


Abstract
Investigation of human UDP-glucuronosyltransferase (UGT) isoforms has been limited by a lack of specific substrate probes. In this study serotonin was evaluated for use as a probe substrate for human UGT1A6 using recombinant human UGTs and tissue microsomes. Of the 10 commercially available recombinant UGT isoforms, only UGT1A6 catalyzed serotonin glucuronidation. Serotonin-UGT activity at 40 mM serotonin concentration varied more than 40-fold among human livers (n = 54), ranging from 0.77 to 32.9 nmol/min/mg of protein with a median activity of 7.1 nmol/min/mg of protein. Serotonin-UGT activity kinetics of representative human liver microsomes (n = 7) and pooled human kidney, intestinal and lung microsomes and recombinant human UGT1A6 typically followed one enzyme Michaelis-Menten kinetics. Serotonin glucuronidation activity in these human liver microsomes had widely varying Vmax values ranging from 0.62 to 51.3 nmol/min/mg of protein but very similar apparentKm values ranging from 5.2 to 8.8 mM. Pooled human kidney, intestine, and lung microsomes hadVmax values (mean ± standard error of the estimates) of 8.8 ± 0.4, 0.22 ± 0.00, and 0.03 ± 0.00 nmol/min/mg of protein (respectively) and apparentKm values of 6.5 ± 0.9, 12.4 ± 2.0, and 4.9 ± 3.3 mM (respectively). In comparison, recombinant UGT1A6 had a Vmax of 4.5 ± 0.1 nmol/min/mg of protein and an apparent Km of 5.0 ± 0.4 mM. A highly significant correlation was found between immunoreactive UGT1A6 protein content and serotonin-UGT activity measured at 4 mM serotonin concentration in human liver microsomes (Rs = 0.769; P < 0.001) (n = 52). In conclusion, these results indicate that serotonin is a highly selective in vitro probe substrate for human UGT1A6.
Glucuronidation, catalyzed by the UDP-glucuronosyltransferases (UGTs1), represents an important metabolic process in mammals involving the conjugation of drugs, xenobiotics, and endogenous compounds to glucuronic acid (Mackenzie et al., 1997). UGT1A6 is an important UGT isoform known to catalyze the glucuronidation of planar and short aromatic molecules, including drugs such as acetaminophen and potential carcinogens chemically related to hydroxylated polycyclic aryl hydrocarbons (Bock et al., 1993). In humans, UGT1A6 is expressed in liver, kidney (Ouzzine et al., 1994), brain (Martinasevic et al., 1998; Gradinaru et al., 1999), lung (Munzel et al., 1996), and intestine (Strassburg et al., 2000). The expression of UGT1A6 in human liver has been reported to be extremely variable as shown by assays using specific antibodies (Ouzzine et al., 1994).
Isoform selective substrate probes have proved extremely useful in quantifying interindividual variability in the function of drug metabolizing enzymes in vitro and in vivo (Venkatakrishnan et al., 2001). However, like other xenobiotic-metabolizing enzymes, the UGTs have overlapping substrate specificities and most substrates are glucuronidated by more than one isoenzyme (Radominska-Pandya et al., 1999). The impact of genetic differences in individual UGT isoforms on drug glucuronidation remains to be established because of lack of specific probe substrates. UGT1A6 activity has been quantified using a number of probe substrates, including α-naphthol (Harding et al., 1988; Ebner and Burchell, 1993), 4-methylumbelliferone (Sabolovic et al., 2000), 4-nitrophenol (Hanioka et al., 2001), and acetaminophen (Fisher et al., 2000; Court et al., 2001b). Unfortunately, studies using expressed UGTs and immunoinhibition with a UGT1A6 antibody indicate that these substrates are glucuronidated by other isoforms besides UGT1A6 (Tukey and Strassburg, 2000; Tephly and Burchell, 1990;Ouzzine et al., 1994). For example, we have shown that acetaminophen is glucuronidated in vitro by at least two UGT isoforms (UGT1A9 and 1A1) in addition to UGT1A6, with the relative contribution of each isoform dependent on acetaminophen concentration and relative isoform abundance (Court et al., 2001b). As yet, a specific substrate probe for UGT1A6 has not been identified.
Recently, serotonin has been identified as a substrate for human UGT1A6 (King et al., 1999). Although relatively little is known about this biotransformation, serotonin was shown to be glucuronidated by rat liver microsomes (Yokota et al., 1989), tissue homogenates (Leakey, 1978), and perfused rat liver (Tyce et al., 1968). Serotonin glucuronide has also been identified in sheep urine (Bartlet and Gilbert, 1971). Interestingly, serotonin glucuronidation activity was reported to be absent in liver homogenates from domestic cat (Leakey, 1978), which is one of the species that lacks a functional UGT1A6 gene (Court and Greenblatt, 2000). In a previous study in this laboratory, serotonin glucuronidation by human liver microsomes and recombinant UGT1A6 showed almost identical apparentKm values (manuscript submitted). Consequently, serotonin would appear to be a viable candidate as a specific UGT1A6 probe substrate. In the present study, serotonin was evaluated for use as an isoform-specific in vitro probe substrate for human UGT1A6 using recombinant human UGT isoforms and microsomes from various tissues.
Materials and Methods
Materials.
Unless otherwise indicated, most reagents including alamethicin, UDP-glucuronic acid (UDPGA; sodium salt), 5-hydroxytryptamine hydrochloride (serotonin-HCl) were purchased from Sigma-Aldrich (St. Louis, MO). All reagents were of analytical or better grade.
Human Liver, Kidney, Intestinal, and Lung Microsomes.
Livers (n = 55) were randomly selected from a frozen bank maintained in the Department of Pharmacology and Experimental Therapeutics, Tufts University School of Medicine, Boston. Microsomes were prepared from frozen liver by differential centrifugation as previously described (Court and Greenblatt, 1997b). The resultant pellet was reconstituted in 20% glycerol/phosphate buffer, aliquoted, and stored at −80°C. Frozen microsomes were thawed once only immediately prior to use. Microsomal protein concentrations were determined using the bicinchoninic acid assay (Pierce Chemical, Rockford, IL). The quality of the liver samples used was ascertained by reference to at least 10 other UGT and cytochrome P450 enzyme activities measured in this laboratory using the same set of liver samples. Livers that consistently showed low activity values (> 2-fold lower for all measured activities) relative to the median activity value for the entire liver set were excluded from the study. Human kidney (pooled, 6 donors) and intestinal (pooled, 10 donors) microsomes were purchased from In Vitro Technologies Inc. (Baltimore, MD). Positive control activity data showed testosterone 6-β-hydroxylation to be 38 and 34 pmol/min/mg of protein for kidney and intestinal microsomes, respectively. Human lung microsomes (pooled, 6 donors) were purchased from Human Biologics International (Scottsdale, AZ). Positive control activity data for this preparation showed 7-ethoxyresorufinO-dealkylation to be 2.1 pmol/min/mg of protein, and cytochrome c reductase as 47 nmol/min/mg of protein.
Recombinant UGT Isoforms.
Microsomes from insect cells transfected with baculovirus containing cDNA for human UGT isoforms 1A1, 1A3, 1A4, 1A6, 1A8, 1A9, 2B7, and 2B15 were obtained from BD Gentest (Woburn, MA) whereas microsomes containing UGTs 1A7 and 1A10 were obtained from Panvera Corp. (Madison, WI). Lymphoblast derived microsomes containing cDNA-expressed human UGT1A6 was obtained from BD Gentest.
Serotonin Glucuronidation Assay.
Microsomal serotonin glucuronidation activities were measured using a method described by S. Krishnaswamy, S. X. Duan, L. L. von Moltke, D. J. Greenblatt, J. L. Sudmeier, W. W. Bachovchin, and M. H. Court (submitted) with slight modifications. Briefly, incubation mixtures contained 25 mM phosphate buffer (pH 7.5), 2.5 mM MgCl2, and UDPGA (20 mM for kinetic studies, otherwise 5 mM) in a final volume of 100 μl. Alamethicin (0.0025% w/v) was also included in HLM incubations at a concentration determined in preliminary experiments to result in optimal activation. Protein concentrations of 0.05 to 0.8 mg/ml of microsomes were used. Incubations were performed in a water bath for 20 to 60 min at 37°C and terminated by addition of 100 μl of acetonitrile containing acetaminophen as an internal standard. After centrifugation at 16,000g for 10 min, the supernatants were dried down in a vacuum oven at 45°C and reconstituted with 150 μl of water and used for HPLC. Standard curves were generated using purified serotonin glucuronide (10 to 100 nmol). Serotonin glucuronide was obtained after incubation of serotonin with mouse liver microsomes under conditions mentioned above followed by HPLC purification and concentration as described by S. Krishnaswamy, S. X. Duan, L. L. von Moltke, D. J. Greenblatt, J. L. Sudmeier, W. W. Bachovchin, and M. H. Court (submitted). The lower limit of sensitivity of the assay under these conditions was 3 pmol/min/mg of protein. Serotonin glucuronidation activities of recombinant UGT isoforms and vector controls were measured at 4 and 40 mM serotonin concentrations using 5 mM UDPGA and 0.5 mg/ml microsomal protein concentration. Incubation was for 30 min at 37°C. Serotonin glucuronidation kinetics with recombinant UGT1A6 and human liver microsomes were performed over a range of serotonin concentrations (0.2 to 40 mM) with a fixed UDPGA concentration of 20 mM.
Other UGT Activities.
UGT activities using nine other marker substrates (estradiol, trifluoperazine, 4-nitrophenol, propofol, acetaminophen, azidothymidine, R- and S-oxazepam and morphine) were measured with the same set of human liver microsomes. Acetaminophen-UGT activity was assayed as described previously (von Moltke et al., 1993; Court and Greenblatt, 1997a). All other assays were similar to that described for serotonin-UGT with the following relevant differences. Final aglycone concentrations were 100 μM for propofol and estradiol, 200 μM for trifluoperazine, and 1 mM for 4-nitrophenol and oxazepam. Microsomal protein concentrations were 0.8 mg/ml for oxazepam-UGT activity, 0.5 mg/ml for AZT-UGT and morphine-UGT; 0.25 mg/ml for trifluoperazine-UGT, propofol-UGT, and estradiol-UGT activities; and 0.05 mg/ml for 4-nitrophenol-UGT activity. Incubation times were 20 min for 4-nitrophenol-UGT; 30 min for propofol-UGT, trifluoperazine-UGT, morphine-UGT, and estradiol-3-UGT; and 120 min for AZT-UGT and oxazepam-UGT activities. Internal standards were acetaminophen for trifluoperazine-UGT, morphine-UGT, and AZT-UGT; thymol for propofol-UGT; and phenacetin for estradiol-3-UGT, 4-nitrophenol-UGT, and oxazepam-UGT activities. The UV absorbance detector was set at 254 nm for trifluoperazine-UGT, 214 nm for propofol-UGT and oxazepam-UGT; 280 nm for estradiol-3-UGT; 266 nm for AZT-UGT; and 295 nm for 4-nitrophenol-UGT activities. A fluorescence detector set at 215 nm excitation wavelength and 350 nm emission wavelength was used for morphine glucuronide. Solvent gradients consisted of 20 to 100% solution B over 20 min for propofol-UGT, 20 to 30% solution B over 15 min for estradiol-3-UGT, 10 to 50% solution B over 15 min for 4-nitrophenol-UGT, and 20 to 40% solution B over 15 min for oxazepam-UGT, and 3 to 8% solution B over 20 min for morphine-UGT assay. For the trifluoperazine-UGT assay, solution A consisted of 0.1% trifluoroacetic acid in water and a linear gradient of 10 to 70% solution B over 20 min was used for elution. The C18 column was a 4.6 mm x 25 cm, 10-μm Synergi Hydro-RP (Phenomenex, Torrance, CA). Since pure standards of propofol glucuronide or trifluoperazine glucuronide were not available, concentrations of these metabolites were quantified using a standard curve generated using propofol and trifluoperazine (respectively), assuming similar UV absorbances (C. J. Patten, personal communication, BD Gentest).
UGT1A6 Protein Content.
The relative content of UGT1A6 protein in liver, kidney, lung, and intestine microsomes was determined by adapting a previously described immunoblotting method (Court et al., 2001a). Briefly, 20 μg of microsomal protein (75 μg for lung and 200 μg for intestine) was separated by sodium dodecylsulfate-polyacrylamide gel electrophoresis using a 4 to 15% gradient gel (Criterion; Bio-Rad, Hercules, CA). Proteins were then electrophoretically transferred to polyvinyl difluoride membrane (Immobilon-P; Millipore Corporatiion, Bedford, MA). Blots were blocked in 10% powdered nonfat milk in Tris-buffered saline-Tween (0.15 M NaCl, 0.04M Tris, pH 7.7, and 0.1% Tween 20) and then incubated in Tris-buffered saline-Tween/1% milk containing a 1:1000 dilution of a polyclonal antipeptide UGT1A6 antibody (WB-UGT1A6; BD Gentest). After washing, the blots were incubated in a 1:10,000 dilution of horseradish peroxidase conjugated secondary antibody (Sigma-Aldrich), washed, and chemiluminescence reagent applied (Super Signal; Pierce Chemical). Blots were imaged (Kodak Image Station 440CF; Kodak, Rochester, NY) and bands quantified using Kodak 1D Image Analysis Software (Kodak). Net intensity values were expressed relative to the sample containing the lowest amount of UGT1A6.
Enzyme Kinetic Analysis.
For enzyme kinetic studies, substrate concentration (S) and velocity (V) data were fitted to the appropriate model by nonlinear least-squares regression. Since the appearance of Eadie-Hofstee (V/S versus V) plots for all sets of data of serotonin-UGT kinetic study were consistent with single enzyme kinetics, the standard Michaelis-Menten model was used to derive the kinetic parametersVmax (maximal velocity) and apparentKm (substrate concentration at half-maximal velocity). Parameters are given as the estimate ± the standard error of the estimate from the nonlinear regression analysis.
Statistical Analyses.
Statistical analyses were performed using the Sigmastat 2.03 software package (SPSS, Chicago, IL). A “P ” value of less than or equal to 0.05 was considered significant. Potential differences in serotonin glucuronidation activities related to gender and species were evaluated by the Mann-Whitney rank sum test. Linear regression analysis was used to determine the effect of age on serotonin glucuronidation activities. Spearman rank order correlation analysis was used to determine the relationship between UGT1A6 protein content and UGT activities. For this, a Spearman correlation rank order correlation coefficient (Rs) of greater than 0.7 and a P value of less than 0.001 were considered significant.
Results
Serotonin Glucuronidation by Expressed UGTs.
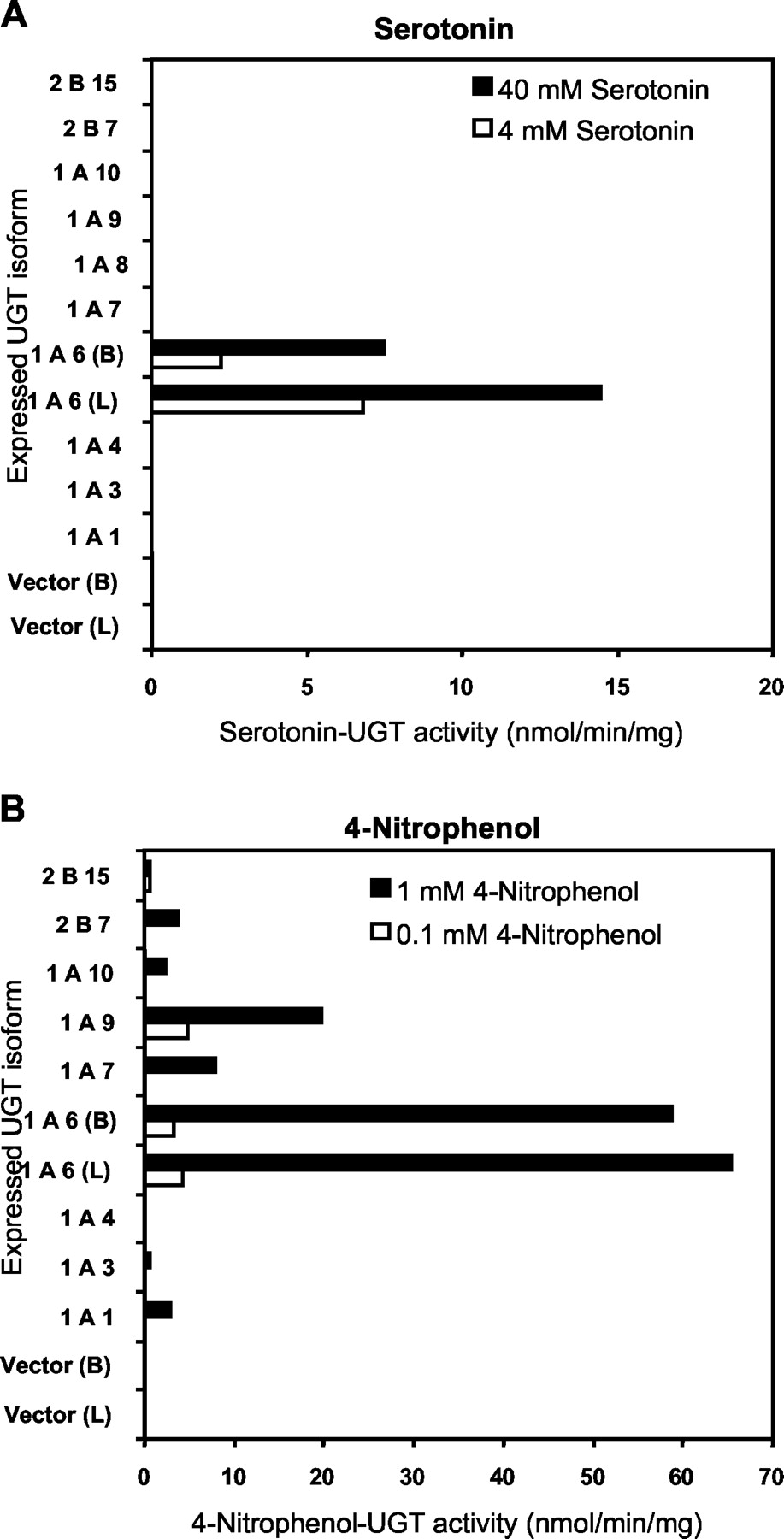
Serotonin glucuronidation activity of UGT isoforms was measured at 4 mM (approximate Km of human liver microsomes) and 40 mM serotonin concentrations using all commercially available recombinant human UGT isoforms including UGT 1A1, 1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2B7, and 2B15 (Fig.1A). At both these substrate concentrations only UGT1A6 isoform showed serotonin glucuronidation activity and none of the other isoforms or the vector controls had any measurable activity (lower limit of quantification 3 pmol/min/mg of protein). Lymphoblast expressed UGT1A6(L) had about 3 times higher serotonin glucuronidation activity (6.8 nmol/min/mg of protein) than insect cell expressed UGT1A6(B) (2.2 nmol/min/mg of protein). Using the same set of recombinant UGT isoforms (except UGT1A8), glucuronidation of a commonly used UGT1A6 probe substrate, 4-nitrophenol, were measured at 0.1 and 1 mM substrate concentrations (Fig. 1B). At 0.1 mM concentration, 4-nitrophenol was glucuronidated with similar activities by UGTs 1A6(L), 1A6(B), and 1A9 (3.3–4.82 nmol/min/mg of protein) and to a lesser extent by UGTs 2B7 and 2B15 (0.23 and 0.54 nmol/min/mg of protein, respectively). At 1 mM concentration, 4-nitrophenol was glucuronidated largely by 1A6(L) and 1A6(B) isoforms (65.3 and 58.8 nmol/min/mg of protein, respectively) and to a lesser extent (3–20 fold less) by UGT isoforms 1A1, 1A7, 1A9, and 2B7 (2.8–19.8 nmol/min/mg of protein).

Serotonin-UGT activities measured at 4 and 40 mM serotonin concentrations (panel A) and 4-nitrophenol-UGT activities measured at 0.1 and 1 mM 4-nitrophenol concentrations (panel B) using microsomes from baculovirus-infected insect cells expressing individual UGT isoforms or vector control.
Activities by microsomes from human lymphoblast cells expressing UGT1A6(L) were also included for comparison. Activities are expressed as a reaction velocity (nanomoles of serotonin glucuronide formed per minute per milligram of protein).
Kinetics of Tissue Microsomes and Recombinant UGT1A6.
Enzyme kinetic studies were performed with a fixed high UDPGA (20 mM) concentration and varied concentrations of serotonin (from 0.2 to 40 mM) using microsomes from representative low (LV3, LV19, and LV22), intermediate (LV39 and LV49), and high (LV35 and LV41) serotonin glucuronidation activity livers, kidney, intestine, and lung, as well as recombinant human UGT1A6(B). Enzyme kinetic data for recombinant UGT1A6(L) from a previous study (S. Krishnaswamy, S. X. Duan, L. L. von Moltke, D. J. Greenblatt, J. L. Sudmeier, W. W. Bachovchin, and M. H. Court, submitted) was included because of its relevance and for comparison. A simple one enzyme Michaelis-Menten kinetic model described all these kinetic data adequately (Fig.2). As shown in Table1, liver microsomes had widely varyingVmax values ranging from 0.62 to 51.3 nmol/min/mg of protein, whereas apparentKm values varied little, between 5.2 and 8.8 mM. Pooled human kidney, lung, and intestinal microsomes hadVmax values of 8.8 ± 0.4, 0.03 ± 0.00, and 0.22 ± 0.00 nmol/min/mg of protein (respectively) and apparent Km values of 6.5 ± 0.9, 4.9 ± 3.3, and 12.4 ± 2.0 mM (respectively). Lymphoblast and insect cell derived recombinant human UGT1A6 preparations showed Vmax values of 15.8 ± 0.2 and 4.5 ± 0.1 nmol/min/mg of protein (respectively), and apparent Km values of 5.9 ± 0.2 and 5.0 ± 0.4 mM (respectively).

Enzyme kinetics of serotonin glucuronidation.
Shown are Michaelis-Menten (panel A) and Eadie-Hofstee (panel B) plots of serotonin-UGT activities measured in low (LV3), intermediate (LV39), and high (LV41) activity human liver microsomes and pooled kidney microsomes with substrate concentrations varied from 0.2 to 40 mM serotonin. Also shown are Michaelis-Menten (panel C) and Eadie-Hofstee (panel D) plots of serotonin-UGT activities measured in insect cell-expressed UGT1A6(B) and lymphoblast cell-expressed UGT1A6(L) with substrate concentrations varied from 0.2 to 40 mM serotonin. Activities are expressed as a reaction velocity (nanomoles of serotonin glucuronide formed per minute per milligram of protein). The solid line connects best-fit estimates of these data determined by nonlinear least-squares curve fit to the Michaelis-Menten equation.
Enzyme kinetics of serotonin glucuronidation by representative liver (n = 7) and pooled kidney, lung and intestine microsomes and UGT1A6 expressed in insect cells (B) or lymphoblast (L)
Interindividual Variability in Serotonin Glucuronidation Activity.
Serotonin glucuronidation activities measured using liver microsomes from 54 individuals at a serotonin concentration of 40 mM ranged from 0.77 to 32.9 nmol/min/mg of protein representing more than a 40-fold variation. Mean and median activity values were 9.3 and 7.1 nmol/min/mg, respectively. The distribution of these data was not normal (Kolmogorov-Smirnov normality test; K-S Dist. = 0.172;P < 0.001) with significant skewing toward lower activities (Fig. 3A). No significant effect of donor gender (P = 0.142) or age (P = 0.856) on serotonin-UGT activities could be discerned. Serotonin glucuronidation activities of the liver microsomes (n = 54) determined at 40 mM concentration of serotonin correlated extremely well with activities determined at 4 mM serotonin (Rs = 0.987; P < 0.001) (Fig. 3B).

A, frequency distribution of serotonin-UGT activities measured in vitro at 40 mM serotonin concentration using HLMs derived from 54 donors; B, comparison of serotonin-UGT activities measured at 4 and 40 mM serotonin concentrations in HLMs (n = 54).
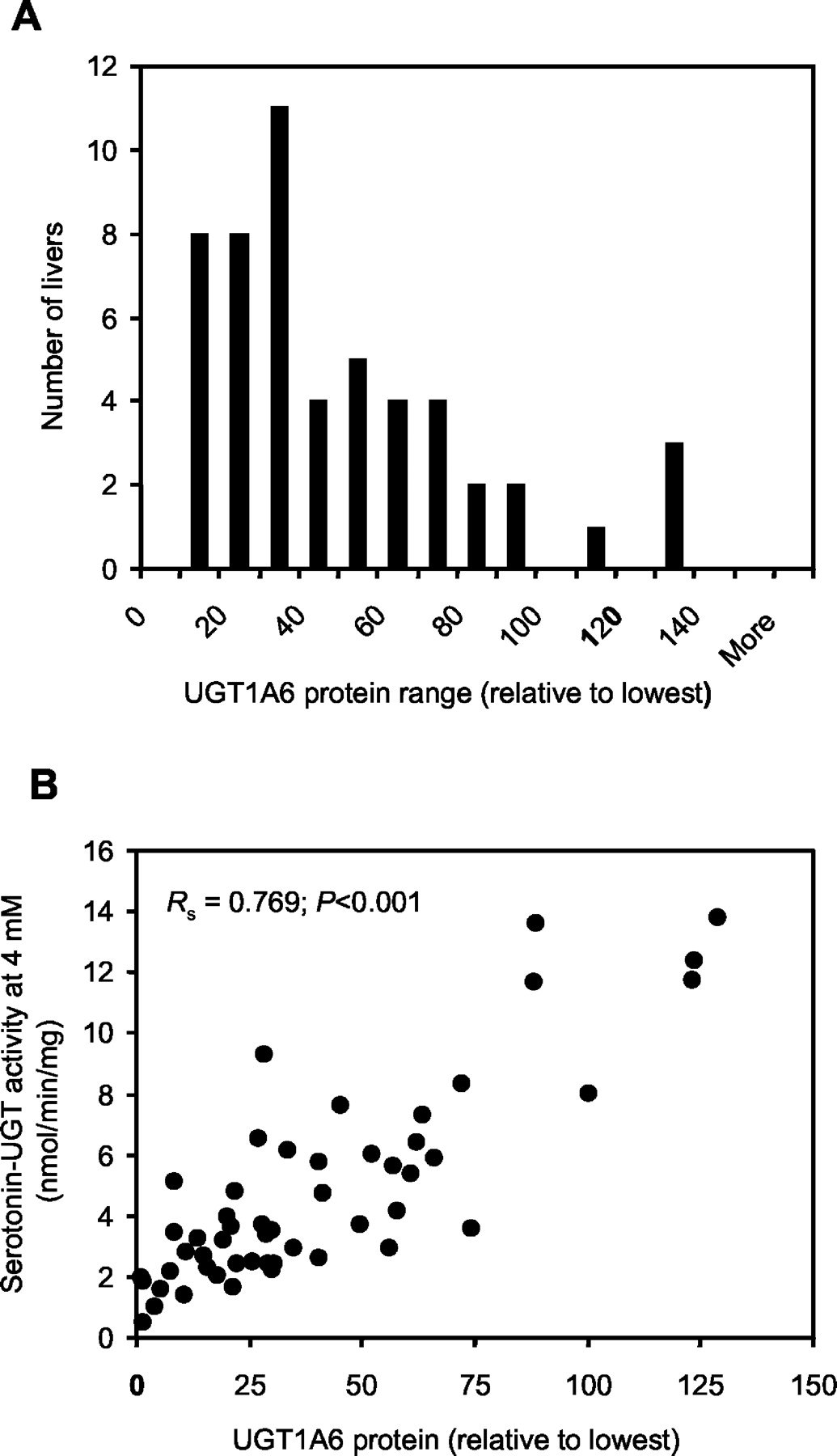
Immunoreactive UGT1A6 Protein in Human Liver and Other Tissue Microsomes.
The relative immunoreactive UGT1A6 protein content of the human liver microsomes (n = 52) determined by Western blotting varied 125-fold. The distribution of these data was not normal (Kolmogorov-Smirnov normality test; K-S Dist. = 0.168;P < 0.001) with significant skewing toward lower protein content (Fig. 4A). No significant effect of donor gender (P = 0.185) or age (P = 0.868) on UGT1A6 protein content was observed.
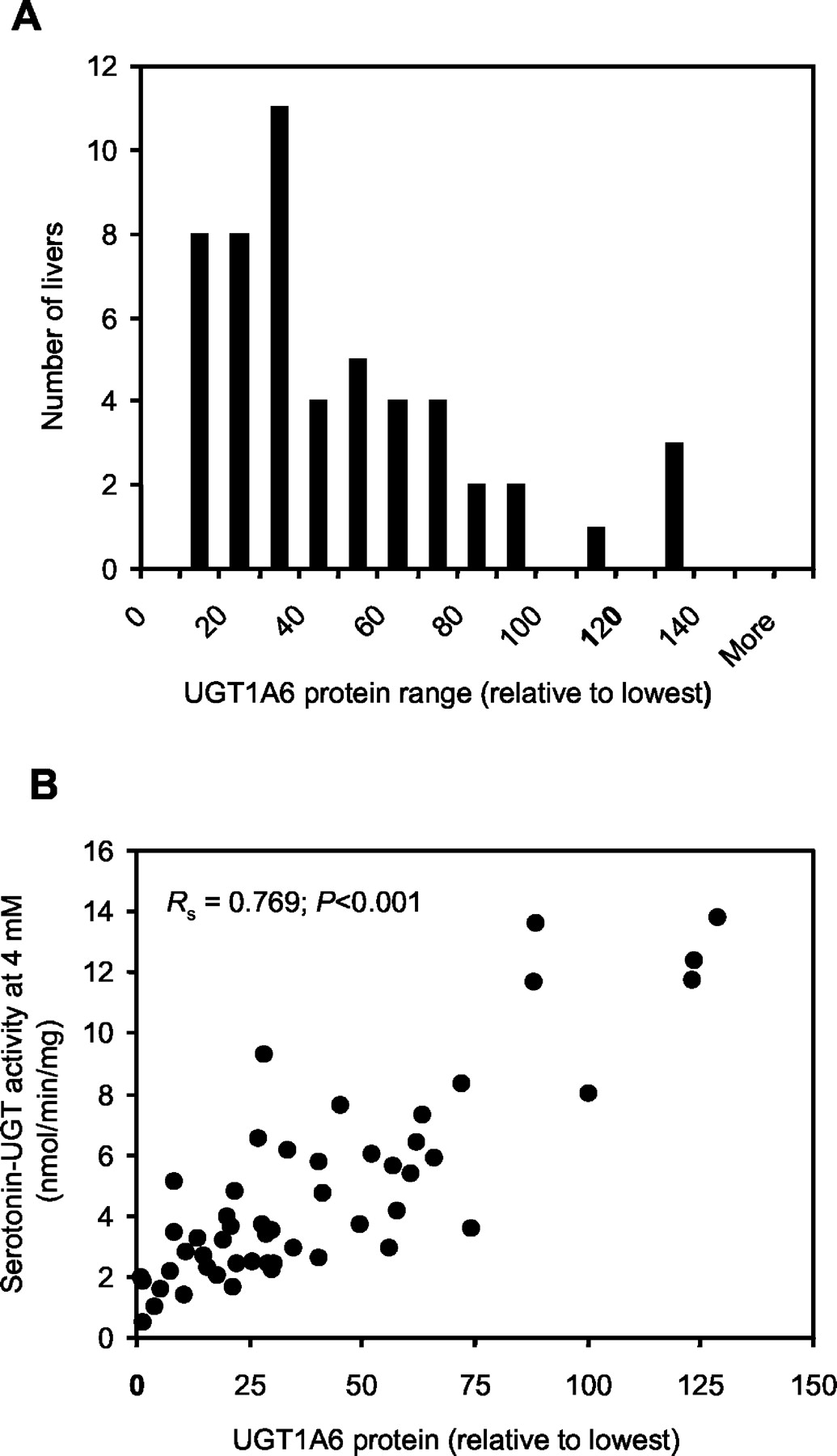
A, frequency distribution plot of UGT1A6 protein; B, comparison of the relative amount of immunoreactive UGT1A6 protein to serotonin-UGT activities measured in the same set of HLMs (n = 52) using 4 mM serotonin concentration.
Also shown is the Spearman rank order correlation coefficient (Rs) and P value for the comparison.
Correlational analyses with immunoquantified UGT1A6 protein as the dependent variable indicated highly significant relationships with serotonin-UGT activity determined at 4 mM (Rs = 0.769; P < 0.001) (Fig. 4B) and at 40 mM serotonin concentration (Rs = 0.721; P < 0.001) and with 4-nitrophenol-UGT activity (Rs = 0.756; P < 0.001) (Table 2). Reasonable correlations were also found with β-estradiol-UGT (Rs = 0.537; P < 0.001), trifluoperazine-UGT (Rs = 0.638; P < 0.001) and propofol-UGT (Rs = 0.433; P < 0.001) activities. No correlational relationships were found with morphine-3-UGT, AZT-UGT, and R-and S-oxazepam-UGT activities.
Correlation of UGT1A6 protein content with UGT activities
The relative UGT1A6 protein content of representative livers (n = 6) and pooled kidney tissue microsomes was compared with Vmax values for serotonin glucuronidation measured for the same set of microsomes (Fig.5A). For liver and pooled kidney microsomes, increasing UGT1A6 protein concentrations correlated extremely well with increasing Vmaxvalues of serotonin glucuronidation kinetics (Rs = 0.964; P < 0.001) (Fig. 5B). UGT1A6 protein band signal for intestinal and lung microsomes (Fig. 5A) was very faint despite loading large amounts (75 and 200 μg, respectively) of protein and could not be quantified.

A, Western blot analysis of UGT1A6 protein in human liver, kidney, intestine, and lung microsomes; B, comparison of the relative amount of immunoreactive UGT1A6 protein with serotonin-UGT Vmax values measured in the same set of liver and pooled kidney microsomes.
Panel A, lanes 1 and 2 were loaded with 0.75 and 0.375 μg (respectively) of total protein of UGT1A6 expressed in insect cells. Lanes 3 to 8 were loaded with microsomes (20 μg of protein) from human livers LV19, LV3, LV22, LV39, LV49, and LV41. Lanes 9 to 11 were loaded with microsomes from pooled kidney (20 μg), intestine (200 μg), and lung (75 μg). Panel B, also shown is the Spearman rank order correlation coefficient (Rs) andP value for the comparison.
Discussion
Lack of specific probe substrates has so far hindered the quantitative estimation of UGT1A6 mediated glucuronidation. To our knowledge, serotonin is the most selective substrate probe for human UGT1A6 identified to date. The isoform selectivity of serotonin glucuronidation was established by showing the absence of glucuronidation activity by any recombinant UGT isoforms other than UGT1A6 and the almost identical apparentKm values for expressed UGT1A6 compared with liver, kidney, and lung microsomes. Furthermore, for the given set of human liver microsomes, immunoreactive UGT1A6 protein level correlated most highly with serotonin glucuronidation activity, whereas marker activities for other UGT isoforms were less significantly or very poorly correlated.
Significant correlation was also found between serotonin and 4-nitrophenol glucuronidation activities. 4-Nitrophenol is a known substrate used widely for quantitative estimation of UGT1A6 (Tephly and Burchell, 1990; Radominska-Pandya et al., 1999). Results from this study indicate that the selectivity of 4-nitrophenol as a UGT1A6 probe may be concentration dependent with significant contributions from other UGT isoforms at lower (<1 mM) substrate concentrations. In contrast, the selectivity of serotonin is likely to be independent of substrate concentration. This is indicated by the lack of serotonin glucuronidation activity by UGTs other than UGT1A6, the highly significant correlation between activities in HLMs measured at 4 and 40 mM serotonin concentration, as well as between the serotonin-UGTVmax values and UGT1A6 protein content in human liver and kidney microsomes. Another advantage of serotonin over other UGT1A6 substrate probes (except perhaps acetaminophen) is that it is nontoxic and could potentially be used as an in vivo probe by ingesting foods such as bananas and walnuts, which contain substantial amounts of serotonin (Feldman and Lee, 1985).
Acetaminophen has been proposed as a specific substrate for UGT1A6 (Bock et al., 1993; Fisher et al., 2000). However studies in our laboratory (Court et al., 2001b) have shown that acetaminophen is glucuronidated by at least two UGT isoforms other than UGT1A6. Although enzyme kinetic studies of the three major isoforms (UGT1A1, 1A6, and 1A9) showed that expressed UGT1A6 had the highest substrate affinity,Km values for liver microsomes were most similar to values determined for expressed UGT1A1 and UGT1A9. Furthermore, acetaminophen glucuronidation activity measured in human liver microsomes did not correlate with UGT1A6 protein content (Rs = 0.30; P = 0.19) but correlated well with propofol glucuronidation activity (Rs = 0.85; P < 0.001) mediated by UGT1A9. Consequently, serotonin may be a better choice than acetaminophen as an isoform-selective substrate for UGT1A6.
UGT1A6 protein content of the liver microsomes showed some correlation with other UGT1A isoform marker activities (estradiol-3-UGT for UGT1A1, trifluoperazine-UGT for UGT1A4, and propofol-UGT for UGT1A9) (Rs = 0.507 to 0.653,P < 0.001). Since there is no evidence for glucuronidation of any of these substrates by UGT1A6, this could result from coregulation of expression of all the UGT1A isoforms as has been previously suggested (Venkatakrishnan et al., 2001). This is likely since all the UGT1A isoforms appear to have evolved by duplication of the exon I and associated upstream regulatory region (Mackenzie et al., 1997).
All commercially available expressed UGTs were screened for serotonin-UGT activity, which includes all of the known UGT1A subfamily isoforms as well as UGT2B7 and UGT2B15. UGT isoforms that were not studied include UGTs 2A1, 2B4, 2B10, and 2B17. Absence of serotonin glucuronidation activity has previously been reported for recombinant human UGT1A1 (Senafi et al., 1994; King et al., 1996), 1A4 (Green and Tephly, 1996), 2B4 (Fournel-Gigleux et al., 1991), 2B7 (Ritter et al., 1990), and 2B15 (Green et al., 1994). UGT2B7 was previously reported to have a small amount of serotonin glucuronidation activity (King et al., 1999), although the assay method used (colorimetric) was less specific than the HPLC method used here. We did incubate UGT2B7 with serotonin for up to 18 h and could not demonstrate serotonin glucuronide formation (data not shown). UGT2B10 has been shown not to have any glucuronidation activity with the previously used substrates of UGT1A6 (α-naphthol, 4-nitrophenol, acetaminophen, and 4-methylumbelliferone) (Jin et al., 1993). Recombinant human UGT2A1 has been shown to glucuronidate both α-naphthol and 4-methylumbelliferone to a significant level (Jedlitschky et al., 1999). However, its expression is highly tissue specific, and UGT2A1 mRNA was absent in liver, and lung in human and rat (Jedlitschky et al., 1999).
Although recombinant-expressed enzymes are extremely useful in evaluating UGT isoform substrate selectivity, certain caveats need to be applied. Firstly, a recent report suggests that UGT substrate selectivity can be altered through formation of heterodimers (Ishii et al., 2001). It is therefore possible that two or more of the isoforms that we identified as being inactive toward serotonin might show significant glucuronidation activity when coexpressed. Secondly, it is also possible that there are novel human UGT isoforms that can use serotonin as a substrate that have not yet been identified, cloned, and expressed.
The very high correlation between the glucuronidation activities for the entire set of liver microsomes measured at 4 and 40 mM serotonin suggests that the Km value may not vary between different livers. This is consistent with the proposed specificity of serotonin as a substrate for UGT1A6 in liver. The slightly higher apparent Km value seen with the intestinal microsomes could be due to the involvement of one or more low affinity isoforms in serotonin glucuronidation in human intestine coupled with low expression of UGT1A6 in this tissue. However, the low Vmax value for intestinal microsomes indicates that absolute contribution to serotonin glucuronidation in vivo by these isoforms probably is minor. The low activity and UGT1A6 protein content in the pooled intestinal microsomes indicate that UGT1A6 expression in intestine is much lower than in liver (averaging less than 5% that of liver). Expression of UGT1A6 has been demonstrated in human duodenum with large interindividual differences in the level of mRNA (Munzel et al., 1996). Compared with liver, significantly lower UGT1A6 activity has been shown in intestinal microsomes on a per milligram of microsomal protein basis (Fisher et al., 2000). The results obtained here are also consistent with observations made earlier in which only mRNA and not the protein for UGT1A6 was detected in human intestinal samples (Strassburg et al., 2000). The results we obtained with pooled kidney microsomes are consistent with earlier reports indicating UGT1A6 expression in kidney, which is comparable with that in the liver (Fisher et al., 2000). Although UGT1A6 expression has been demonstrated in human lung (Munzel et al., 1996; Auyeung et al., 2001) glucuronidation activity has not been shown until now.
Human liver serotonin-UGT activities and UGT1A6 protein content showed a substantial degree of interindividual variability (40- and 125-fold, respectively). This high variability is most likely explained either by extrinsic influences (inducing chemicals in the diet or environment) or by the existence of genetic variations, in the regulatory or expressed regions of the UGT1A6 gene sequence. Identification of serotonin as a highly selective probe substrate for human UGT1A6 will greatly facilitate future investigations of the molecular basis for this variability.
Acknowledgements
This work was supported by Grants RR-00104, GM-61834, DA-05258, MH-58435, DA-13209, DK-58496, DA-13834, AG-17880, and RR-00054 from the National Institutes of Health (Bethesda, MD).
Footnotes
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- UDPGA
- UDP-glucuronic acid
- HLM
- human liver microsomes
- HPLC
- high performance liquid chromatography
- AZT
- 3′-azido-2′,3′-dideoxythymidine
- Rs
- Spearman correlation coefficient
- Received August 13, 2002.
- Accepted October 15, 2002.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}