


Abstract
The UDP glucuronosyltransferases (UGT) are a family of enzymes in which substrates include drugs, xenobiotics, and products of endogenous catabolism. The main source of most UGT enzymes is the liver, a major organ in the detoxification and inactivation of compounds. Previous studies have indicated that glucuronidation, as measured by pharmacokinetic studies, is relatively spared in liver disease. Because UGT activity toward most substrates is the result of metabolism by different isoforms with overlapping specificities, these studies may not indicate the effect of disease on the levels of individual isoforms. We sought to extend these studies to the measurement of mRNA for individual isoforms in the liver of patients with various forms of liver disease. RNA was extracted from liver tissue samples of patients undergoing clinically necessary percutaneous liver biopsies. UGT mRNA levels for isoforms 1A1, 1A3, 1A4, 1A6, 1A9, 2B4, 2B7, 2B10, 2B11, 2B15, and 2B17 were determined by real-time reverse transcription-polymerase chain reaction. Biopsies were graded using the Metavir system. Results from patients with low fibrosis or inflammatory scores were compared with those with high scores. We found large interindividual variation in the levels of the various isoforms. This was greatest for UGT2B17. A consistent downward trend, reaching statistical significance for UGT1A4, UGT2B4, and UGT2B7, was observed in samples from patients with high inflammation scores. There was no such correlation with the degree of fibrosis. Our results indicate that hepatic UGT mRNA levels are reduced while the tissue is inflamed, but they are not affected in the noninflamed, chronically diseased liver.
It is well established that liver disease is associated with decreased elimination of many drugs. This is particularly so for those metabolized by enzymes of the cytochrome P450 family (Adedoyin et al., 1998; Rodighiero, 1999). Based mainly on pharmacokinetic studies, however, glucuronidation is usually comparatively well preserved in liver disease.
The interpretation of pharmacokinetic studies in liver disease is complicated by the expression of UGT1 in extrahepatic tissues (McGurk et al., 1998; Radominska-Pandya et al., 1998; Strassburg et al., 1998, 1999; Jedlitschky et al., 1999; Tchernof et al., 1999; Barbier et al., 2000). A further limitation of this approach is that human UGT comprises a superfamily of at least 15 isoforms bound to the membrane of the endoplasmic reticulum. There is considerable structural similarity between them, and they have overlapping substrate specificities.
Based on sequence homology, they can be divided into two subfamilies. The seven isoforms of family 2B are each encoded in a separate gene locus on chromosome 4. The eight isoforms of family 1A are coded for in an unusual arrangement on chromosome 2. Each of these proteins has an identical C terminus coded by 4 exons. The N termini differ and are each encoded by a specific exon, the final mRNA depending on which exon I product is spliced to the four constant regions (for recent review see Tukey and Strassburg, 2000).
The mechanism by which cytochrome P450 and UGT enzymes are altered in liver disease is poorly understood. Pathological components present to varying degrees in liver diseases include inflammation, cell loss, regeneration, and fibrosis. These in turn are due to a large number of changes in cellular function influenced by substances released from resident and inflammatory cells. At a gross level, it is possible to categorize a biopsy sample of diseased liver in terms of the degree of inflammation and fibrosis present. This categorization, although crude, permits correlation of each of these processes with the expression of a particular mRNA.
Although there is a number of studies on the expression of UGT mRNA in animal tissues (Debinski et al., 1996; Pellizzer et al., 1996; Strasser et al., 1998; Choo et al., 1999), there is little data on the expression of individual isoforms of UGT mRNA levels in the diseased human liver. The recent advent of real-time PCR greatly simplifies the process of detection and semiquantitation of mRNA from small biopsy tissue samples. The aim of this study was to use these methods to investigate the contribution of inflammation and fibrosis to the mRNA levels of individual UGT isoforms in patients with liver disease.
Materials and Methods
Sample Collection.
Surplus tissue (5–10 mg) available from patients undergoing percutaneous liver biopsy was snap frozen in liquid nitrogen and stored at −70°C until processing. Informed written consent was obtained from each patient, and the study protocol was approved by the St. Vincent's Hospital Human Ethics Committee.
Extraction of RNA.
RNA was extracted from the tissue by homogenization with a Polytron homogenizer in Trizol (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions. The extracted RNA was dissolved in 200 μl of water treated with 1% diethyl pyrocarbonate, and contaminating genomic DNA was digested with 1.5 units of DNase I (Promega, Madison, WI). The solution was extracted with Tris-equilibrated phenol, and the RNA precipitated with 2.5 volumes of ethanol. Finally, the RNA was dissolved in 100 μl of water, and aliquots removed for spectroscopic quantitation and agarose gel electrophoresis to assess RNA integrity. Approximately 5 μg of RNA was reverse transcribed in a reaction containing 10 U avian myeloblastosis virus reverse transcriptase, 1 mM dNTPs, 40 U RNasin (Promega), and 0.5 μg of random 10-mer oligonucleotides (GenSet Pacific, Lismore, Australia) for 10 min at room temperature and then for 60 min at 42°C. The resultant cDNA was diluted 10- to 20-fold and stored at −70°C until use.
PCR Primers.
Sequences of the PCR primers used for the detection of UGT isoforms were selected from UGT sequences aligned using DNAMAN software (Lynnon corporation, Quebec, Canada). Primers were made for the detection of UGT isoforms 1A1, 1A3, 1A4, 1A6, 1A9, 2B4, 2B7, 2B10, 2B11, 2B15, and 2B17. In addition, for the detection of all family 1 isoforms, primers were made to regions spanning exons 3 and 4 and, for the detection of all known family 2 isoforms, to regions spanning exons 1 to 3. Primers were also made for the detection of CYP1A2, IL-1β, and TGF-β. Sequences of all primers used in the PCR are shown in Table1.
Nucleotide sequences of primers used in the PCR reactions
Real-Time PCR.
PCR was performed on 5 μl of cDNA using 10 pmol of forward and reverse primers (Table 1; Sigma Genosys, Castle Hill, Australia) on a GenAmp model 5700 cycler using SYBR Green detection in a total volume of 20 μl with 10 μl of SYBR Green PCR Master-Mix (Applied Biosystems, Foster City, CA). Cycling conditions were 1 cycle for 2 min at 60°C, 1 cycle at 95°C for 10 min, followed by 40 cycles of a 15-s denaturation at 95°C, and a 1-min annealing/extension at 60°C. The threshold value of detection (Ct) is the cycle at which PCR product rises above an arbitrary number and was set at the exponential phase of amplification. Values were determined from standard curves generated for each gene from serial dilution of genomic or cDNA and normalized to the value of 18S ribosomal RNA measured for each sample. PCR products were of the correct size, as determined by agarose gel electrophoresis. Statistical significance was determined with StatView software (SAS Institute, Inc., Cary, NC) using Fisher's protected least significant difference. Experimental values are the means ± S.E.
Grading of Samples.
Samples were graded (by J.L.S. and P.V.D.) for inflammation or fibrosis according to the Metavir scoring system (Bedossa, 1994), and the mean mRNA levels was determined for each group. To determine the effect of inflammation, samples were graded into three groups according to the inflammation component of their Metavir score (Table2). Groups consisted of samples with low (score 0), medium (score 1), and high inflammation scores (score 2 or 3). To determine the effect of fibrosis, samples were separated into four groups consisting of samples with no fibrosis (score 0), mild fibrosis (score 1), medium fibrosis (score 2), and high fibrosis or established cirrhosis (score 3 or 4; Table 2).
Demographics of sample groups
Because insufficient cirrhotic (score 4) or inflammation (score 3) samples were available, samples with a fibrosis score of 3 or 4 or an inflammation score of 2 or 3 were grouped together for statistical purposes. The mean value for samples with a fibrosis or inflammation score of 0 was set at 100%, and means in other groups are expressed relative to this number. Graphs show the mean mRNA levels ± S.E. for each group.
Results
Interindividual Variation.
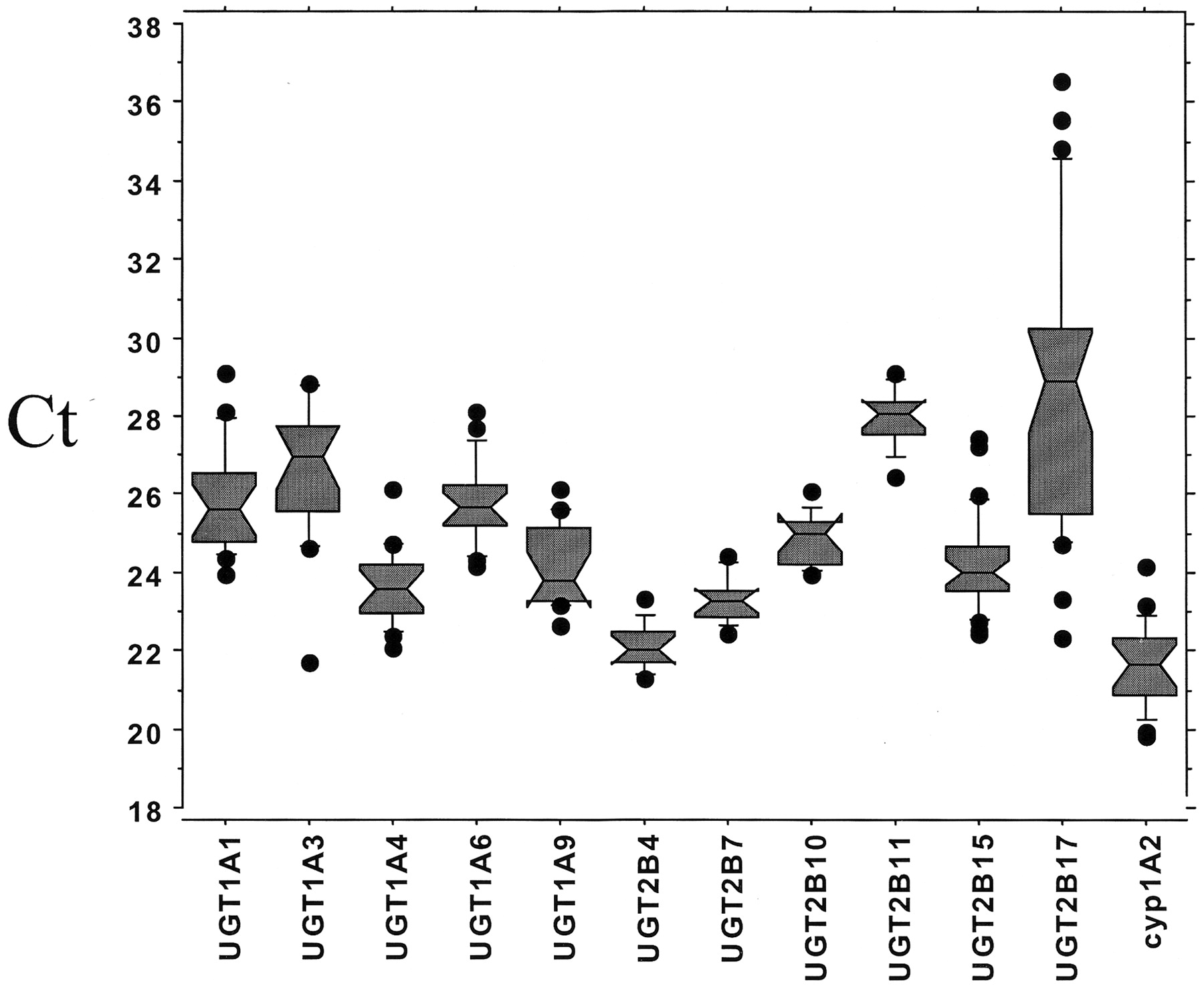
Most UGT isoforms, with the exception of UGT2B17, were readily detectable. Ct values were between 20 and 30 (Fig. 1).

Distribution of Ct values.
Distribution Ct values at which measurement of the PCR product was made.
Interindividual variability was estimated by analyzing results from samples that had minor or no inflammatory activity or fibrosis (inflammation and fibrosis scores of 1 or 0). We found a large interindividual variation in enzyme mRNA levels, ranging from 5- to 15-fold for most isoforms. Of the UGT enzymes, isoform 2B17 had the greatest interindividual variability (Fig. 1; Table3).
Descriptive statistics for UGT family and CYP1A2 genes showing interindividual variability
Effect of Fibrosis.
We found no difference in UGT mRNA levels between the low-score (fibrosis 0) and high-score (fibrosis 3–4) groups using primers designed to amplify all (Fig. 2A) or individual isoforms (Fig. 2B).

Effect of fibrosis on mRNA levels.
Samples were separated into four groups according to their fibrosis score. Values are arbitrary units determined by reference to a standard curve generated for each isoform and normalized to the level of 18S ribosomal RNA determined for each sample. The fibrosis samples with a score of 0 are set to 100%. Data show means ± S.E. ∗, indicates p < 0.05. A, levels of UGT, CYP1A2, TGF-β, and IL-1β mRNA. UGT family 1 and family 2 genes were measured using primers designed to amplify all known isoforms. B, levels of individual UGT isoforms.
Levels of mRNA for CYP1A2, an enzyme representative of the cytochrome P450 family, tended to be elevated in fibrosis groups with a score of 1 and 2, and these were significantly higher than levels in the fibrosis groups with a high score (3 to 4). However, levels of the mRNA for TGF-β, a cytokine known to promote matrix deposition (Border and Noble, 1994), tended to be higher in the high-score fibrosis group (Fig. 2A). Levels of the mRNA for the cytokine IL-1β, frequently associated with reduced cytochrome P450 expression (Abdel-Razzak et al., 1995; Calleja et al., 1998; Kudo and Kawano, 1999) showed no apparent trend.
Effect of Inflammation.
UGT family 1A and family 2B mRNA levels as measured by primers designed to amplify all isoforms within each family tended to be lower in high-inflammation score samples (Fig.3A). Measurement of individual isoforms using specific primers showed a consistent downward trend in the levels of most UGT isoforms in high-inflammation score samples, reaching statistical significance for isoforms 1A4, 2B4, and 2B7 (Fig. 3B), which were reduced to 43.0 ± 12.3, 39.6 ± 19.5, and 38.4 ± 20.3% of control levels, respectively.
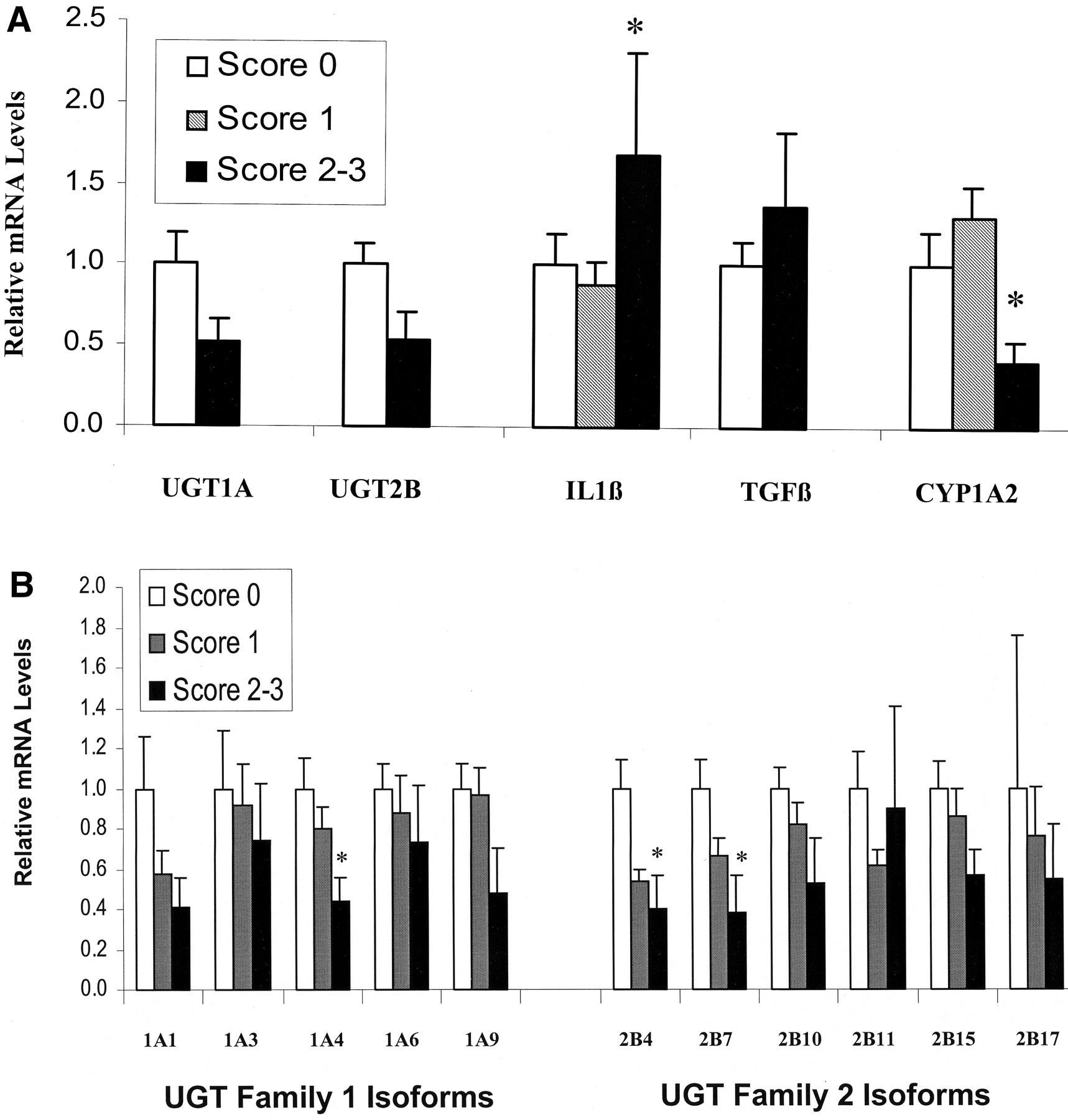
Effect of inflammation on mRNA levels.
Samples were separated into three groups according to their inflammation score. Values are arbitrary units determined by reference to a standard curve generated for each isoform and normalized to the level of 18S ribosomal RNA determined for each sample. The inflammation samples with a score of 0 are set to 100%. Data show means ± S.E. ∗, indicates p < 0.05. A, levels of UGT, CYP1A2, TGF-β, and IL-1β mRNA. UGT family 1 and family 2 genes were measured using primers designed to amplify all known isoforms. B, levels of individual UGT isoforms.
High-score inflammation samples had lower CYP1A2 and higher IL-1β mRNA levels than samples with a score of 1 (Fig. 3A). TGF-β mRNA levels in the group with score from 2 to 3 were similar to those with a score of 0.
Discussion
Although it is commonly accepted that liver disease results in a decreased activity of the cytochrome P450 system, the effect on glucuronidation is controversial (Hoyumpa and Schenker, 1991). Glucuronidation has commonly been determined from either pharmacokinetic studies in humans or experimental models of liver disease in animals. UGT isoforms have overlapping specificities, and a metabolite may be the product of several isoforms. Furthermore, extra hepatic metabolism may be significant (McGurk et al., 1998; Nowell et al., 1999), hepatic blood flow may be impaired, or the binding of drug to plasma proteins may be affected (Davies and Skjodt, 2000). The direct measurement of UGT mRNA in hepatocytes overcomes limitations and assumptions of the pharmacokinetic approach. A number of studies in animals and hepatic cell lines (Vargas et al., 1998; Kardon et al., 2000; Walle et al., 2000; Luquita et al., 2001) and in humans (Ritter et al., 1999) indicate that hepatic UGT protein and/or activity, for at least some isoforms, in general follow the mRNA levels.
In these studies, we used real-time PCR to examine the effect of inflammation and fibrosis on the levels of individual isoforms of the UGT family in patients with liver disease. We found that a reduction in the expression of UGT mRNA in our samples was associated with the degree of inflammation. There was no correlation with the level of fibrosis. In addition our data indicates that the large interindividual variability in drug metabolism, which is often observed across patients, may at least partly be a result of differential mRNA levels of these enzymes. This may be a result of genetic factors or even induction by dietary components.
In a number of studies in human liver (Debinsky et al., 1995) or animal models of cirrhosis (Debinsky et al., 1996), results have indicated that levels of the glucuronidating enzymes as measured by immuno or hybridization histochemistry are maintained or elevated when compared with control tissue. Our results support findings that glucuronidation is unaffected in the fibrotic liver and that hepatic UGT mRNA levels are not reduced.
The mechanism by which the inflammatory process may repress these enzymes is complex and poorly understood (Morgan et al., 1998). The reduction in UGT mRNA in our high-score inflammation samples may be the result of substances released by resident or infiltrating inflammatory cells, as evidenced by the increase in interleukin-1β mRNA in the high-score inflammation group. A number of studies have found that cytokines can affect the expression of UGT (Monshouwer et al., 1996;Levesque et al., 1998; Strasser et al., 1998) or cytochrome P450 enzymes (Ferrari et al., 1993; Abdel-Razzak et al., 1995; Monshouwer et al., 1996; Sindhu et al., 1996; Guillen et al., 1998; Levesque et al., 1998; Kudo and Kawano, 1999; Monshouwer and Witkamp, 2000). There is evidence for the involvement of nuclear factor-κB in the repression of the rat CYP2C11 gene in response to bacterial lipopolysaccharide or interleukin-1β, but interleukin-6 induced repression occurs by an alternative mechanism (Iber et al., 2000). In addition, because our data are standardized to 18S ribosomal RNA, some reduction in hepatic UGT mRNA is expected as a result of an increase in nonhepatic 18S RNA by infiltrating inflammatory cells.
Because of the consistent downward trend observed across most isoforms, the relatively small number of samples with severe liver disease, and the large variation, more comprehensive studies will need to be done to establish the precise relationship between inflammatory liver disease and the expression of individual UGT isoforms. However, the reduction in mRNA levels reached statistical significance for three of the isoforms that we measured, indicating that inflammatory liver disease may have differential effects on the various UGT isoforms.
Our data indicate that there is the potential for decreased drug glucuronidation in the liver of patients with an active inflammatory condition and some hepatic fibrosis, but glucuronidation may recover once the inflammation subsides. This may help explain the uncertainty surrounding drug glucuronidation in liver disease using pharmacokinetic approaches in which experimental results may be from pools of patients with mixed inflammatory/cirrhotic conditions.
Footnotes
- Abbreviations used are::
- UGT
- uridine 5′-diphosphate glucuronosyltransferase
- PCR
- polymerase chain reaction
- IL-1β
- interleukin-1β
- TGF-β
- transforming growth factor-β
- Ct
- the threshold cycle
- Received August 9, 2001.
- Accepted October 29, 2001.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}