


Abstract
The effects of microsomal concentration on the inhibitory potencies of four compounds—fluoxetine, quinidine, imipramine, and ezlopitant—on heterologously expressed recombinant CYP2D6-catalyzed bufuralol 1′-hydroxylase activity were determined. Increasing microsomal concentration from 0.0088 to 2.0 mg/ml, using additional microsomes not containing cytochrome P450, resulted in a marked increase in IC50 and KI values for fluoxetine, ezlopitant, and imipramine, when inhibition constants were calculated using the nominal concentration of inhibitor added to the incubation mixture. The extent of nonspecific binding of these inhibitors to microsomes was determined using equilibrium dialysis. The extent of binding increased with increasing microsomal concentration. Binding was greatest for ezlopitant, followed by fluoxetine, imipramine, and quinidine. Correcting inhibition constants for the extent of nonspecific binding resulted in greater consistency of these values with differing microsomal protein concentrations. This effect was also studied with added phospholipid. Inhibition constants increased with increasing phospholipid, and nonspecific binding was also observed for these four drugs to phospholipid. This suggests that the phospholipid component of microsomes possesses some or all of the responsibility for nonspecific binding, and its effect on inhibitors of drug-metabolizing enzymes. These findings suggest that inhibition constants for drugs as inhibitors of microsomal drug-metabolizing enzymes, such as cytochrome P450, should be corrected for the extent of nonspecific binding to components of the in vitro matrix. The implications of this on the prediction of drug-drug interactions from in vitro data are discussed.
The inhibition of drug-metabolizing enzymes, such as cytochrome P450 enzymes, by one drug can lead to alterations in the exposure to a second drug (Thummel et al., 2000). Such drug interactions can have severe consequences in clinical practice, depending upon the potential for toxicity of the affected drug. The need for a profound understanding of drugs to cause such interactions has been highlighted over the past decade, brought about by the demonstrated interactions between azole antifungal agents and other inhibitors of CYP3A enzymes with agents such as terfenadine and cisapride (Honig et al., 1993; Michalets and Williams, 2000;Venkatakrishnan et al., 2000a). Thus, in the discovery and development of new drugs, research efforts are expended in the design of agents intended to be devoid of the liability of causing drug interactions. Although pharmacokinetic drug interactions are easily determined in small phase 1 clinical studies, such studies are not amenable to testing the hundreds to thousands of compounds considered in early drug discovery efforts. To address this need, in vitro inhibition experiments are done to assess the potential for new chemical entities to cause drug interactions.
In order for in vitro experiments to possess value in the prediction of drug interactions in vivo, measured parameters must have a high degree of accuracy and relevance to the in vivo situation. The prediction of the extent of drug interactions in vivo from in vitro data is a difficult process due to the number of variables involved, highlighted by an inability to project in vivo concentrations of the inhibitor that are available to inhibit the drug-metabolizing enzyme (Levy and Trager, 2000). According to classic theory of drug action, the concentration of a drug available to bind a target receptor is that which is free in solution within the target organ. The same theory should hold true for the effect on drug-metabolizing enzymes, considering the drug-metabolizing enzyme to be the “target receptor” in these cases and the liver or gut the target organ in most cases.
In in vitro drug metabolism experiments, it has been previously demonstrated that nonspecific binding of drug molecules to components within the incubation mixture can have an effect on the kinetic constants measured (e.g., KM, CLint; Obach, 1999; Venkatakrishnan et al., 2000b; Kalvass et al., 2001). Nonspecific binding reduces the concentration of free drug available to interact with the enzyme of interest, and thus the nominal concentration of drug added to the incubation may not be truly reflective of the concentration available to interact with the enzyme. Correction for nonspecific binding, by measurement of the unbound fraction of drug under in vitro incubation conditions, can yield more accurate determinations of enzyme kinetic constants. The experiments described herein were undertaken to address the impact of nonspecific binding to microsomes on the determination of enzyme inhibition constants using recombinant heterologously expressed CYP2D6. Examination of the effect of microsomal concentration on IC50 values was made, as this parameter is most frequently measured due to its simplicity. However, the effect of nonspecific binding in vitro could affect IC50values by binding either the inhibitor or the substrate, or both. The effect of nonspecific binding on Kivalues was also determined, which should only be dependent on nonspecific binding of inhibitor and not substrate. Additionally, as microsomes are to a large extent comprised of phospholipid, experiments were conducted in an attempt to determine whether phospholipid or protein possesses the greater responsibility for the effect of nonspecific binding on the capability of these drugs to inhibit CYP2D6-mediated bufuralol 1′-hydroxylase activity.
Materials and Methods
Materials.
Baculovirus-expressed human CYP2D6 microsomes were prepared as previously described (Christopherson et al., 1995). Dioleoylphosphatidylcholine, NADPH, quinidine, metoprolol, and imipramine were obtained from Sigma-Aldrich (St. Louis, MO). Ezlopitant and fluoxetine were synthesized at Pfizer, Inc. (Groton, CT). 1′-Hydroxybufuralol was obtained from Sigma/RBI (Natick, MA).
Microsomal Incubations.
Incubation mixtures consisted of rhCYP2D6 in insect cell microsomes (8.8 μg/ml; 1.04 pmol of P4501/ml), bufuralol (2–40 μM), and NADPH (1.3 mM) in a total volume of 200 μl of KH2PO4 (100 mM, pH 7.5) containing MgCl2 (3.3 mM). Incubations were commenced with the addition of NADPH and carried out at 37°C in a shaking water bath open to air. At 10 min, the reactions were terminated by addition of 100 μl 0.1 M NaOH. In investigations of the effects of microsomal protein on inhibitory activity, insect cell microsomes from nontransfected cells were added to make final protein concentrations ranging from 0.2 to 2.0 mg/ml. For the effect of phospholipid, sonicated stocks of dioleoylphosphatidylcholine (4.0 mg/ml) were added to incubation mixtures to final concentrations of 0.2 to 2.0 mg/ml. All incubations were conducted in duplicate.
Analysis of 1′-Hydroxybufuralol.
To terminated incubation mixtures was added internal standard (50 ng of metoprolol in 20 μl of methanol) and extraction with methylt-butyl ether (3 ml). The organic solvent was evaporated under N2 at 35°C and the residue reconstituted in 0.2 ml of HPLC mobile phase.
Reconstituted extracts were analyzed for 1′-hydroxybufuralol by HPLC-mass spectrometry. The HPLC system consisted of an Agilent model 1100 quaternary HPLC pump with membrane degasser (Agilent, Palo Alto, CA), CTC PAL autosampler (CTC Analytics, Carrboro, NC), and PE Sciex API100 mass spectrometer containing a Turbo Ionspray source (PE Sciex, Thornhill, ON, Canada). The column used was a Phenomenex Luna C18 narrow bore column (2.0 × 50 mm, 5 μ; Phenomenex, Torrance, CA) equilibrated in 20 mM CH3COOH (adjusted to pH 4 with NH4OH) containing 15% CH3CN. The flow rate was 0.5 ml/min. Reconstituted extracts were injected (10 μl) on the column, and analytes were eluted with isocratic flow. The eluent was introduced into the source of the mass spectrometer operated in the positive ion mode at an orifice voltage of 20 V and a source temperature of 400°C. Other state file settings were adjusted to optimize the response. 1′-Hydroxybufuralol and metoprolol were monitored at m/z of 278 and 268 and eluted at 1.1 and 1.4 min, respectively. Quantitation was accomplished from a standard curve ranging from 10 to 2000 nM using 1/x weighting of analyte/internal standard peak height ratios.
Determination of Binding.
Nonspecific binding of inhibitors and substrate to microsomes or phospholipid was determined by equilibrium dialysis. Compounds, at concentrations proximal to the nominal IC50values, were added to solutions containing microsomes (0.0088 to 2.0 mg/ml) or phospholipid (0.2 to 2.0 mg/ml), KH2PO4 (0.1 M, pH 7.4) and MgCl2 (3.3 mM). These solutions were subject to equilibrium dialysis using an in-house devised 96-well apparatus similar to that described by Kariv et al. (2001), using dialysis membranes with a molecular weight cutoff of 12 to 14 kDa. The dialysis buffer was KH2PO4(0.1 M, pH 7.4) and MgCl2 (3.3 mM). Dialysis was conducted in triplicate for 5 h to attain equilibration at 37°C. Analysis of dialysates for bufuralol, fluoxetine, quinidine, ezlopitant, and imipramine was accomplished by HPLC-mass spectrometry. All samples were alkalinized with NaOH (0.1 ml, 0.1 M), extracted with methyl t-butyl ether (3 ml), the organic fraction evaporated under N2, the residue reconstituted in HPLC mobile phase (50 μl), and injected (20 μl) onto a Phenomenex Luna C18 column (2.0 × 50 mm; 5μ) at a flow rate of 0.5 ml/min in 20 mM CH3COOH (adjusted to pH 4 with NH4OH) containing varying amounts of CH3CN. Mass spectrometer conditions were optimized for each analyte. The following conditions of mobile phase composition, internal standard used, ions monitored, and retention times unique for each of the five compounds are as follows: ezlopitant (mobile phase contained 45% CH3CN, internal standard = 5 ng of [2H3]ezlopitant, ions monitored = m/z 455 and 458,Rt = 1.2 min), fluoxetine (mobile phase contained 45% CH3CN, internal standard = 5 ng of (3-(4-methanesulfonylphenyl)-3-(4-trifluoromethylphenoxy)propyl)dimethylamine, ions monitored = m/z 310 and 402,Rt = 0.7 and 0.5 min), imipramine (mobile phase contained 32% CH3CN, internal standard = 10 ng of clomipramine, ions monitored =m/z 281 and 315, Rt = 1.0 and 2.0 min), quinidine (mobile phase contained 19% CH3CN, internal standard = 10 ng of cinchonidine, ions monitored = m/z 325 and 295,Rt = 1.2 and 1.0 min), and bufuralol (mobile phase contained a gradient of 18 to 28% CH3CN, internal standard = 50 ng of metoprolol, ions monitored = m/z 263 and 268,Rt = 2.8 and 0.7 min).
Calculations.
Enzyme kinetic parameters and inhibition constants were determined using the Enzyme Kinetics module of SigmaPlot (v8, SPSS Science, Chicago, IL). Data were best fit to competitive or partial-competitive models of enzyme inhibition, as assessed by Akaike's information criteria values. Values are reported as mean values with standard errors of the entire datasets. Nonspecific binding to microsomes and phospholipid vesicles was calculated as previously described (Obach, 1999).
Results
Effect of Microsomal Protein Concentration on Inhibition.
Since the activity of bufuralol 1′-hydroxylation is high in recombinant heterologously expressed CYP2D6 and since the analytical method for 1′-hydroxybufuralol developed for these experiments was highly sensitive, very low concentrations of total active microsomal protein in metabolic incubations could be employed. This enabled the examination of the effect of additional noncatalytic microsomes on the inhibitory potency of the following CYP2D6 inhibitors: quinidine, imipramine, fluoxetine, and ezlopitant. The effect of increasing microsomal protein concentration on percentage of control activity versus inhibitor concentration plots for each of these inhibitors are shown in Fig. 1, with IC50 values listed in Table 1. For fluoxetine and ezlopitant, increasing microsomal protein concentration exhibited a substantial decrease in inhibition of bufuralol 1′-hydroxylase activity. The effect on the inhibitory potency of quinidine and imipramine was not as marked, and in fact, the trend for quinidine was such that a slight increase in inhibitory potency with increasing microsomal protein concentration was observed (Table 1).
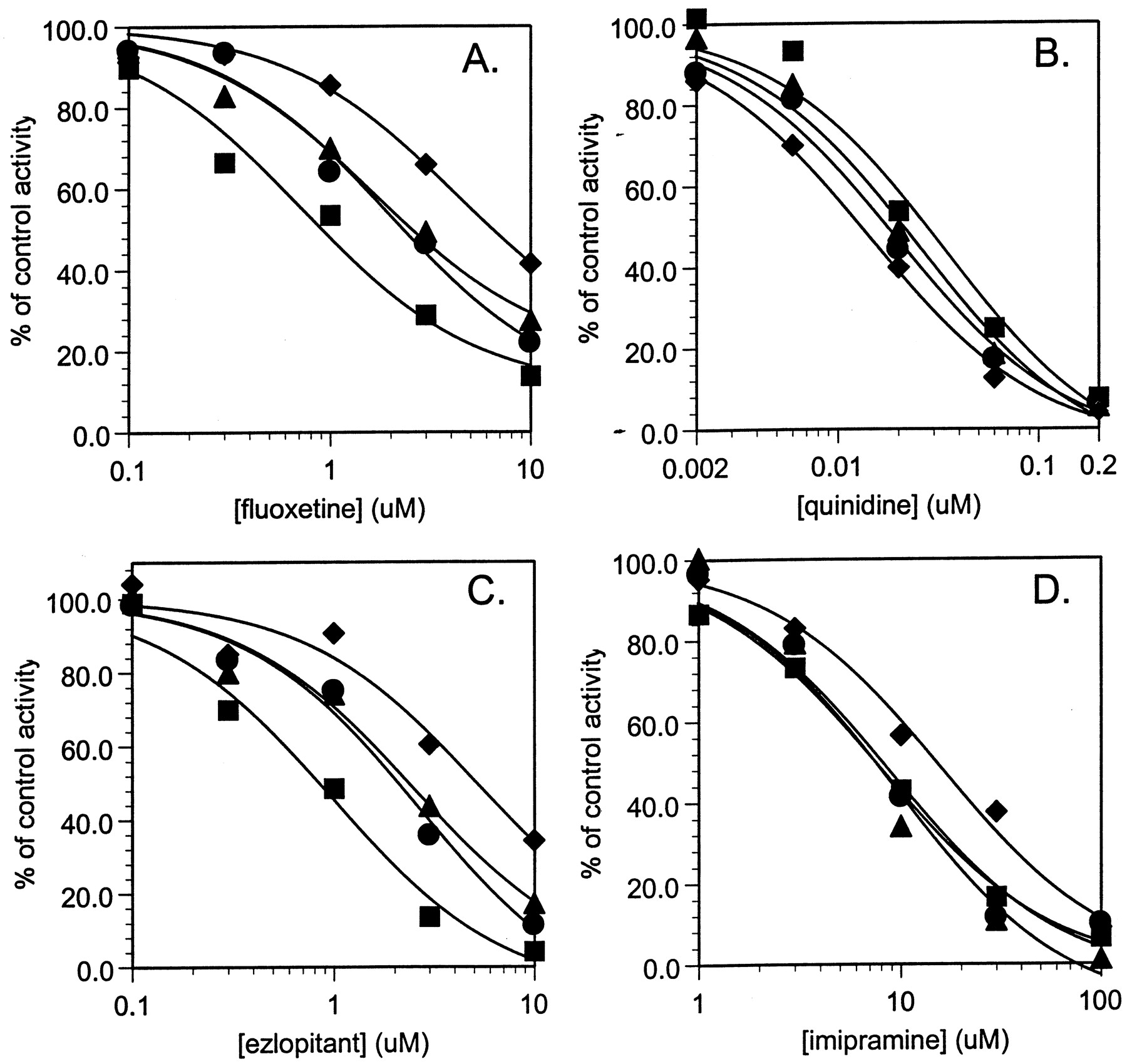
Inhibition of CYP2D6 catalyzed bufuralol 1′-hydroxylase activity by fluoxetine (panel A), quinidine (panel B), ezlopitant (panel C), and imipramine (panel D), in the absence and presence of increasing concentrations of control Sf9 cell microsomes.
Symbols are defined as follows: control, ▪; 0.2 mg/ml, ●; 0.5 mg/ml, ▴; 2.0 mg/ml, ♦.
Inhibition of CYP2D6-catalyzed bufuralol 1′-hydroxylase by fluoxetine, ezlopitant, imipramine, and quinidine with varying microsomal protein and phospholipid concentrations
Effect of Phospholipid Concentration on Inhibition.
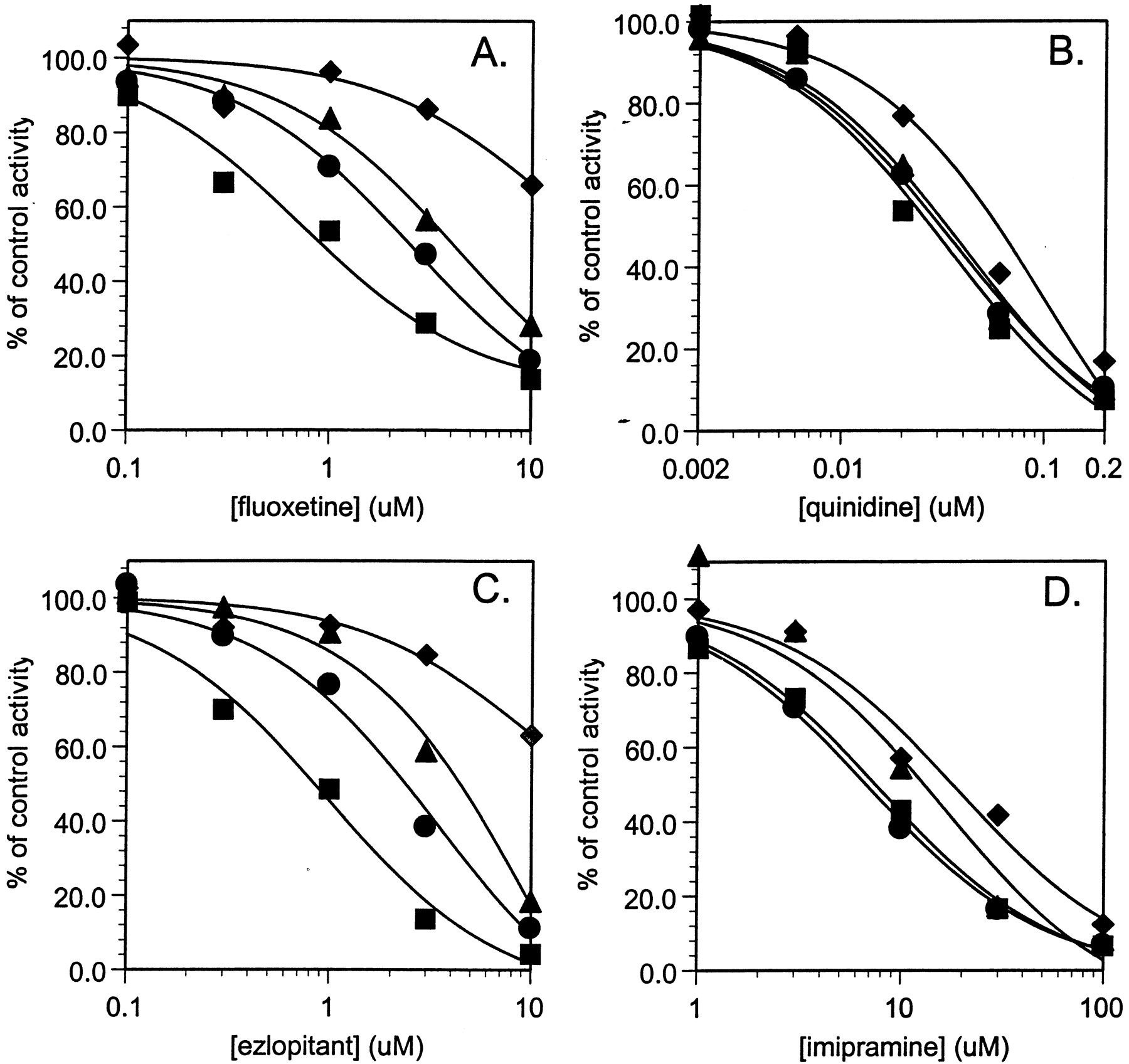
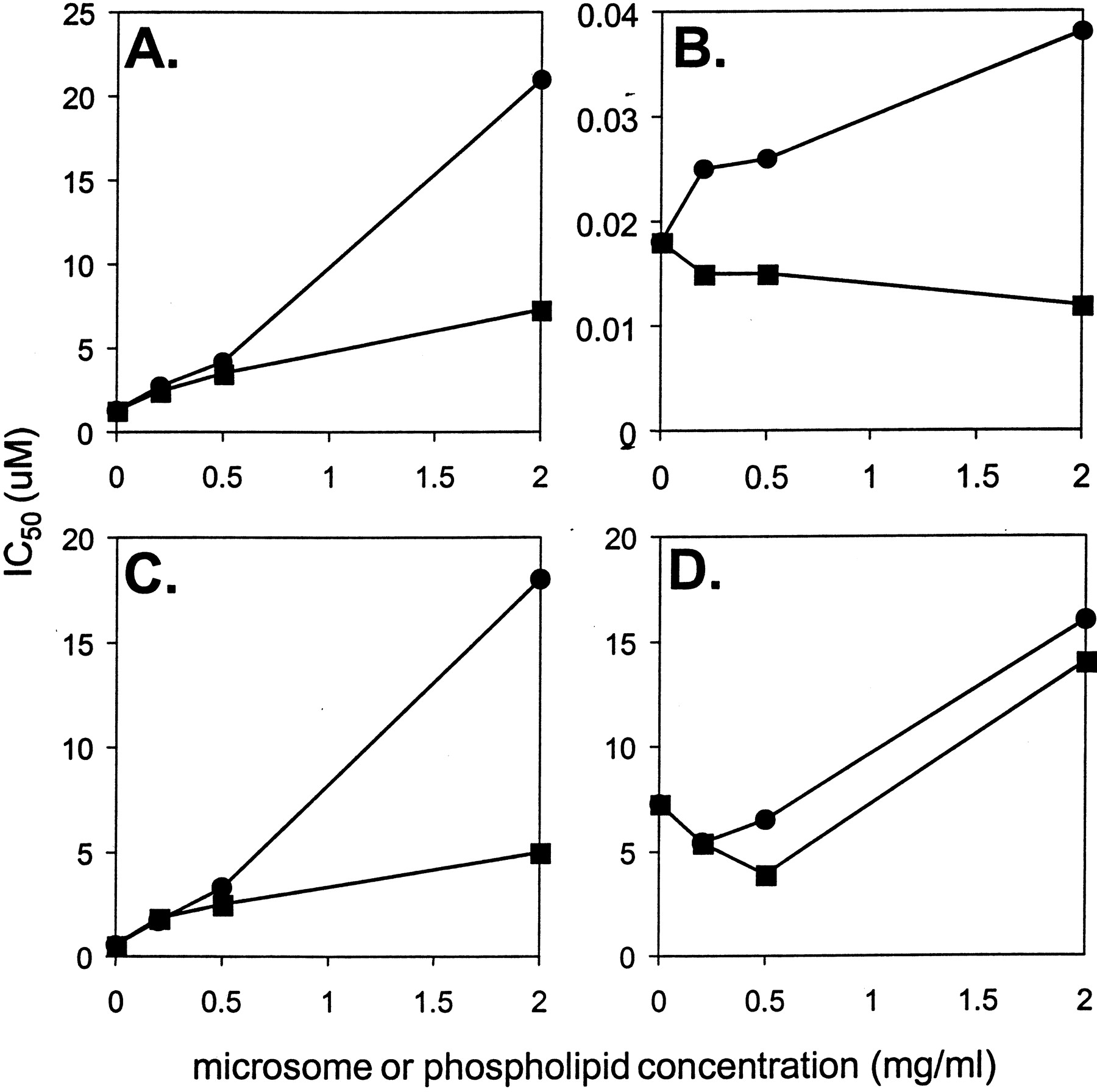
Microsomes are composed of both protein and phospholipid. To address whether the reduction in inhibitory potency with increasing amounts of microsomes is due to protein or phospholipid, the effect of added phospholipid on inhibition was determined (Fig.2). A comparison of the effect of equal concentrations of microsomes or phospholipid on inhibitory activity showed that phospholipid exhibited a greater effect at 2.0 mg/ml and similar effects at the two lower concentrations examined. IC50 values in the presence of 0.2 to 2.0 mg/ml phospholipid are listed in Table 1. Inhibition by fluoxetine and ezlopitant was affected more than inhibition by imipramine and quinidine. The relationship between microsomal or phospholipid concentration and IC50 values are plotted in Fig.3.

Inhibition of CYP2D6 catalyzed bufuralol 1′-hydroxylase activity by fluoxetine (panel A), quinidine (panel B), ezlopitant (panel C), and imipramine (panel D), in the absence and presence of increasing concentrations of phospholipid.
Symbols are defined as follows: control, ▪; 0.2 mg/ml, ●; 0.5 mg/ml, ▴; 2.0 mg/ml, ♦.

Apparent IC50 values as a function of microsomal protein and phospholipid concentrations.
Fluoxetine (panel A), quinidine (panel B), ezlopitant (panel C), and imipramine (panel D). Symbols are defined as follows: microsomes, ▪; phospholipid, ●.
Nonspecific Binding to Microsomes and Phospholipid.
The extent of nonspecific binding of the inhibitors fluoxetine, ezlopitant, quinidine, and imipramine to microsomes and phospholipid is shown in Table 2. Binding increased with increasing microsomal protein and phospholipid concentrations. The binding to phospholipid vesicles appeared to be equal to or greater than binding to microsomes. It should be noted that in microsomes, for every 1.0 mg of microsomal protein there is approximately 0.6 mg of phospholipid (Ernster et al., 1962). Ezlopitant showed the greatest binding, followed by fluoxetine, imipramine, and quinidine. The nonspecific binding of the substrate, bufuralol, was also determined at microsomal protein and phospholipid concentrations of 0.2 to 2.0 mg/ml. Bufuralol also demonstrated an increased amount of binding as phospholipid or microsome concentrations were increased.
Nonspecific binding of bufuralol fluoxetine, ezlopitant, imipramine, and quinidine to Sf9 cell microsomes and phospholipid
Unbound Inhibition Values.
Correction of IC50 values for unbound fractions of the four inhibitors was made, and values are shown in Table3. For ezlopitant and fluoxetine, IC50 values based on free inhibitor concentrations were substantially changed from those based on nominal concentrations due to the high extent of nonspecific binding. Although IC50 values based on nominal inhibitor concentrations increased with increasing microsome or phospholipid concentrations, when inhibitor concentrations were corrected for nonspecific binding, in some cases there was a downward trend in IC50 values with increasing microsome or phospholipid concentrations (Table 3). This may be due to the extent of binding of the substrate, bufuralol. For competitive inhibition, IC50 values are dependent on the substrate concentration used to measure activity, with inhibitors appearing more “potent” at lower substrate concentrations.
Unbound IC50 values for fluoxetine, ezlopitant, imipramine, and quinidine on CYP2D6-catalyzed bufuralol 1′-hydroxylase activity
KI values, which represent binding constants and should thus be independent of substrate concentration, were measured for the four inhibitors at microsomal concentrations of 0.0088, 0.2, and 2.0 mg/ml. KI values determined using nominal substrate and inhibitor concentrations and substrate and inhibitor concentrations corrected for nonspecific binding are listed in Table 4. AlthoughKI values based on nominal inhibitor and substrate concentrations differed with different concentrations of added microsomes, in many cases, KIvalues based on free inhibitor and substrate concentrations at various microsome concentrations converged, although they were not identical.
Ki values for fluoxetine, ezlopitant, imipramine, and quinidine on CYP2D6 catalyzed bufuralol 1′-hydroxylase activity based on total (nominal) and free inhibitor and substrate concentrations
Discussion
When making measurements of activities of compounds using in vitro systems, it is frequently and mistakenly assumed that the components of the system do not affect the concentration of the test compound available to bind to the target receptor or enzyme. However, this has been shown to not be the case, especially for compounds that are lipophilic. In drug metabolism studies, in vitro systems frequently contain phospholipid membranes, either in assays that use whole cells or assays that use subcellular fractions (e.g., microsomes). Compounds that are lipophilic, and especially those that are also cationic at physiological pH, appear to be able to nonspecifically bind to phospholipid membranes (Bickel and Steele, 1974; Di Francesco and Bickel, 1977; McLure et al., 2000). Such nonspecific binding has been demonstrated to have an effect on measurements of microsomal intrinsic clearance (Obach, 1997, 1999) and Michaelis constants for drug metabolism reactions (Venkatakrishnan et al., 2000b; Kalvass et al., 2001).
These experiments were undertaken to assess whether nonspecific binding phenomena has a similar effect on inhibition of drug metabolism reactions, and furthermore, to determine whether microsomal protein or phospholipid is the greater contributor to the effect of nonspecific binding. The system chosen for study was the CYP2D6-mediated hydroxylation of bufuralol using recombinant, heterologously expressed enzyme. Since the specific activity for this reaction is high, very low microsomal protein concentrations were able to be used for control activity and inhibitory potency determinations, and the system could be manipulated to address the experimental questions at hand. Four CYP2D6 inhibitors, all cationic at neutral pH were selected for study that span a range of physicochemical properties. Quinidine is the least lipophilic of the inhibitors studied, followed by imipramine, fluoxetine, and ezlopitant. This was consistent with the amounts of nonspecific binding exhibited by these compounds relative to each other (Table 2).
The data gathered support the hypothesis that nonspecific binding affects measurements of inhibitory potency. In general, as the concentration of microsomes (and hence phospholipid) is increased, the inhibitory potency decreases as exhibited by increases in IC50 values based on nominal inhibitor concentrations. However, IC50 values are also a function of substrate concentration. Bufuralol also exhibited some nonspecific binding, which complicated the relationship between IC50 and microsomal or phospholipid concentration. An IC50 value represents an experimentally convenient parameter to obtain and is typically the parameter of choice when testing a large number of compounds as inhibitors, rather than the more onerous task of measuringKi values. However, it must be kept in mind that IC50 values are functions of both inhibitor and substrate concentrations and that nonspecific binding could occur for either or both of these compounds. On the other hand,KI values are binding constants and are independent of substrate concentration (with the possible exception of when the inhibitor is actually a competing substrate) and therefore should be similar at different microsome or phospholipid concentrations, provided that the nominal substrate and inhibitor concentrations are corrected to free concentrations before enzyme kinetic analysis. Although the data in Table 4 do not provide exact agreement among Ki values corrected for the unbound fraction across the varying microsome concentrations, the values are closer to each other than when left uncorrected.
The data support the hypothesis that the phospholipid component of microsomes is the primary contributor to nonspecific binding of inhibitors. It has been reported that there is approximately 0.6 mg of phospholipid for every milligram of protein in liver microsomes (Ernster et al., 1962). Comparison of the unbound fractions of these compounds with equal concentrations of microsomal protein or phospholipid demonstrates that the extent of binding to phospholipid is equal to or greater than that of microsomes. This is consistent with a previous study in which the binding of chlorpromazine to microsomes, delipidated microsomes, and phospholipid suggested that the latter component was most responsible for binding (Di Francesco and Bickel, 1977). Furthermore, quinidine and imipramine have been demonstrated to bind various phospholipids (Yata et al., 1990). It appears that compounds that have the greatest requirement of correction of inhibition constants for nonspecific binding are those that are the most lipophilic. Also, cationic character at neutral pH, which all four inhibitors and the substrate examined in this study possess, also appears to be an important determinant for such binding (Di Francesco and Bickel, 1977; Obach, 1999), although this structural feature was not specifically tested in this study.
The prediction of the extent of drug interactions via inhibition of metabolism (especially for P450-mediated metabolic clearance) is not straightforward (Levy and Trager, 2000; Lin, 2000; Bachmann and Ghosh, 2001; Venkatakrishnan et al., 2001; Yamano et al., 2001). There appear to be several poorly understood variables that prevent quantitative prediction of drug interactions from in vitro inhibition data. Perhaps most important is a lack of knowledge of the concentration of inhibitor available to the enzyme in vivo. This concentration could be equal only to the unbound concentration of drug within the cell, which in many cases is presumed to be equal to the unbound concentration in the systemic circulation. Alternately, it is not known whether inhibitor that is nonspecifically bound to tissues is available to interact with the drug-metabolizing enzyme. However, the data presented in this report suggest that the concentration of inhibitor available to the enzyme in vitro is not necessarily the nominal concentration added to the in vitro system. Determination of in vitro inhibition constants should be corrected for nonspecific binding of the inhibitor to components of the in vitro matrix. This is consistent with corresponding corrections needed for measurement of intrinsic clearance and Michaelis constants. This binding can have a substantial effect, asKi values for fluoxetine and ezlopitant, determined at 2.0 mg/ml microsomal protein were 40× and 330× greater, respectively, when nonspecific binding was disregarded. Although there are likely other factors that confound our capabilities to predict the magnitude of drug interactions from in vitro data, the correction of in vitro inhibitory potency for free fraction in vitro should provide one more step toward making more accurate drug interaction predictions. In conclusion, these data suggest that the extent of nonspecific binding may frequently need to be determined to obtain meaningful values for inhibitory potency used in either the comparison of drugs or in the prediction of the extent of in vivo drug interaction potential.
Footnotes
- Abbreviations used are::
- P450
- cytochrome P450
- HPLC
- high-performance liquid chromatography
- Received November 15, 2002.
- Accepted February 3, 2003.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}