and 7) total microscan of 2.
Results and Discussion
Considering the extensive use of AZT for treating HIV-infected individuals, this study was performed to identify the UGT enzyme(s) involved in conjugation and metabolism of this drug. To assess whether the UGT1A proteins are involved in the glucuronidation of AZT, previous studies demonstrated that neither bilirubin nor acetaminophen, which are conjugated by UGT1A enzymes, interact in vivo with the glucuronidation of AZT (Rajaonarison et al., 1992; Pacifici et al., 1996). In addition, AZT glucuronidation activity in Gunn rats, which have a deletion in the conserved portion of theirUGT1A gene resulting in a loss of all UGT1 activity, was comparable with normal rats (Haumont et al., 1990; Iyer et al., 1998). Together, these results suggest that glucuronidation of AZT is catalyzed by UGT2B subfamily enzymes. To identify the UGT2B enzyme that can conjugate AZT, a screening assay was performed with HK293 cell lines stably expressing UGT2B4, UGT2B7(H268), UGT2B7(Y268), UGT2B10, UGT2B11, UGT2B15, and UGT2B17. Incubation of microsomal proteins from the stable cells with AZT and labeled UDPGA demonstrated that only UGT2B7(H268) and the variant UGT2B7(Y268) can catalyze the transfer of glucuronic acid to AZT (Fig. 2). The other human UGT2B clones, UGT2B4, UGT2B15, and UGT2B17, did not demonstrate conjugation activity with this substrate, whereas they all catalyzed glucuronidation of eugenol, which was used as positive control (Fig. 2). The proteins UGT2B10 and UGT2B11, which thus far are not active on any substrates tested, also did not conjugate AZT (data not shown). The identity of the polar product formed by UGT2B7 as a glucuronide conjugate was confirmed by comparison of the retention time and molecular ion with the standards AZT and AZT-G as determined by liquid chromatography coupled with mass spectrometry (LC-MS/MS) (Fig. 3). Nonconjugated AZT remains present after incubation with microsomes containing UGT2B7 (H268) and UGT2B15 (Fig. 3, B and D); however, UGT2B7 (H268) yielded conjugated AZT-G, which is not observed with UGT2B15 (Fig. 3, C and E). In agreement with results from thin-layer chromatographic analysis, LC-MS/MS also did not detect any AZT-G when AZT was incubated with 20 and 100 μg of microsomal proteins from HK293 cells stably expressing UGT2B4 and UGT2B17 (data not shown).

Glucuronidation of AZT by human UGT2B proteins analyzed by thin layer chromatography.
Glucuronidation of AZT and eugenol (200 μM) by human liver and HK293 cells stably expressing human UGT2B4, UGT2B7(H268), UGT2B7(Y268), UGT2B15, and UGT2B17, and untransfected HK293 cells was assessed by incubation of microsomal protein with 7.5 μM [14C]UDPGA and 92.5 μM unlabeled UDPGA and revealed by thin layer chromatography. The free UDPGA is found at the bottom of the chromatogram.

LC-MS/MS analysis of AZT and AZT-G standard and AZT metabolites formed after incubation with UGT2B7(H268) and UGT2B15 microsomal proteins.
Microsomal proteins from UGT2B7(H268) and UGT2B15 stably expressing cell lines were incubated with 200 μM AZT and 500 μM UDPGA for 16 h at 37°C. The incubation products were then analyzed by liquid chromatography-mass spectrometry, and the retention times of metabolites were compared with standards of AZT and AZT-G. For each enzyme, a full mass scan (260–450m/z) was performed to detect both AZT and its glucuronide metabolite AZT-G (A); to refine the analysis, additional scans ranging between 265.9 and 267.9m/z (B), and 442.0 and 444.2m/z (C) were analyzed to detect specifically AZT and AZT-G, respectively; and the mass of the compounds detected in the scans were determined by mass spectrometry for AZT (D) and AZT-G (E). The y-axis represents the relative abundance of each compound relative to the highest peak value set at 100.
Although UGT2B enzymes are well known for their role in glucuronidation of endogenous compounds (Hum et al., 1999), UGT2B7 has also been demonstrated to conjugate several drugs, including opioids such as morphine, codeine, and nalorphine (Coffman et al., 1998). It was demonstrated that morphine caused an inhibition of AZT glucuronidation (Howe et al., 1992), suggesting a competition between the two drugs for the active site of UGT2B7. The expression of UGT2B7 was recently demonstrated in the human central nervous system, where the enzyme is suggested to play an important role in morphine analgesia by serving to generate the potent morphine-6-O-glucuronide (King et al., 1999). Therefore, it will be important to determine whether the decreased analgesic effect of morphine in AIDS patients treated with AZT is a result of competition between the two compounds for UGT2B7.
Consistent with a role of UGT2B7 in AZT glucuronidation in the liver, patients with liver diseases such as ethanol-induced cirrhosis have diminished hepatic morphine glucuronidation (reduced by 25%) (Crotty et al., 1989), which correlates with reduced plasma levels of AZT-glucuronide found in patients with similar pathologies (Taburet et al., 1990; Moore et al., 1995). UGT2B7 cDNA was cloned from a human liver cDNA library, and analysis of mRNA tissue distribution showed UGT2B7 highly expressed in human liver (Jin et al., 1993a; Hum et al., 1999).
In a study involving 93 different human liver microsomal preparations, it was demonstrated that AZT glucuronidation was neither sex- nor age-dependent, but displayed interindividual variability that ranged over one order of magnitude (Pacifici et al., 1996). Interestingly, the same variability was observed for the glucuronidation of the UGT2B7 substrates, (S)- and (R)-oxazepam, with human liver microsomes (Patel et al., 1995). To assess if the polymorphism of UGT2B7 may be implicated in the observed interindividual variability in AZT glucuronidation, kinetic analysis performed with microsomal proteins demonstrated that the two allelic variants of UGT2B7 have the same affinity for AZT, yielding apparent Km values of 91.0 and 80.1 μM for UGT2B7(H268) and UGT2B7(Y268), respectively (Fig.4). The apparentVmax values were 1900 and 350 pmol · min−1 · mg protein−1 for UGT2B7(H268) and UGT2B7(Y268), respectively, leading to AZT glucuronidation efficiencies (ratioVmax/Km) of 21.3 and 4.4 μl · min−1 · mg protein−1 (Fig. 4). The difference observed between the apparent Vmax of AZT glucuronidation by the two isoforms is due in part to the 2.5-fold higher expression of UGT2B7(H268) in comparison to UGT2B7(Y268) (Fig.5). Normalization of the apparentVmax values by the level of protein expression led to a difference of AZT glucuronidation efficiency of 1.9-fold (Fig. 4). Moreover, analysis of polymorphic UGT2B7 gene expression in 69 Caucasian subjects demonstrated that individuals homozygous for H268or Y268 were distributed equally in this population [frequencies of 26 and 24% for UGT2B7(H268) and UGT2B7(Y268), respectively], with heterozygotes comprising approximately 50% of the study population (Miners et al., 1998). These results suggest that the two known polymorphisms of UGT2B7 are similar in their AZT glucuronidation activities, and that polymorphism expression cannot explain the variability of AZT glucuronidation observed in human liver, or the variability in drug disposition observed with standard doses of AZT in patients (Mentre et al., 1993; Fletcher and Balfour, 1996). It remains possible that other uncharacterized polymorphisms of the UGT2B7 gene, or additional UGT enzymes may also be involved in AZT glucuronidation to explain this variability. Other parameters such as the competing phosphorylation pathway may also be implicated, which could be reflected by increased or decreased availability of the substrate for glucuronidation.

Lineweaver-Burk plots of AZT conjugation by UGT2B7(H268) and UGT2B7(Y268) in HK293 cell microsomes.
Values represent the mean ± S.D. of three independent experiments.

Western blot analysis of microsomes from HK293 cells and HK293 cells stably expressing UGT2B7(H268) or UGT2B7(Y268).
A, relative expression of UGT2B7(H268) and UGT2B7(Y268). B, microsomal proteins (20 μg) from untransfected or transfected HK293 cells were separated by 10% SDS-polyacrylamide gel electrophoresis, transferred onto nitrocellulose membrane, and probed with the anti-UGT2B17 EL-93 antisera. Levels of UGT2B7(H268) and UGT2B7(Y268) proteins were quantitated by BioImage Visage 110s.
In a previous study, Haumont et al. (1990) reportedKm values of 13 mM for AZT glucuronidation by human microsomes, which is 150-fold higher than theKm value obtained with UGT2B7. This difference may be partially attributed to intrinsic differences in the two systems used of liver microsomes that express multiple UGT enzymes versus microsomes expressing only UGT2B7.
Considering the wide use of AZT for the treatment of HIV infection, the variability observed in liver conjugation of this drug and the interindividual variability of plasma AZT levels (Mentre et al., 1993;Fletcher and Balfour, 1996), it is important to identify the enzymes involved in the formation of AZT metabolites. Genetic polymorphisms may yield variant metabolic enzymes that can lead to variations in AZT transformation. As well, identification of the enzymes involved is needed to assess the contribution of the different metabolic pathways in AZT metabolism. As an example, AZT requires intracellular phosphorylation to AZT-triphosphate before the inhibition of HIV replication (Lavie et al., 1998); therefore, it will be important to identify the components in the phosphorylation or glucuronidation pathway that may alter AZT metabolism and influence the pharmacokinetics of the drug in different individuals. This study demonstrates for the first time a specific UGT enzyme, UGT2B7, which conjugates AZT. Consistent with previous studies, this enzyme is from the UGT2B family and is expressed in human liver, which is a major site of AZT glucuronidation.
Acknowledgments
We thank Dr. Keith Gallicano for providing AZT and AZT-G, and Patrick Bélanger for performing the LC-MS/MS analysis.
Footnotes
-
Send reprint requests to: Dr. Alain Bélanger or Dr. Dean W. Hum, Laboratory of Molecular Endocrinology, CHUL Research Center, 2705 Laurier Blvd., Quebec G1V 4G2, Canada. E-mail:Alain.Bélanger{at}crchul.ulaval.ca orDean.Hum{at}crchul.ulaval.ca
-
↵1 These authors contributed equally to this work.
-
This work was supported by the Medical Research Council of Canada (A.B. and D.W.H.), the Fonds de la Recherche en Santé du Québec (A.B. and D.W.H.), Endorecherche and the National Institutes of Health (GM26221 to T.R.T.). Olivier Barbier and David Turgeon are holders of scholarships from La Fondation de l'Université Laval, and Caroline Girard is holder of a Summer Research Fellowship from the American Endocrine Society.
- Abbreviations used are::
- HIV-RT
- HIV-reverse transcriptase
- AZT
- 3′-azido-3′-deoxythimidine
- AZT-G
- 3′-azido-3′-deoxy-5′-glucuronylthymidine
- AZT-TP
- AZT-5′-triphosphate
- UDPGA
- UDP-glucuronic acid
- HK293
- human kidney 293
- UGT
- UDP-glucuronosyltransferase
- LC-MS/MS
- liquid chromatography coupled with mass spectrometry
- Received December 10, 1999.
- Accepted February 16, 2000.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}